
Abstract
Salicornia herbacea L. (Chenopodiaceae), an edible salt marsh plant with anti-inflammatory effects, was examined in macrophages and trophoblasts whether it modulates NLRP3 inflammasome activity. Pretreatment and delayed treatment of S. herbacea extract (SHE) in bone marrow-derived macrophages (BMDMs) reduced the activity of NLRP3 inflammasome induced by lipopolysaccharide (LPS) and adenosine triphosphate stimulation and downregulated interleukin (IL)-1β production. SHE also inhibited pyroptotic cell death, the adaptor molecule apoptosis-associated speck-like protein containing a CARD (ASC), oligomerization, and speck by NLRP3 inflammasome activity in BMDM. Similarly, SHE decreased the mRNA expression of NLRP3, ASC, IL-1β, and IL-6 in the LPS-stimulated human trophoblast cell line, Swan 71 cells. In addition, SHE inhibited the production of IL-6 and IL-1β and decreased the expression of cyclooxygenase-2 and prostaglandin E2 in stimulated Swan 71 cells. Finally, 3,5-dicaffeoylquinic acid (3,5-DCQA), one of the components of S. herbacea, inhibited IL-1β produced by NLRP3 inflammasome activity. In conclusion, SHE downregulated the activity of the NLRP3 inflammasome in macrophages and trophoblasts.
Introduction
The NLRP3 inflammasome-mediated inflammation can cause pregnancy complications such as miscarriage, preterm birth, and preeclampsia. 1 –3 The NLRP3 inflammasome consists of NLRP3 (sensor), the adaptor molecule apoptosis-associated speck-like protein containing a CARD (ASC), (adaptor), and caspase-1 (effector). A two-step process (priming and activation) is required for activation of the NLRP3 inflammasome. 4 As a first step (signal 1; priming step), toll-like receptor (TLR) is sensed by pathogen-associated molecular pattern, which activates nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), leading to increased expression levels of NLRP3, prointerleukin (IL)-18, and pro-IL-1β.
The second step (signal 2; activation step) is the activation of NLRP3 by microorganisms, damage-associated molecular patterns, or environmental stimulants, leading to the complex assembly of NLRP3, ASC, and procaspase-1. The complicatedly assembled NLRP3 inflammasome activates caspase-1 to cleave the precursor form of pro-IL-1β and pro-IL-18. Activated caspase-1 also induces pyroptosis by cleaving the N-terminal of gasdermin D. Pyroptosis, an inflammatory form of lytic programmed cell death, promotes the release of the inflammatory cytokines IL-1β and IL-18. 4
Therefore, IL-1β, which is excessively increased by NLRP3 inflammasome activity, is difficult to maintain normal pregnancy. Overproduction of IL-1β by Group B streptococci infection in mice increased preterm birth. 5 IL-1β production was increased in human decidual cells and gestational tissues by bacterial infection or lipopolysaccharide (LPS), a toll-like receptor (TLR)4 ligand, leading to preterm birth. 6,7 So, inhibiting NLRP3 inflammasome activity is essential for a healthy pregnancy.
Salicornia herbacea, Chenopodiaceae, is a plant that grows on salt marshes and muddy seashores along the western coast of Korea. S. herbacea has long been used as a food or folk remedy to treat bowel disease, diabetes, obesity, and hepatitis. 8,9 S. herbacea contains active constituents, including flavonol glycosides, caffeoylquinic acids, sterols, and saponins. Previously, our group reported that the major components of S. herbacea were 12 caffeoylquinic acid derivatives and 3 flavonol glycosides. 10,11
The caffeoylquinic acid derivatives were chlorogenic acid, methyl chlorogenate, 3,5-dicaffeoylquinic acid (3,5-DCQA), 3-caffeoyl-4-dihydrocaffeoylquinic acid, 3,4-dicaffeoylquinic acid, methyl 3,5-dicaffeoyl quinate, methyl 4-caffeoyl-3-dihydrocaffeoyl quinate, 3,4-dicaffeoylquinic acid, 3-caffeoyl-5-dihydrocaffeoylquinic acid methyl ester, 3-caffeoyl-5-dihydrocaffeoylquinic acid, 3,5-di-dihydrocaffeoylquinic acid methyl ester, and 3-caffeoyl-4-dihydrocaffeoylquinic acid methyl ester. The flavonol glycosides were isoquercitrin 6′′-O-methyloxalate, isorhamnetin 3-O-β-
However, the role of S. herbacea extract (SHE) in pregnancy complications caused by NLRP3 inflammasome activation is unknown. Therefore, we investigated whether SHE regulates the activation of the NLRP3 inflammasome in macrophages and trophoblasts and shows a beneficial effect on pregnancy complications.
Materials and Methods
Salicornia herbacea extraction
The aerial parts of S. herbacea were collected in Shinan County, located on the southwestern coast of Korea. The aerial parts were washed, freeze dried, and ground. The SHE was prepared by boiling the dried aerial portion (200 g) with 4 L of distilled water at 90°C for 20 min, followed by vacuum filtration through a glass fiber filter. The residue was re-extracted with 2 L of distilled water at 90°C for 20 min, concentrated by vacuum evaporation, and then freeze dried in a freeze dryer (TFD8503; Ilshin Lab, Seoul, Korea). Chonnam National University Herbarium provided the SHE used in the experiment (No. JNU CM20180908). 3,5-DCQA, one of the components of S. herbacea, was purchased from Enzo Life Sciences (Farmingdale, NY, USA).
