
Abstract
The abnormal change of vascular smooth muscle cell (VSMC) behavior is an important cellular event leading to neointimal hyperplasia in atherosclerosis and restenosis. Plantamajoside (PMS), a phenylethanoid glycoside compound of the Plantago asiatica, has been reported to have anti-inflammatory, antioxidative, and anticancer activities. In this study, the protective effects of PMS against intimal hyperplasia and the mechanisms underlying the regulation of VSMC behavior were investigated. MTT and BrdU assays were performed to evaluate the cytotoxicity and cell proliferative activity of PMS, respectively. Rat aortic VSMC migrations after treatment with the determined concentration of PMS (50 and 150 μM) were evaluated using wound healing and Boyden chamber assays. The inhibitory effects of PMS on intimal hyperplasia were evaluated in balloon-injured (BI) rat carotid artery. PMS suppressed the proliferation in platelet-derived growth factor-BB-induced VSMC, as confirmed from the decrease in cyclin-dependent kinase (CDK)-2, CDK-4, cyclin D1, and proliferating cell nuclear antigen levels. PMS also inhibited VSMC migration, consistent with the downregulated expression and zymolytic activities of matrix metalloproteinase (MMP)2, MMP9, and MMP13. PMS specifically regulated MMP expression through p38 mitogen-activated protein kinase and focal adhesion kinase pathways. Tissue inhibitor of metalloproteinase (TIMP)1 and TIMP2 levels were upregulated via Smad1. TIMPs inhibited the conversion of pro-MMPs to active MMPs. PMS significantly inhibited neointimal formation in BI rat carotid arteries. In conclusion, PMS inhibits VSMC proliferation and migration by upregulating TIMP1 and TIMP2 expression. Therefore, PMS could be a potential therapeutic agent for vascular atherosclerosis and restenosis treatment.
Introduction
Alterations in the behavior of vascular smooth muscle cells (VSMCs) are a major factor that contributes to the process of cardiovascular disease, including atherosclerosis, neointimal hyperplasia, and myocardial infarction. 1,2 Cellular and phenotypic changes in VSMCs are mainly driven by abnormally elevated or modified levels of paracrine stimuli such as cytokines and growth factors secreted from inflammatory cells. 3 Among them, platelet-derived growth factor (PDGF) is considered the most potent mitogen regulating VSMC behavior. 4,5 The expression levels of PDGF-BB among the five isoforms were significantly higher in human atherosclerotic plaques than in normal arteries. 6 In addition, PDGF-BB-induced VSMC migration has been reported to lead to neointimal hyperplasia in carotid artery-injured animal models. 7,8
Plantamajoside (PMS), a bioactive component of Plantago asiatica, has been applied as a traditional plant medicine in East Asia and is shown to have medicinal properties. 9 Evidence suggests the beneficial effects of PMS treatment on various cell types and tissues. For instance, PMS was shown to modulate the advanced glycation end products-induced endothelial dysfunction by downregulating the expression of adhesion molecules and inflammatory cytokines. 10 In addition, PMS also exhibited protective effects against renal injury induced by cadmium through its antioxidative activity. 9 With respect to cardiovascular diseases, PMS was shown to attenuate isoproterenol-induced cardiac hypertrophy and hypoxia/reoxygenation-induced injury. 11,12
In particular, PMS repressed the growth and metastasis of malignant melanomas by promoting the expression of apoptotic genes and breast cancer by decreasing activities of matrix metalloproteinase (MMP)2 and MMP9. 13,14 Indeed, MMP2 and MMP9 are crucial mediators of VSMC migration and phenotypic changes, and PDGF-BB may increase the expression levels of MMPs in VSMCs, 15,16 leading to neointima formation. Despite these known beneficial functions of PMS in cancer and other diseases, its effect on VSMC behavior in neointima formation remains unknown. Here, we hypothesized that PMS may prevent or reverse the altered behaviors of PDGF-BB-induced VSMCs and inhibit intimal hyperplasia in balloon-injured (BI) rat carotid arteries by regulating MMPs or by unknown mechanisms.
In this study, we investigate the effects of PMS on VSMC proliferation and migration in vitro and neointimal hyperplasia in vivo and examine the mechanism involved in this process.
Materials and Methods
Materials
PMS (C29H36O16; molecular weight: 640.59; purity: high-performance liquid chromatography ≥95%) was obtained from Sigma–Aldrich (St. Louis, MO, USA), and its chemical structure is shown in Figure 1A. PDGF-BB and collagenase type I were obtained from Sigma–Aldrich. Elastase was purchased from Worthington Biochemical Co. (Lakewood, NJ, USA). Zoletil 50® was obtained from Virbac Corp. (Fort Worth, TX, USA). Fetal bovine serum and Dulbecco's modified Eagle's medium were obtained from WelGENE (Seoul, Korea). Rompun® was purchased from Bayer Corp. (Pittsburgh, PA, USA). A 2F balloon catheter was purchased from Edwards Lifesciences Corp. (Irvine, CA, USA).

PMS inhibits PDGF-BB-induced VSMC proliferation.
BrdU Cell Proliferation Colorimetric ELISA (enzyme-linked immunosorbent assay) Kit was obtained from Promega (Madison, WI, USA). Enhanced chemiluminescence (ECL) reagent was obtained from Animal Genetics Inc. (Tallahassee, FL, USA). TRIzol reagent was purchased from Qiagen (Germantown, MD, USA). The TF (transcription factor) Activation Profiling Plate Array Kit and Nuclear Extraction Kit were purchased from Signosis, Inc. (Sunnyvale, CA, USA). Bradford protein assay kit and siLentFect were obtained from Bio-Rad (Hercules, CA, USA). The HelixCript™ First-Strand cDNA Synthesis Kit and RealHelix™ qPCR Kit were obtained from NanoHelix (Seoul, Korea).