Preparation of murine macrophages
Wild-type female C57BL/6J mice were purchased at 8 weeks of age (21–23 g body weight) from Dae Han Bio Link Co., Ltd. (Eumseong, Korea). After sacrificing mice by cervical dislocation, macrophages were obtained from the femurs and tibias. All animal studies were conducted with the approval of the institutional animal care and use committee of Konyang University (IACUC P-19-22-E-01). Bone marrow-derived macrophages (BMDMs) from murine were prepared as described previously. 12
In brief, BMDMs were cultured in a complete Iscove's Modified Dulbecco's Medium (Thermo Fisher Scientific, Waltham, MA, USA) including 30% L929 cell culture supernatant, 1% sodium pyruvate (Thermo Fisher Scientific), 1% Eagle's minimum essential medium nonessential amino acids (NEAAs; Thermo Fisher Scientific), 1% penicillin/streptomycin (Thermo Fisher Scientific), and 10% fetal bovine serum (FBS) (Corning, Manassas, VA, USA) in a 5% CO2 incubator at 37°C for 6 days. The cells were seeded in 48-well plates (2 × 105 cells/well) or in 6-well plates (2 × 106 cells/well) or 35-mm confocal dishes (SPL Life Sciences, Pocheon, South Korea; 2 × 106 cells/well).
Culture of human trophoblasts
Swan 71 cells (immortalized human first-trimester trophoblast cell line) were received from Dr. Gil Mor (Wayne State University, School of Medicine, Detroit, MI, USA). Swan 71 cells were cultured in Dulbecco's modified Eagle medium (DMEM) (Welgene, Gyeongsangbuk-do, Korea) containing 10% FBS, 1% penicillin/streptomycin, 1 mM hydroxyethyl piperazine ethane sulfonic acid (Thermo Fisher Scientific), 1% NEAA, and 1 mM sodium pyruvate. The cells were seeded in 48-well plates (2 × 104 cells/well) or in 6-well plates (2 × 105 cells/well).
Cell stimulation with LPS and adenosine triphosphate and SHE treatment
Activation of the NLRP3 inflammasome is activated by signal 1 and signal 2, and we investigated where SHE is regulated. First, the activation of the NLRP3 inflammasome was triggered by LPS (100 ng/mL, 6 h, signal-1 inducer) and adenosine triphosphate (ATP) (5 mM, 30 min, signal-2 inducer). And then SHE was treated 2 h before (pretreatment) or 6 h after (delayed treatment) LPS stimulation. ATP was added last 30 min in each experiment.
Measurement of cytokine levels
IL-6, tumor necrosis factor alpha (TNF-α), and IL-1β secreted from cells were measured using an ELISA kit (R&D Systems, Minneapolis, MN, USA).
Western blotting
Proteins were isolated in lysis buffer with protease inhibitor cocktail (Roche Diagnostics Deutschland GmbH) and were separated by 12% or 15% sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE and then electrotransferred onto a polyvinylidene difluoride microporous membrane (Millipore, Bedford, MA, USA). The membranes were incubated with the primary antibody using antimouse caspase-1 (AdipoGen, San Diego, CA, USA), pro and cleaved forms of antimouse IL-1β (R&D Systems), antirabbit cyclooxygenase (COX)-2 (Cell Signaling Technology, Danvers, MA, USA), and anti-β-actin (Santa Cruz Biotechnology, Inc., Dallas, TX, USA) overnight at 4°C.
The membranes were washed and incubated with the corresponding secondary antibody (Cell Signaling Technology) for two hours at room temperature. Proteins were then detected using a chemiluminescence kit (Thermo Fisher Scientific) and blot images were captured using a Fusion Solo S ChemiDoc (Vilber, Collégien, France).
Lactate dehydrogenase cytotoxicity assay
The level of lactate dehydrogenase (LDH) released from cells was analyzed using the LDH Cytotoxicity Assay Kit (DoGenBio, Seoul, Korea) and measured according to the manufacturer's instructions. Absorbance was measured immediately at 450 nm using a plate reader (BioTek Instruments, Inc., Winooski, VT, USA). The cytotoxicity was calculated using the following formula. LDH release (%) = (A − B/C − B) × 100 (where A is experiment background control, B is low control, and C is high control).
Cell viability assay
The effect of SHE on cell viability was measured at 450 nm in a plate reader (BioTek Instruments, Inc., Highland Park, USA) using a colorimetric EZ-CyTox kit (Daeil Lab., Seoul, Korea) according to the manufacturer's instructions. The cell viability was calculated using the following formula. Cell viability (%) = (experiment group/control group) × 100.
ASC oligomerization
ASC oligomerization visualization was performed according to Ref. 13 BMDMs were seeded in six-well plates and inflammasome was activated by LPS and ATP stimulation in the absence or presence of SHE. Rinse the cells with ice-cold PBS and use a scraper to remove the cells, then transfer to a 1.5 mL microcentrifuge tube and centrifuge the cells at 2000 g for 10 min at 4°C. Remove the supernatant and lyse the cells by adding 500 μL of ice-cold lysis buffer to the cell pellet and shearing 10 times using a 21-gauge needle. Here, 50 μL of the lysate was mixed with 5 × sample loading buffer and used to detect antirabbit ASC (Cell Signaling Technology) and anti-β-actin.
Centrifuge the remaining lysate at 5000 g for 10 min at 4°C and transfer the supernatant to a fresh tube. Add 2 mM disuccinimydyl suberate (Thermo Fisher Scientific) to the supernatant and incubate for 30 min at room temperature with agitation using a rotator, then centrifuge for 10 min at 5000 g at 4°C. The pellet was a cross-linked pellet and resuspended in lysis buffer and 50 μL of 5 × sample loading buffer and boiled at 95°C for 10 min. The cross-linked pellets were separated on 12% SDS-PAGE and ASC oligomerization was confirmed by Western blotting using antirabbit ASC.
Confocal microscopy
The BMDMs were fixed in 4% paraformaldehyde for 10 min and permeabilized with 0.1% Triton X-100 for 10 min. They were then incubated for 1 h in 0.1% BSA at room temperature. The cells were then incubated with the primary antibody using antirabbit ASC (Cell Signaling Technology) and antigoat NLRP3 (Abcam, Cambridge, United Kingdom) overnight at 4°C. Secondary antibodies of Alexa Fluor® 647 donkey antirabbit lgG (H+L) and Alexa Fluor 488 donkey antigoat IgG (H+L) were stained for 1 h at 20°C. Finally, cells were stained with a mounting solution containing DAPI and analyzed using a confocal microscope (LSM 700; Zeiss, Jena, Germany) and ZEN 2012 black edition software.
cDNA synthesis and real-time quantitative polymerase chain reaction
The expression of NLRP3, ASC, IL-1β, IL-6, PTGES2, and β-actin genes of Swan 71 cells was determined by quantitative real-time polymerase chain reaction (RT-qPCR). The mRNA of cells was extracted using the easy-BLUE™ Total RNA Extraction Kit (iNtRON Biotechnology, Seongnam, Korea). For cDNA synthesis, 0.1 μg of RNA and ReverTra Ace® qPCR RT Master Mix (TOYOBO Bio-Technology, Osaka, Japan) were used and prepared according to the manufacturer's instructions. RT-qPCR was performed using an SYBR Green PCR Kit (TOYOBO Bio-Technology). β-actin was used to normalize expression levels.
The oligonucleotide primers were used as follow: NLRP3 forward: 5′-GATCTTCGCTGCGATCAACA-3′, reverse: 5′-GGGATTCGAAACACGTGCATTA-3′; ASC forward: 5′-ATCCAGGCCCCTCCTCAGT-3′, reverse: 5′-GTTTGTGACCCTCGCGATAAG-3′; IL-1β forward: 5′-CCAGGGACAGGATATGGAGCA-3′, reverse: 5′-TTCAACACGCAGGACAGGTACAG-3′; IL-6 forward: 5′-CCAGCTATGAACTCCTTCTC-3′, reverse: 5′-GCTTGTTCCTCACATCTCTC-3′; PTGES2 forward: 5′- CCAGTGGCCTGGTAATCACT-3′, reverse: 5′-GCACAGACCAGAAAACACCA-3′; β-actin forward: 5′-CACCAACTGGGACGACAT-3′, reverse: 5′-ACAGCCTGGATAGCAACG-3′. RT-qPCR amplification was performed using a LightCycler® 96 real-time PCR system (Roche, Mannheim, Germany) for 60 sec at 95°C, 10 sec at 95°C, 30 sec at 58°C, and 60 sec at 72°C.
Statistical analysis
The differences between groups were determined by one-way analysis of variance (ANOVA) with Tukey's multiple comparisons or two-way ANOVA followed by the Bonferroni post hoc test for multigroup comparisons (GraphPad Prism 5; GraphPad Software, Inc., La Jolla, CA, USA). P < .05 was considered statistically significant.
Results
SHE inhibits NLRP3 inflammasome activation in macrophage stimulated through signal 1 and signal 2
To determine the effects of SHE on cell viability, BMDMs were treated with different concentrations of SHE (0.5–20 mg/mL), and then a water-soluble tetrazolium salts assay was performed. SHE showed no significant cytotoxic effects at concentrations of 0.5–5 mg/mL, but the cell viability was decreased at concentrations of 10–20 mg/mL (Supplementary Fig. S1). The nontoxic concentration at 1, 2.5, and 5 mg/mL of SHE was used throughout this study.