Animals
All animal experiments were strictly performed in compliance with the International Guidelines for the Care and Use of Laboratory Animals. The study protocol, No. CIACUC2020-S0032, was approved by the Chosun University Animal Care and Use Committee.
Primary culture of VSMCs
Rat aortic VSMCs were primarily isolated from the normal thoracic aorta of 6-week-old Sprague-Dawley male rats using a mixture of enzymes (1 mg/mL of collagenase type I and 0.5 mg/mL of elastase in phosphate-buffered saline [PBS]). 17 The isolated VSMCs were cultivated at 37°C in a 5% carbon dioxide incubator. Cells from passages 6 to 10 were used in this study.
Balloon injury model
Sprague-Dawley male rats (6- to 7-week-old, body weight 200 ± 50 g) were anesthetized with a mixture of Zoletil 50 (20 mg/kg) and Rompun (10 mg/kg), and then, a midline cervical incision was made in the exposed common carotid artery. A standard-sized (2F) balloon catheter was inserted into the incision and advanced to the aortic arch. Then, the inflated catheter to a volume of 0.02 mL was slowly withdrawn until the catheter was close to the arteriotomy incision for denuding the endothelium. After repeating this step three times, the external carotid artery was ligated and the wound was closed. The PMS dosage for rats was established in preliminary experiments to determine the minimum effective concentration of PMS without exhibiting toxicity in rat organs, including the lung, liver, and kidney. Therefore, 150 μL of PMS (3.3 mg/kg) was intraperitoneally administered to the treatment group of animals for 14 days. The same volume of PBS was injected into a sham group on the same days with PMS injection.
Cell proliferation assay
VSMCs were seeded onto a 96-well plate and cultivated until they reached 80% confluence. After 24 h treatment of cells with PDGF-BB and/or PMS, the proliferative activity of cells was measured using the BrdU incorporation assay.
Cell migration assay
The migratory activities of cells were evaluated by a two-dimensional (2D) scratch assay and a three-dimensional (3D) Boyden chamber assay. For the scratch assay, a rectangular region within the VSMCs was made using a cell scraper in each cultured well, and then, the cells were treated with PDGF-BB or PMS. After 48 h, the cells were fixed and stained, and then, the migratory activity of VSMCs was determined by the ratio of the wound closure area. For the Boyden chamber assay, cells (5 × 103/100 μL) were seeded on the upper chambers coated with 1% of gelatin solution on the lower surface. PDGF-BB or PMS were treated in each lower or upper chamber, respectively. After incubation for 8 h at 37°C, cells placed on the lower surface of the upper chamber were fixed with 100% methanol and stained with 1% Coomassie blue, and five random fields/membranes were counted.
Zymography
Diluted sample media were subjected to a 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) containing 0.8% of gelatin. Gels were washed with washing buffer to remove SDS (2.5% Triton X-100) for 1 h. For proteolysis, gels were incubated in gelatin incubation buffer (5 mM calcium chloride, 50 mM Tris-HCL, pH 7.5, 1 μM zinc chloride, 1% Triton X-100, 0.02% sodium azide) at 37°C for 48 h, and then, gels were stained with 1% Coomassie blue for 1 h and destained. The gels were dried, and the zymolytic areas were evaluated using the ImageJ Software (NIH, Bethesda, MD, USA).
TF analysis
The activity of the TFs was examined using the TF Activation Profiling Plate Array Kit. Briefly, VSMCs were seeded on a 60-mm plate at 4.0 × 105 cells per mL and starved for 24 h, and the cells were pretreated with PMS for 1 h, followed by treatment with 20 ng/mL of PDGF-BB for another 3 h. First, only the nuclei were extracted from the cultured cells using the Nuclear Extraction Kit. Then, 10 μg of the extracted nuclear protein was incubated with biotin-labeled probes to form a TF/DNA complex. After washing and elution, hybridization was performed in a 96-well plate by adding streptavidin–horseradish peroxidase and measuring the luminescence level with a luminometer (Microlumat Plus, Berthold, Germany).
Immunocytochemistry
VSMCs were seeded on the cover glass placed in a 24-well plate at 2 × 104 per well and incubated at 37°C for 24 h. The media was then replaced with a starvation medium for 24 h. Cells were then pretreated with PMS for 6 h, followed by treatment with 20 ng/mL of PDGF-BB for another 24 h. For the immunofluorescent labeling, cells were washed with cold 1 × PBS and fixed with cold 2% paraformaldehyde (PFA) in PBS for 15 min at room temperature (RT). Then, cells were washed with PBS and permeabilized or non-permeabilized for 15 min at RT with or without 0.5% Triton X-100 in PBS, respectively. The primary antibody was incubated overnight at 4°C in PBS with 2% bovine serum albumin, and then, the secondary antibody was incubated for 1 h at RT in the dark. After washing with PBS, the cell nuclei were stained with 0.2 μg/mL of DAPI, and the cover glass was mounted on the slide. Finally, cells were photographed with a laser scanning confocal microscope (Fluoview FV1000; Olympus, Tokyo, Japan).
Morphometric analysis
For morphometric analysis, the bilateral carotid arteries were excised and fixed in 4% PFA. The tissues were embedded in paraffin and cut into 5 μm sections for further hematoxylin and eosin (H&E) staining to assess the neointima area. The intimal and medial areas of the arterial specimens were evaluated using the ImageJ Software (NIH).