To block signal 1 and signal 2, SHE has been treated before LPS and ATP stimulation (Fig. 1A). The production of IL-6, TNF-α, and IL-1β were significantly upregulated by stimulation with LPS and ATP in BMDMs (Fig. 1B-D), and these increases were significantly inhibited by pretreatment with SHE. Next, we assessed the change in protein expression related to NLRP3 inflammasome activity by SHE. The pretreatment of SHE reduced the cleavage of IL-1β and caspase-1 in a dose-dependent manner (Fig. 1E). Collectively, these results indicate that SHE pretreatment can effectively downregulate NLRP3 inflammasome activation in BMDMs stimulated through signal 1 and signal 2.
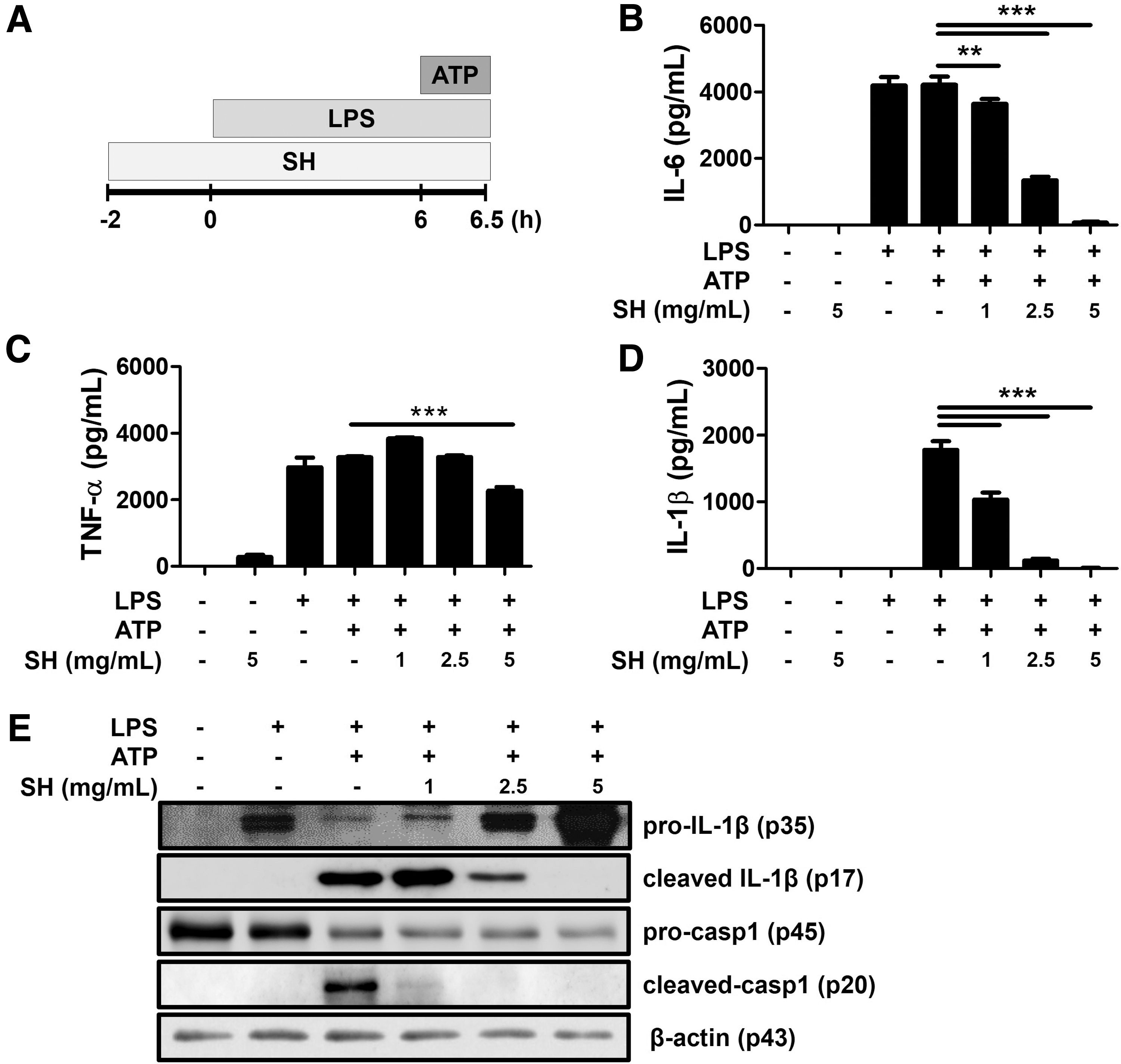
SHE downregulates NLRP3 inflammasome-mediated production of IL-6, TNF-α, and IL-1β in BMDMs.
SHE significantly inhibits NLRP3 inflammasome activation in macrophage stimulated through signal 2
Next, we investigated whether SHE inhibited the activation of the NLRP3 inflammasome when it blocked signal 2 in BMDMs. LPS-primed BMDMs were treated with SHE, followed by ATP treatment (Fig. 2A). Delayed treatment of SHE significantly inhibited IL-1β secreted by NLRP3 inflammasome activation. However, the production of IL-6 and TNF-α from these macrophages was not affected by SHE treatment (Fig. 2B–D). This can be explained by the fact that LPS, a signal-1 inducer, already stimulated macrophages, and delayed SHE treatment could affect the signal-2 pathway only. Delayed treatment of SHE effectively blocked IL-1β and caspase-1 cleavage induced by LPS and ATP stimulation (Fig. 2E). These results demonstrated that delayed treatment of SHE inhibited NLRP3 inflammasome activation in LPS-primed BMDMs.

SHE directly suppresses NLRP3 complex formation in BMDMs and inhibits IL-1β production but not IL-6 and TNF-α production.
SHE inhibits pyroptosis induced by NLRP3 inflammasome activation in macrophages
NLRP3 inflammasome activation induces pyroptotic cell death (a rapid inflammatory form of lytic programmed cell death). 4 Therefore, we investigated whether SHE modulates the effect of pyroptotic cell death in BMDMs. LPS and ATP treatment resulted in pyroptotic cell death, which indicates that both SHE pretreatment and delayed treatment of BMDM increased cell viability and decreased LDH release (Fig. 3A–D). These results exhibited that pretreated and delayed treatment of SHE suppresses NLRP3 inflammasome-mediated pyroptotic cell death by regulation of signal-1 and signal-2 pathways.

SHE inhibits cell death and LDH release induced by NLRP3 inflammasome activation in BMDMs.
SHE inhibits ASC oligomerization and speck formation in macrophages
Activated caspase-1 is induced through the assembly of an inflammasome complex composed of oligomerized ASC, NLRP3, and caspase-1. 4 In addition, ASC also forms dimers or oligomers under conditions where inflammation is activated and assembled into large protein platforms called “speck.” 4,14 We investigated whether SHE inhibits ASC oligomerization and speck formation in BMDM, which is considered a hallmark of inflammasome formation. ASC oligomerization and ASC speck induced by LPS and ATP were reduced by SHE pretreatment and delayed treatment (Fig. 4A, B). This finding suggests that SHE suppresses NLRP3 inflammasome activation through the inhibition of ASC oligomerization.