Immunohistochemistry
Paraffin-embedded sections were deparaffinized with xylene and rehydrated with serially diluted ethanol. Specimens were processed by the heat-induced epitope retrieval method for antigen retrieval. After blocking with 10% goat serum for 2 h, the sections were incubated with the primary antibodies such as tissue inhibitor of metalloproteinase (TIMP)1, TIMP2, MMP2, MMP9, and MMP13 (1:50 dilution; Abcam, Cambridge, MA, USA), and p-FAK(Y397) and p-FAK(Y925) (1:50 dilution; Cell Signaling, Danvers, MA, USA) for overnight at 4°C. The sections were washed with washing buffer and stained with secondary antibodies at RT for 1 h in dark. The nucleus was stained with 0.2 μg/mL of DAPI (Sigma–Aldrich) for 1 min. Finally, the specimens were photographed with a laser scanning confocal microscope (Fluoview FV1000; Olympus).
Immunoblot analysis
The tissue lysates of the carotid artery were prepared using RIPA buffer containing 1 mM phenylmethylsulfonyl fluoride and a protease inhibitor cocktail. Protein concentration was determined using the Bradford protein assay kit, and equal amounts of protein were subjected to SDS-PAGE and transferred to a polyvinylidene fluoride (PVDF) membrane. After incubation with blocking buffer (Tris-buffered saline–0.1% Tween 20, 5% skim milk) for 1 h at RT, the membrane was incubated overnight with primary antibodies at 4°C.
The primary antibodies were used at the following dilutions: MMP2 (1:4000), TIMP1 (1:1000), TIMP2 (1:2000), Talin (1:1000), MMP9 (1:1000), and integrin-β1 (1:10,000) (Abcam), protein kinase B (Akt, 1:4000), p-FAK(Y397) (1:500), p-Akt (1:2000), extracellular signal-regulated kinase (ERK, 1:3000), p-ERK (1:5000), P38 (1:2000), p-P38 (1:1000), Src (1:4000), p-FAK(Y925) (1:500), p-Src (1:2000), focal adhesion kinase (FAK, 1:1000), proliferating cell nuclear antigen (PCNA, 1:2000), and cyclin-dependent kinase-2 (CDK2, 1:1000) (Cell Signaling, Beverly, MA, USA), paxillin (1:2000; Millipore, Burlington, MA, USA), vinculin (1:1000; Santa Cruz, Dallas, TX, USA), MMP13 (1:1000; Novus, Centennial, CO, USA), Smad1 (1:1000; Abcam), and β-actin (1:10,000; Sigma–Aldrich). Then, the membrane was washed with washing buffer and incubated with the appropriate secondary antibodies for 1.5 h at RT. After washing five times, an ECL reagent was applied and the bands were detected.
Small-interfering RNA transfection
Single-stranded rat Smad1-specific sense and antisense RNA oligonucleotides were synthesized by Bioneer, and double-stranded RNA molecules were generated according to the manufacturer's instructions. The small-interfering RNA (siRNA) sequence was 5′- CCU GCU GGA UUG AGA UAC ACC UGC A -3′. For transfection, VSMCs were grown to 80% confluency and were then transfected with the Smad1 siRNA using siLentFect.
Real-time quantitative polymerase chain reaction
Total RNA was extracted by using TRIzol reagent. Total RNA was reverse transcribed to cDNA using a HelixCript First-Strand cDNA Synthesis Kit. Real-time quantitative polymerase chain reaction (qPCR) was performed with RealHelix SYBR Green qPCR Kit using an Applied Rotor-Gene 3000™ (Corbett Research, Sydney, Australia). The transcript levels of the genes of interest in all tested samples were normalized against the transcript levels of glyceraldehyde-3-phosphate dehydrogenase. The primer sequences for qPCR are listed in Table 1.
The Sequences of Primers Used for Real-Time Quantitative Polymerase Chain Reaction
bp, base pair; MMP, matrix metalloproteinase; TIMP, tissue inhibitor of metalloproteinase.
Statistical analysis
All quantified data were analyzed with GraphPad Prism 8.0 Software (La Jolla, CA, USA). Data are shown as mean ± standard deviation. Student's t-test was used for statistical comparisons between the two groups. One-way analysis of variance was used for statistical comparisons among multiple groups, followed by the Bonferroni post hoc test when the F statistic was significant. A two-tailed P < .05 was considered statistically significant.
Results
PMS inhibits PDGF-BB-induced VSMC proliferation
To determine the cytotoxicity of PMS, VSMCs were treated with serially diluted concentrations of PMS for 24 h. As shown in Figure 1B, 25–150 μM PMS treatment did not cause any alterations in the viability of VSMCs; however, at the 200 μM concentration, PMS induced significant cytotoxicity to VSMCs. Therefore, we evaluated the inhibitory effects of PMS on VSMC proliferation excluding the 200 μM concentration. As shown in Figure 1C, 50 and 150 μM were the minimum and maximum concentrations, respectively, showing the inhibitory effects on VSMC proliferation.
Although treatment with 100 μM was effective, no significant difference was observed from 50 μM. Therefore, we chose 50 and 150 μM for subsequent experiments. We further evaluated the altered expression levels of CDK2, cyclin D1, CDK4, and PCNA. We could observe that the expression levels of all examined proteins were dramatically increased by PDGF-BB, and this effect was significantly suppressed by PMS treatment in a dose-dependent manner (Fig. 1D). These results demonstrate that PMS suppresses VSMC proliferation by inhibiting the expression of cell cycle regulatory proteins.