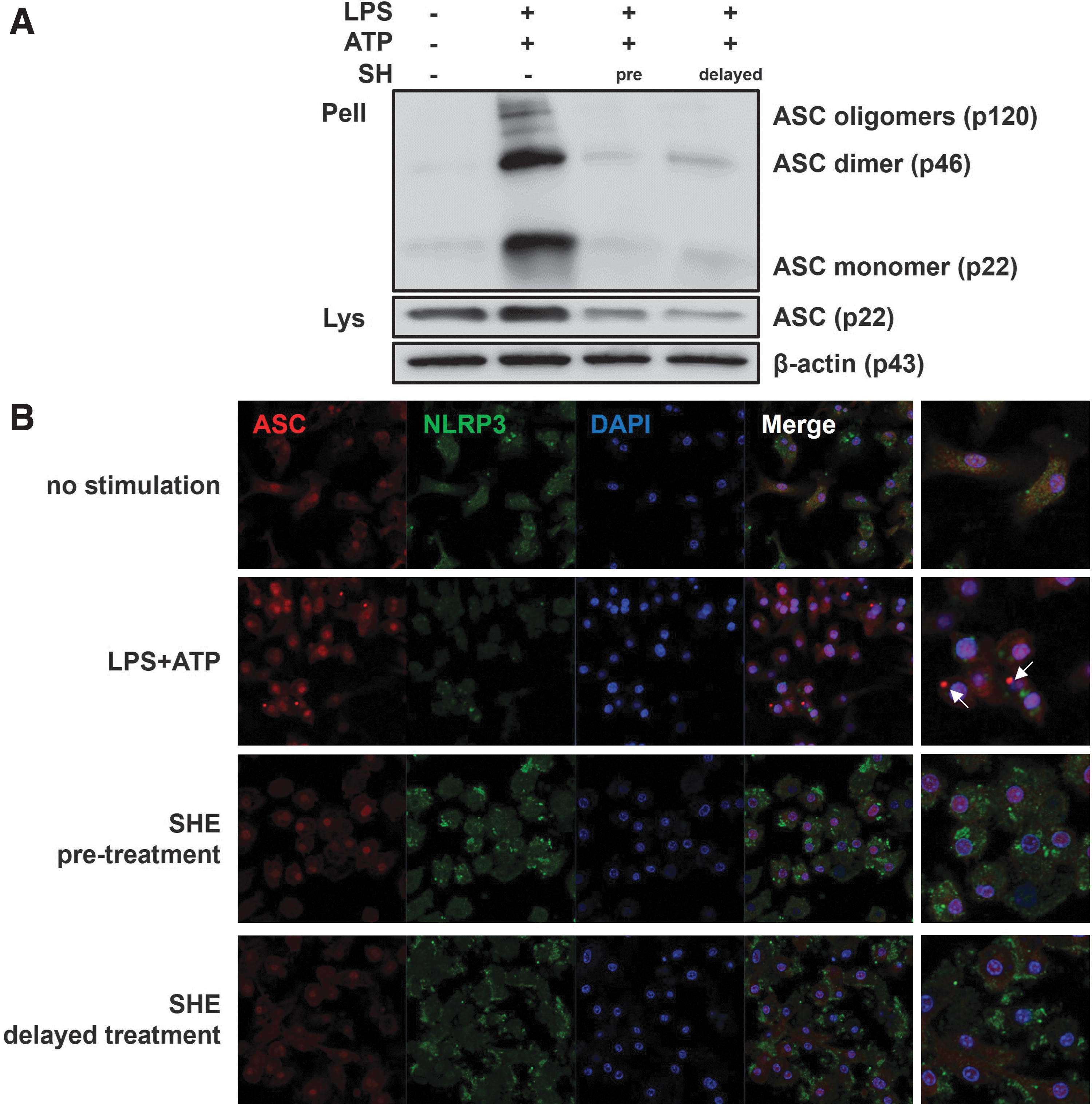
SHE suppressed the oligomerization and speck formation of ASC in BMDMs.
SHE inhibits NLRP3 inflammasome activation in trophoblasts
We investigated whether SHE modulates NLRP3 inflammasome activity in the trophoblast. Because LPS and ATP cotreatment caused Swan 71 cell death but only LPS did not, Swan 71 cells were treated with LPS only for RT-qPCR study. SHE pretreatment significantly reduced the expression of NLRP3, ASC, IL-1β, and IL-6 mRNA in LPS-stimulated Swan 71 cells (Fig. 5A–D). In addition, to determine whether SHE regulates proinflammatory cytokine production by NLRP3 inflammasome activity in the trophoblast, we investigated pretreatment of SHE in Swan 71 cells (Fig. 5E, F). The productions of IL-6 and IL-1β were significantly downregulated by SHE in a dose-dependent manner. Collectively, SHE downregulated NLRP3 inflammasome activation in trophoblasts.
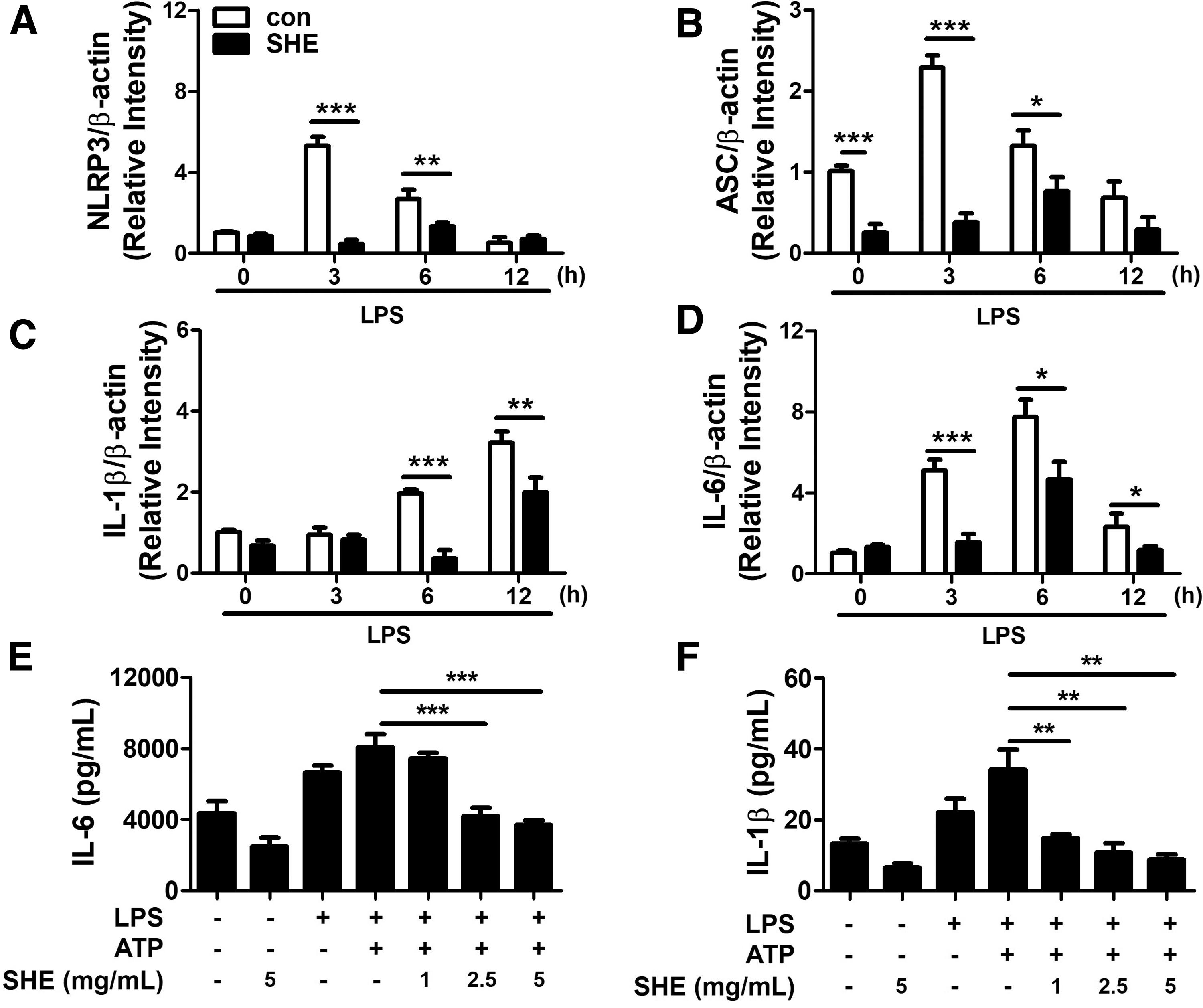
SHE inhibits NLRP3 inflammasome activation in Swan 71 cells.
SHE inhibits the expression of COX-2 and prostaglandin E2 induced by IL-1β stimulation in trophoblasts.
The expression of COX-2 is increased by NLRP3 inflammasome-derived IL-1β and enhances prostaglandin E2 (PGE2). 15,16 To demonstrate the effect of SHE on COX-2 and PGE2 expression in trophoblasts, we pretreated Swan 71 cells with various doses of SHE and then stimulated them with IL-1β. We showed that the expression of COX-2 in the Swan 71 cells was increased by IL-1β stimulation and dose-dependently inhibited by SHE pretreatment (Fig. 6A). IL-1β dose-dependently increased the PTGES2 mRNA expression in SHE-untreated Swan 71 cells (Fig. 6B). SHE-pretreatment Swan 71 cells significantly inhibited the increase of PTGES2 mRNA stimulated by IL-1β (Fig. 6C).
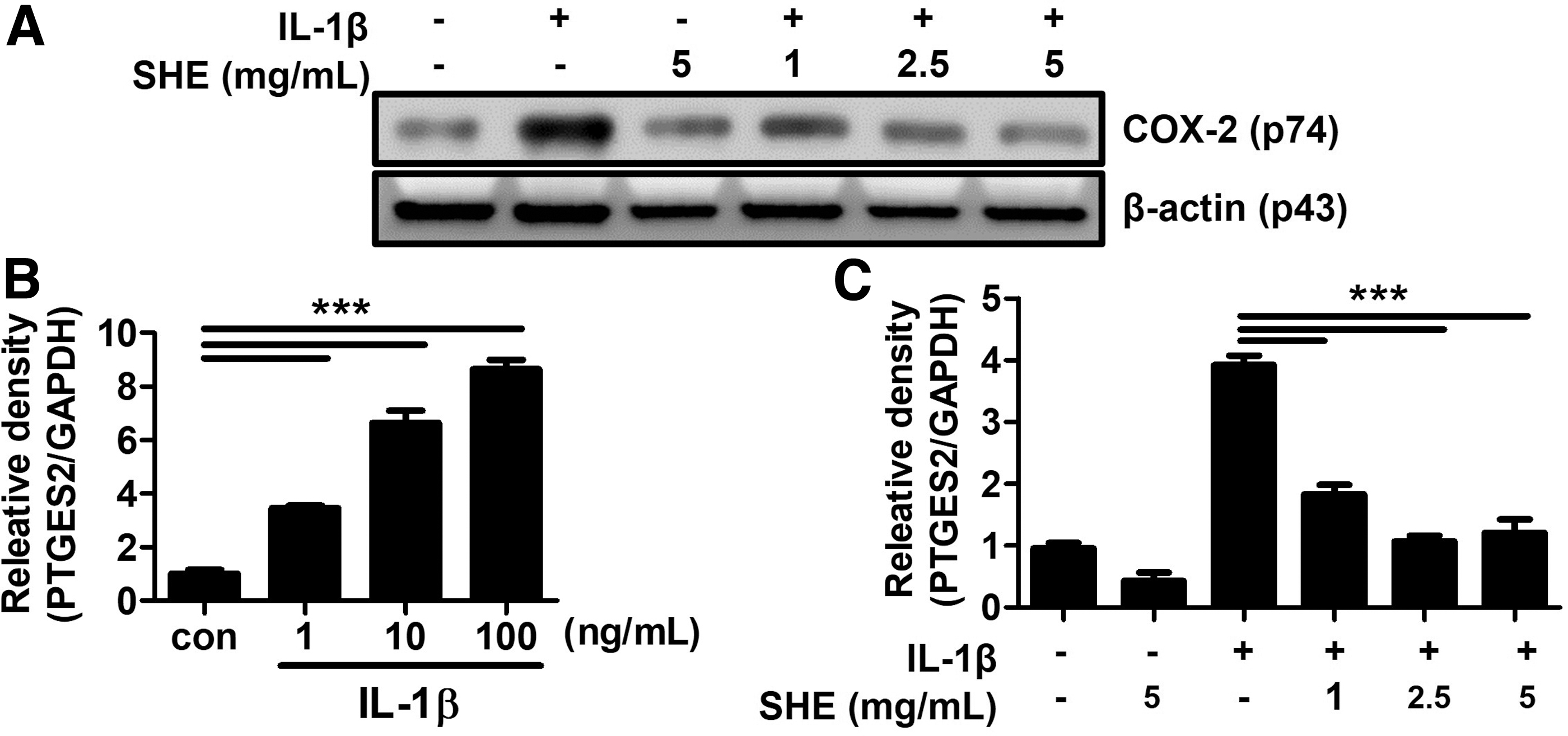
SHE inhibits the expression of COX-2 and PTGES2 mRNA induced by IL-1β in Swan 71 cells.