PMS inhibits PDGF-BB-induced VSMC migration
To investigate the effects of PMS on PDGF-BB-induced VSMC migration, we performed 2D scratch assay and 3D Boyden chamber transwell assay. The scratch assay result showed that PDGF-BB significantly enhanced the migration of VSMCs compared with nontreated control cells and that PMS significantly abolished this effect (Fig. 2A). We also performed the Boyden chamber assay to validate this result, which confirmed that PMS significantly inhibited the PDGF-BB-induced VSMC migration in a dose-dependent manner (Fig. 2B). Thus, PMS attenuates the migration of PDGF-BB-stimulated VSMCs.
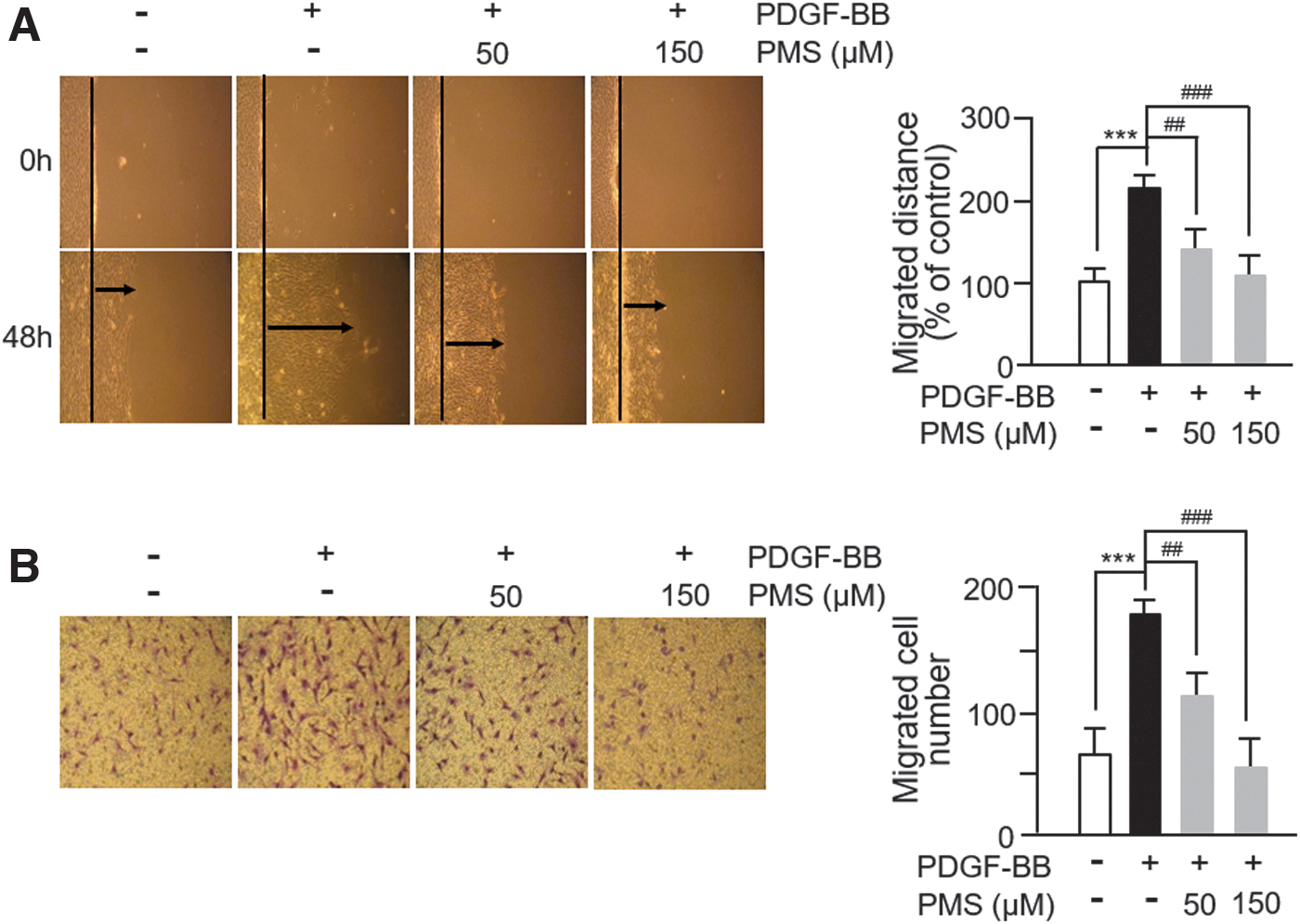
PMS inhibits PDGF-BB-induced VSMC migration.
PMS suppresses PDGF-BB-induced MMP expression and activity in VSMCs
MMPs participate in the process of the migration and invasion of various cell types, including tumors and VSMCs. 18 The zymolytic activities of MMP2 and MMP9 in the extracellular matrix (ECM) of the blood vessels are known to promote VSMC migration. 19 As expected, PDGF-BB significantly increased the expression levels of MMP2 and MMP9, and this effect was successfully abrogated by PMS treatment (Fig. 3A). The level of MMP13 also increased after PDGF-BB treatment, and PMS significantly suppressed the increased level of MMP13 (Fig. 3A). To confirm that PMS inhibits MMP expression in PDGF-BB-induced VSMCs, we examined the mRNA levels of MMPs in PDGF-BB/PMS-treated VSMCs. Consistent with the immunoblotting results, the mRNA levels of MMP2, MMP9, and MMP13 increased by PDGF-BB were significantly inhibited by PMS (Fig. 3B).