3,5-DCQA inhibits the production of IL-1β induced by IL-1β stimulation in trophoblasts.
We have identified 12 caffeoylquinic acid derivatives and 3 flavonol glycosides as major components of S. herbacea in previous studies. 10,11 We previously found that 3,5-DCQA, one of the components of S. herbacea, has antioxidant effects. 10,11 In addition, 3,5-DCQA decreased the expression of inflammation (inducible nitric oxide synthase, TNF-α, and COX-2) by LPS stimulation in RAW264.7 cells. 17 We investigated whether 3,5-DCQA regulates IL-1β production in NLRP3 inflammasome-induced macrophages. BMDMs pretreated with 3,5-DCQA significantly inhibited the increase in IL-1β production by LPS and ATP stimulation (Fig. 7). Therefore, among the caffeoylquinic acids constituting S. herbacea, 3,5-DCQA inhibited IL-1β produced by NLRP3 inflammasome activity.
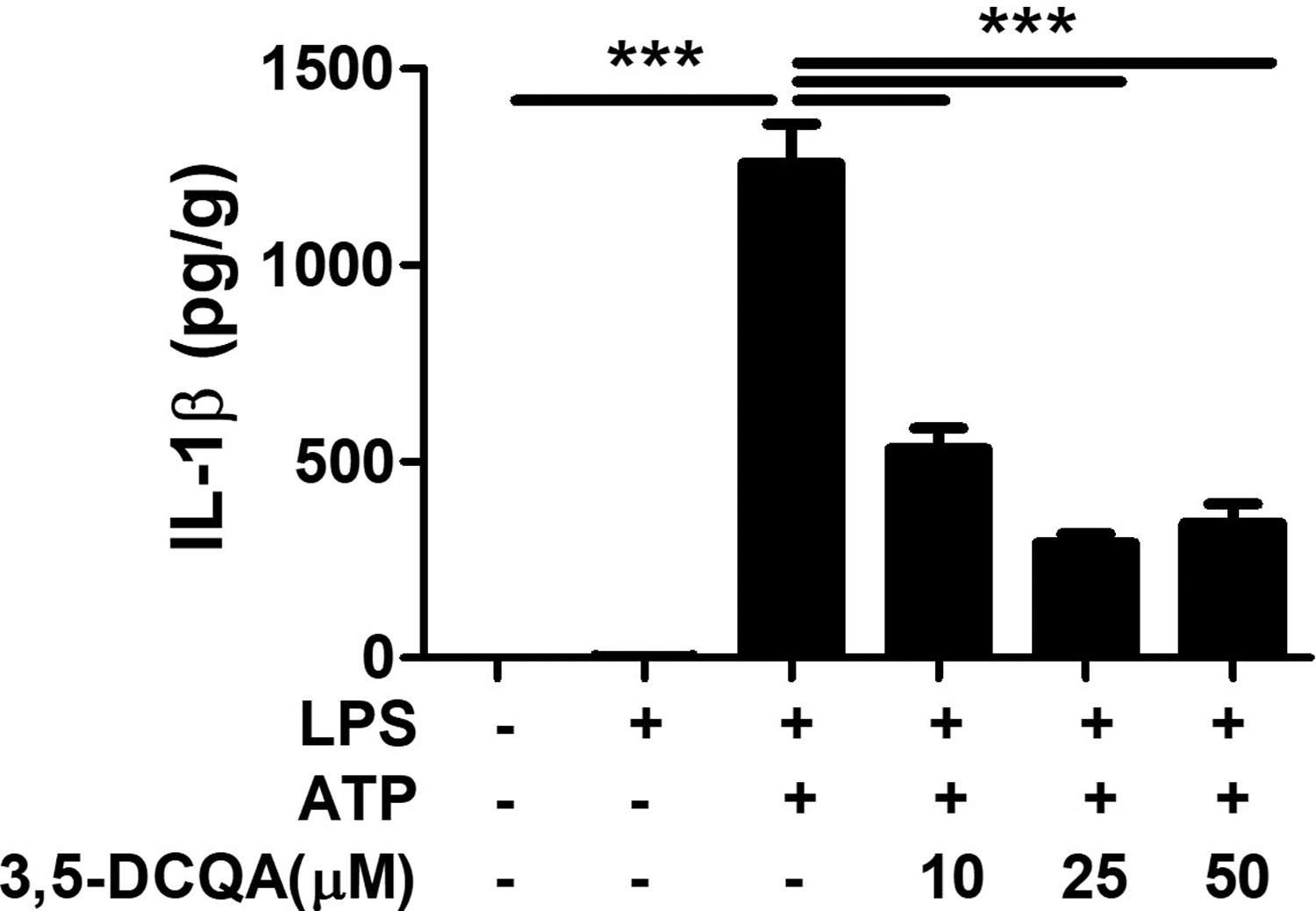
3,5-DCQA inhibits IL-1β production by NLRP3 inflammasome activation in BMDM. BMDMs were pretreated with indicated doses of 3,5-DCQA for 2 h and treated with LPS (100 ng/mL) for 6 h and subsequently treated with ATP (5 mM) for 30 min. The levels of IL-1β in culture supernatants were determined by ELISA. Data were analyzed by one-way ANOVA. ***P < .001. 3,5-DCQA, 3,5-dicaffeoylquinic acid.
Discussion
Inflammation during pregnancy was associated with various pregnancy complications, including miscarriage, preterm birth, and preeclampsia. 18 –22 The expression of NLRP3 inflammasome increased in the placenta of women with severe preeclampsia and the endometrium of women with recurrent miscarriages. 21,22 Therefore, novel therapeutic candidates are needed to control NLRP3 inflammasome activation and inflammation-related pregnancy complications. SHE has been widely used in traditional herbal medicines and foods and has received considerable attention to treat intestinal disorders, diabetes, obesity, and hepatitis. 8,9 However, the efficacy of SHE on pregnancy complications has not been established. Our study showed that SHE can regulate NLRP3 inflammasome activation and IL-1β production in macrophages and trophoblasts.
Intrauterine macrophages play an essential role in early implantation and fetal development by maintaining local immune balance. 23,24 Macrophages are important immune cells that regulate immune responses during pregnancy, and many studies have been reported on the NLRP3 inflammasome. 23,25 –27 Our results showed that LPS- and ATP-induced NLRP3 inflammasome activity in macrophages significantly inhibited NF-κB activity and NLRP3 inflammasome activity with SHE pretreatment. In addition, a delayed SHE treatment experiment was performed to confirm the effect of suppressing signal 2 after signal-1 stimulation.
Delayed treatment did not affect the release of IL-6 and TNF-α and inhibited IL-1β synthesis, suggesting that delayed-SHE treatment could specifically modulate signal 2 of NLRP3 inflammasome activation. Collectively, SHE inhibited not only the priming (signal 1) but also the activation (signal 2) step of NLRP3 inflammasome activity in macrophages.
We further investigated the mechanisms of NLRP3 inflammasome activation and cell damage.
The ASC, which is involved in NLRP3 inflammasome activation, recruits and activates caspase-1 by triggering supramolecular oligomerization into large intertwined fibrils called “ASC-speck.” 28 In previous studies, artemisia princeps extract and pumpkin tendril extract inhibited ASC oligomerization induced by ATP, monosodium urate, silica, and nigericin-induced NLRP3 inflammasome activation. 29,30 In line with them, our result confirmed that ASC oligomerization and speck were increased in LPS-primed and ATP-activated BMDMs and dose-dependently inhibited by SHE. These results indicate that SHE can inhibit NLRP3 inflammasome activation through the regulation of ASC oligomerization.
Pyroptosis, a type of cell death, induces membrane rupture after cell membrane pore formation and cell expansion and releases intracellular contents such as LDH out of the cell. 31 Our results showed that SHE inhibited pyroptotic cell death by inhibiting LDH release and increased cell viability.
The placental trophoblasts are critical for a successful pregnancy by mediating essential steps such as implantation, production of pregnancy hormones, fetus protection, increased maternal blood flow to the placenta, and delivery. 32,33 The placental trophoblast contains TLRs and nucleotide-binding oligomerization domain-like receptors and can produce proinflammatory cytokines such as IL-1β and IL-6. 34 –36 Elevated IL-1β in the trophoblast has been associated with pregnancy complications such as miscarriage, preeclampsia, and preterm birth. 37 –40
Some studies observed that Swan 71 cells secreted IL-1β by activating the NLRP3 inflammasome by monosodium urate, LPS, and hyperglycemia. 20,29,41 We also demonstrated that NLRP3 inflammasome activation was increased in LPS- and ATP-activated trophoblasts and inhibited by SHE. Similarly, pumpkin tendril extract and TANK-binding kinase 1 have controlled NLRP3 inflammasome activation in trophoblast. 29,42
Prostaglandins are one of the important mediators of inflammatory and immune responses induced by various stimuli, including LPS, cytokines, and phagocytosis. 43 Among them, PGE2 is produced by the inducible enzyme COX-2 and is involved in the initiation of labor. 16,44 COX-2 is associated with an inflammatory factor that induces activation of the NLRP3 inflammasome, and an increase in COX-2 expression leads to preterm birth and preeclampsia. 45,46
In a previous study, high levels of PGE2 and IL-1β were observed in the gingival crevicular and blood serum of preterm birth patients with periodontal disease. 47 However, there are few studies on whether IL-1β, the final product of inflammasome activity in the trophoblast, is involved in the expression of PGE2. Therefore, we experimented to prove it and found that the expression of COX-2 and PGE2 was increased in IL-1β-stimulated Swan 71 cells, which was inhibited by SHE.
Finally, we investigated which component is a key molecule of SHE in anti-inflammatory response. The 3,5-DCQA, one of the components contained in SHE, is known to be involved in the inhibition of inflammation. 11 This 3,5-DCQA, which is also contained in the leaves of Ligularia fischeri, has been reported to be effective for antioxidant and anti-inflammatory activity. 17 Our study also confirmed that 3,5-DCQA inhibited IL-1β production through regulation of NLRP3 inflammasome activity. Therefore, not only SHE but also 3,5-DCQA showed anti-inflammatory effects, suggesting that the diseases caused by NLRP3 inflammasome activation can be prevented or controlled by SHE.
However, this study has a limitation of the in vitro experiment, so the following study should be conducted in animal disease models and clinical trials to confirm the efficacy and safety.
In summary, SHE significantly inhibited the activation of NLRP3 inflammasome and IL-1β secretion in macrophages and trophoblasts (Fig.8). These findings suggest that SHE may be a potential candidate for the prevention and treatment of NLRP3 inflammasome-mediated pregnancy complications such as preterm birth, preeclampsia, and recurrent pregnancy loss.

Schematic summary of SHE-mediated NLRP3 inflammasome regulation in macrophages and trophoblasts. SHE inhibits the production of IL-6 and TNF-α and the expression of pro-IL-1β and NLRP3 induced by LPS priming (signal 1). SHE decreases the expression of COX-2 and PGE2 triggered by IL-1β. SHE inhibits caspase-1 activity, IL-1β production, and pyroptosis through NLRP3 inflammasome activation (signal 2). As a result, SHE downregulates NLRP3 inflammasome activation and IL-1β production.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by a grant from the Korean Health Technology R&D project from the Ministry of Health & Welfare, Republic of Korea (HI17C1238) and the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2017R1A6A1A03015713).
Supplementary Material
Supplementary Figure S1