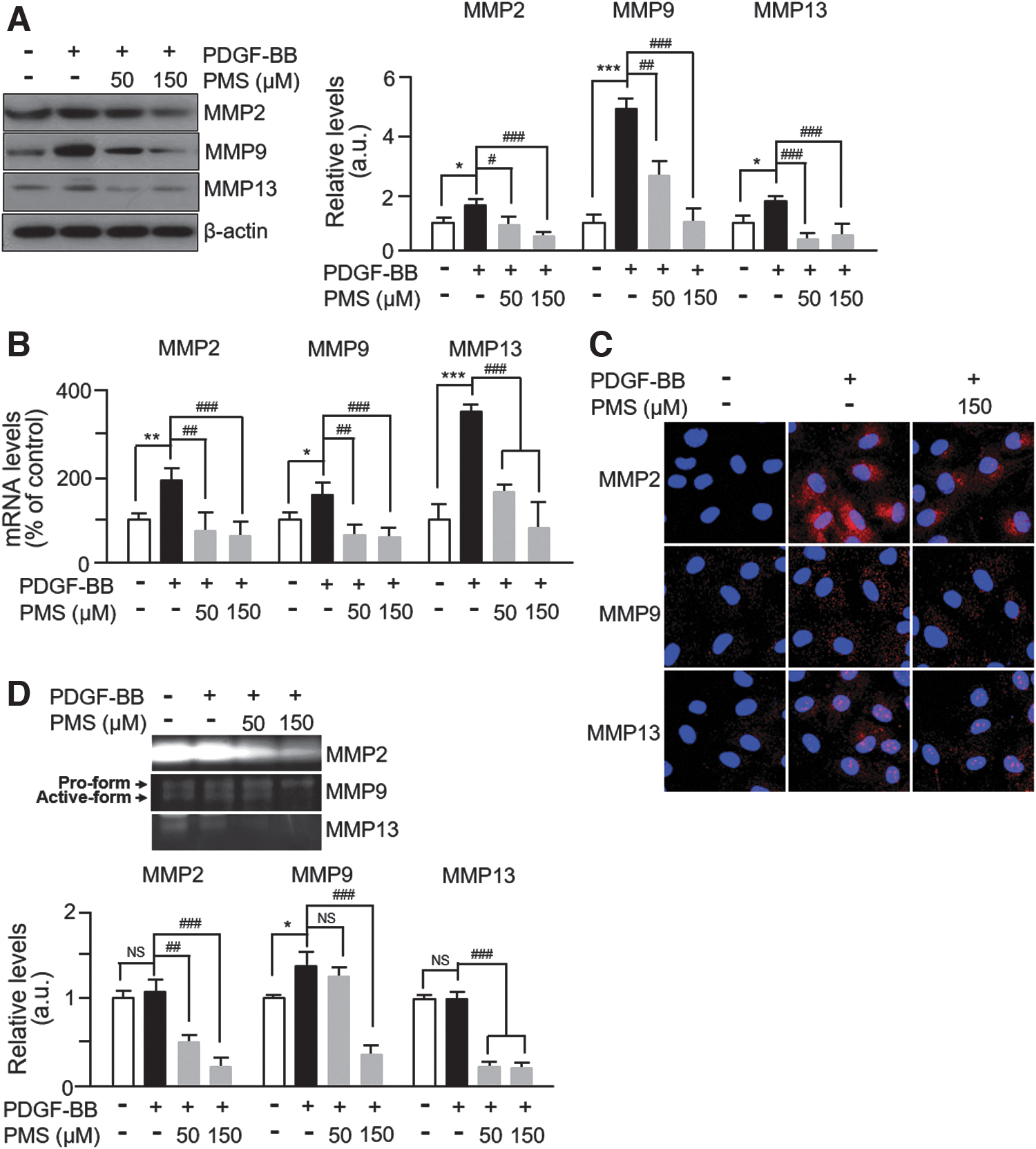
PMS suppresses PDGF-BB-induced MMP expression and activity.
In both cases, the levels of MMP7 remained unchanged (data not shown), suggesting that PMS specifically inhibits PDGF-BB-induced MMP expression. We also performed immunofluorescence staining under unpermeabilized conditions and MMP activity assay in PDGF-BB/PMS-treated VSMCs to investigate any alterations in their zymolytic activities. As shown in Figure 3C, the extracellular levels of MMP2, MMP9, and MMP13 increased in PDGF-BB-stimulated VSMCs compared with those in untreated control cells. However, PMS treatment downregulated the increased expression of MMP2, MMP9, and MMP13. Similar results of zymographic analyses were observed with the culture media from PDGF-BB/PMS-treated VSMCs (Fig. 3D). Together, these results demonstrate that PMS inhibits both MMP expression and activities in PDGF-BB-induced VSMCs.
PMS modulates tissue inhibitor of metalloproteinase (TIMP) expression via regulation of Smad1 factor in VSMCs
As evidence supports the regulatory effect of TIMP on MMP activities, 20 we investigated the alterations in TIMP levels in PDGF-BB/PMS-treated VSMCs. We observed that the levels of TIMP1 and TIMP2 were significantly reduced in PDGF-BB-treated cells, but PMS treatment increased their expression by over 5- and 3-fold, respectively, compared with untreated control cells and over 10-fold compared with PDGF-BB treatment (Fig. 4A). Unpermeabilized immunocytochemical staining showed that the extracellular levels of TIMP1 and TIMP2 were distinctly higher in PMS-treated VSMCs than in untreated control and PDGF-BB-treated cells (Fig. 4B).
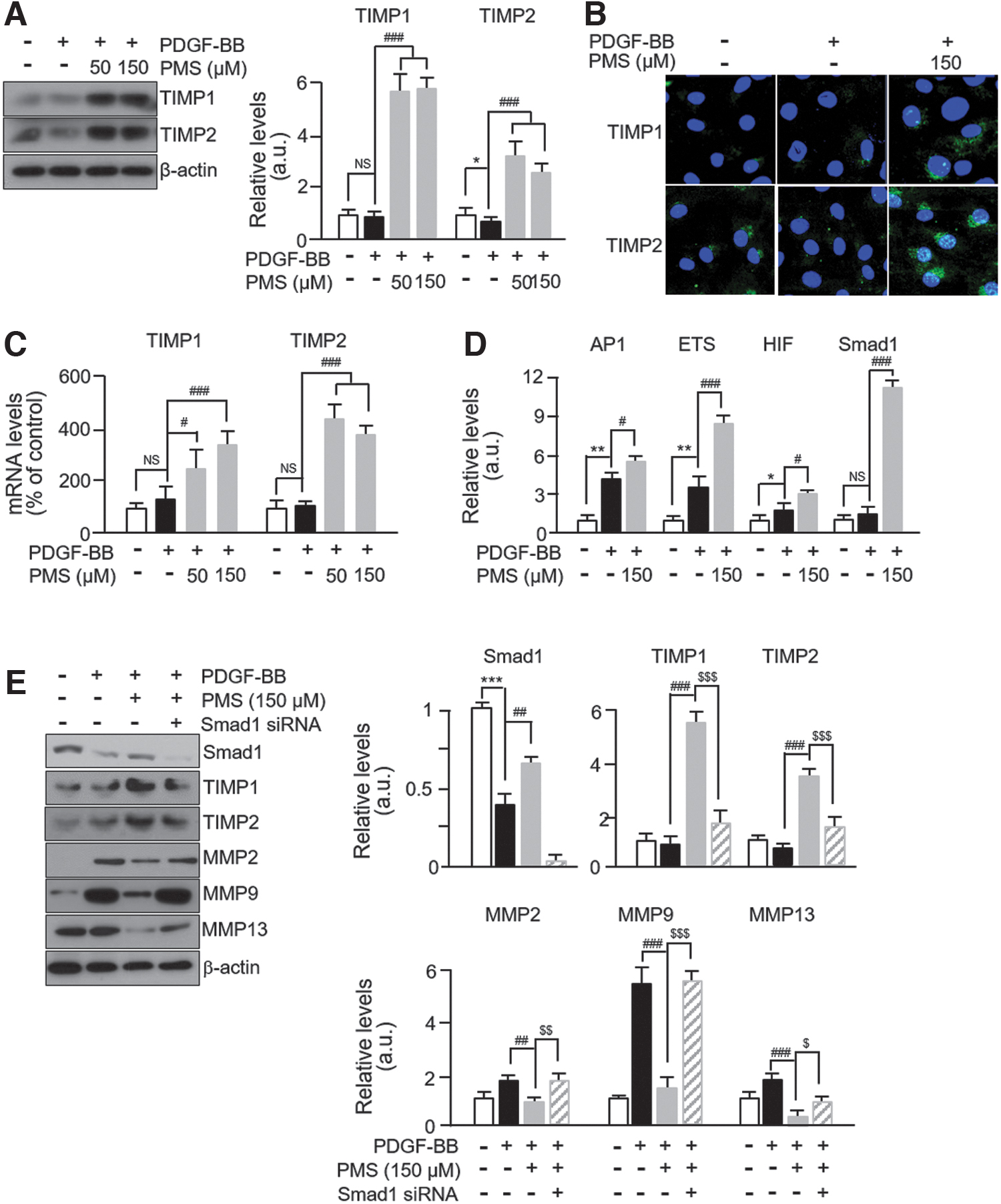
PMS modulates TIMP expression via regulation of Smad1 transcription factor.
We also observed a significant increase in the mRNA expression levels of TIMP1 and TIMP2 in PMS-treated cells compared with those in control and PDGF-BB-treated cells (Fig. 4C). The mRNA expression of TIMP1 and TIMP2 did not significantly decrease in PDGF-BB-treated cells compared with that in untreated controls. To further investigate the TF that participates in modulating TIMP expression, we performed a TF activation profiling assay. Among the 16 TFs tested, activator protein 1, E26 transformation specific family 1, hypoxia-inducible factor, and Smad1 were significantly upregulated in PMS-treated VSMCs compared with those in PDGF-BB-treated cells (Fig. 4D). The data on TF activities are presented in Table 2.
Altered Activities of Transcription Factors in Vascular Smooth Muscle Cells by Plantamajoside
VSMCs were treated with PDGF-BB at 20 ng/mL and then incubated with 150 μM PMS. Data are expressed as mean ± standard deviation compared with nontreated control cells (n = 3).
PDGF, platelet-derived growth factor; PMS, plantamajoside; VSMC, vascular smooth muscle cell.
Although there is no evidence that Smad1 is one of the TFs regulating TIMP expression, it is thought to participate in the regulatory mechanism of MMPs. 21 Therefore, we hypothesized that Smad1 is a key factor participating in TIMP regulation in PMS-treated VSMCs. As expected, the expression levels of Smad1 were significantly decreased in PDGF-BB-treated VSCMs, which were successfully rescued by PMS treatment. We also observed that the expression levels of TIMP1 and TIMP2 were not significantly altered after PMS treatment in Smad1-specific siRNA-transfected VSMCs. In addition, we observed that PMS had no effect on the levels of MMP2, MMP9, and MMP13 in Smad1 siRNA-transfected VSMCs (Fig. 4E). These results indicate that Smad1 is a regulatory TF involved in PMS-mediated TIMP expression modulation in VSMCs.
PMS attenuates PDGF-BB-induced activation of FAK-related and p38 signaling pathways in VSMCs
TIMPs are known to bind to the integrin-β1/FAK complex to regulate MMP expression rather than acting as endogenous inhibitors abrogating MMP activities. 22 We observed that the phosphorylated levels of FAK at both Y397 and Y925 sites significantly increased after PDGF-BB treatment, and this effect was successfully abrogated by PMS. In addition, the levels of FAK-related proteins such as p-Src, vinculin, talin, and paxillin were altered to a similar extent after PDGF-BB or PMS treatment (Fig. 5A). No changes were observed in the level of integrin-β1 after PDGF-BB or PMS treatment.
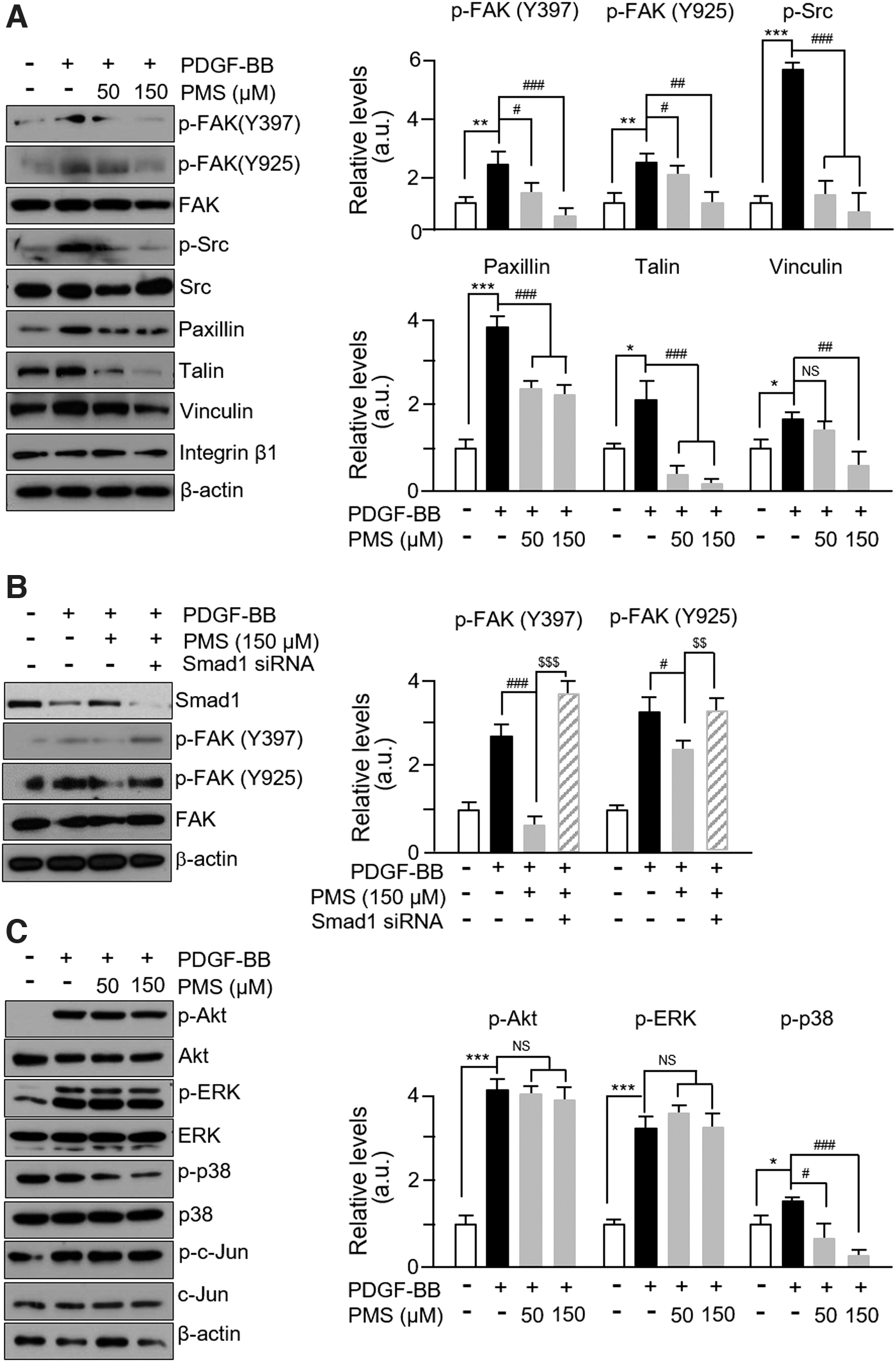
PMS attenuates FAK-related and p38 signaling pathways induced by PDGF-BB.
As expected, the phosphorylation of FAK at both Y397 and Y925 sites was not inhibited by PMS treatment in Smad1 siRNA-transfected VSMCs (Fig. 5B). Furthermore, the phosphorylated levels of p38 were significantly increased after PDGF-BB treatment, and PMS rescued this effect in a dose-dependent manner (Fig. 5C). However, the phosphorylated levels of the ERK and Akt were also significantly increased by PDGF-BB treatment and were unaffected by PMS (Fig. 5C). These results demonstrate that PMS-mediated altered TIMP expression is associated with the regulation of MMP expression through FAK-related and p38 mitogen-activated protein kinase (MAPK) pathways.
PMS inhibits neointimal formation in BI rat carotid arteries
We evaluated the effects of PMS administration on neointimal formation in BI rat carotid arteries. PMS was administered to rats with BI arteries via intraperitoneal injection (Fig. 6A). H&E staining revealed the bulky concentric neointimal formation after balloon injury but not in the control (uninjured) vessels. There was a significant reduction in the neointima area and the ratio of intima/media in PMS-treated BI rats (Fig. 6B). Subsequently, immunohistochemical staining showed that the number of cells stained positive for TIMP1 and TIMP2 decreased in BI vessels compared with that in the vessels of sham animals, and this effect was rescued in PMS-treated BI vessels. The levels of MMP2, MMP9, and MMP13 were reversed. Furthermore, the staining intensity for phosphorylated FAK at both Y397 and Y925 sites increased in BI vessels but decreased in PMS-treated vessels (Fig. 6C).
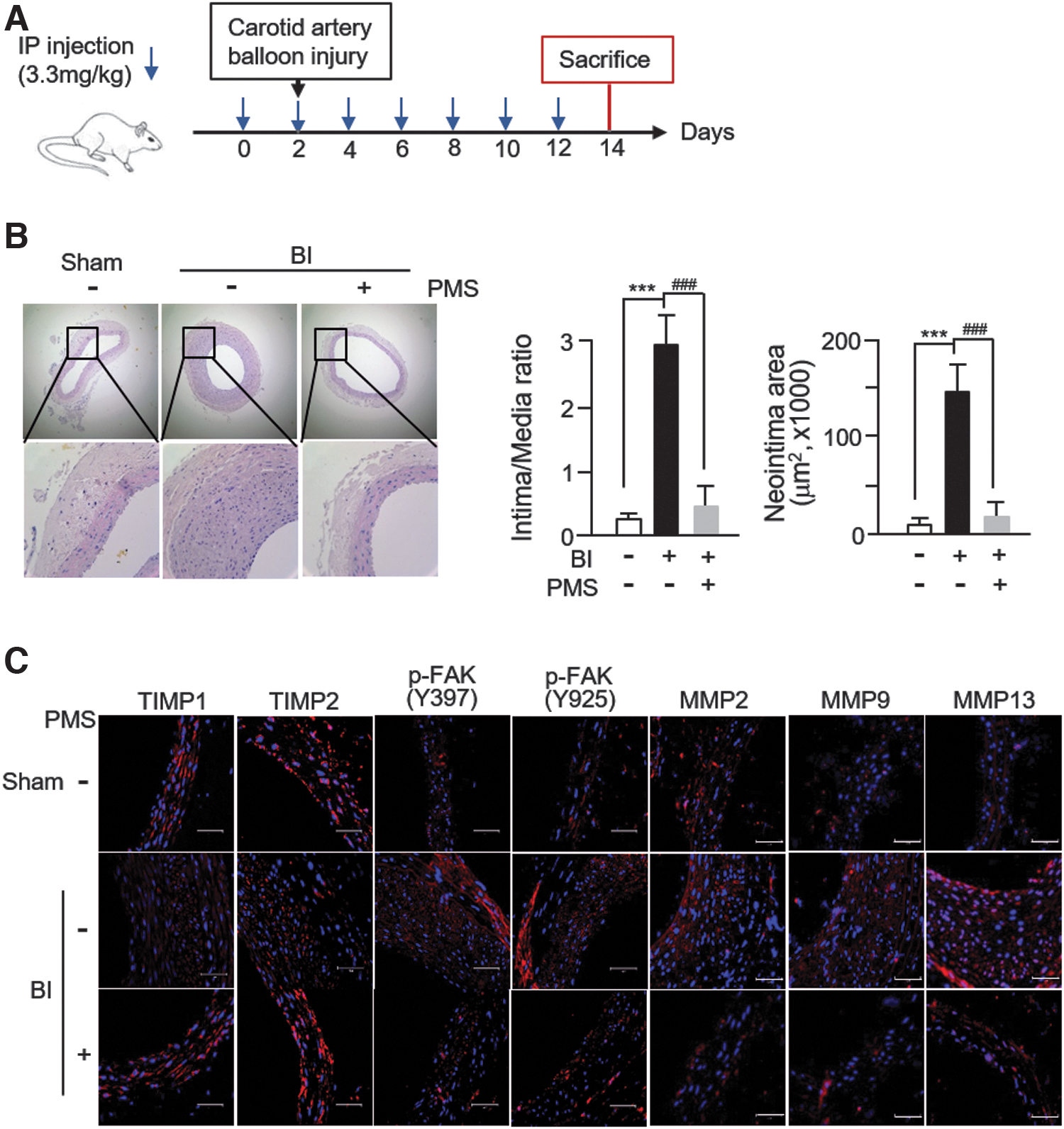
PMS inhibits neointima formation in BI rat carotid arteries.
These results indicate that the protective effects of PMS on neointima formation occurred specifically via the modulation of TIMP expression and the phosphorylation of FAK in VSMCs.
Discussion
Studies have suggested that PDGF-BB induces VSMC migration and proliferation, leading to neointimal hyperplasia in animal models with carotid injury. 7,8,23 Here, we demonstrate that PMS inhibited abnormal VSMC behaviors induced by PDGF-BB and provide potential therapeutic evidence to support its efficacy for preventing atherosclerosis and restenosis. The major findings of our study include: (i) PMS inhibits VSMC proliferation and migration through downregulation of cell cycle regulatory proteins and MMP2, MMP9, and MMP13, respectively; (ii) targeted signals of PMS for regulating proliferation and migration of VSMCs are FAK and p38; (iii) PMS activates Smad1 and subsequently upregulates TIMP expression, resulting in the inhibition of MMP expression and activities.
These findings are schematically illustrated in Figure 7. PMS specifically inhibits the phosphorylation of p38 MAPK among several signaling molecules activated by PDGF-BB, resulting in the suppression of CDK2 and MMP expression. In addition, the levels of other proteins that are well associated with cell proliferation and are known to be upregulated by PDGF-BB in VSMCs, 24,25 such as PCNA, CDK4, and cyclin D1, were also inhibited by PMS treatment. Furthermore, PMS activates Smad1, which further upregulates TIMP expression. TIMPs inhibit the conversion of pro-MMPs to active MMPs and bind to the integrin-β1 and FAK complex, resulting in the suppression of FAK-related VSMC proliferation and migration.
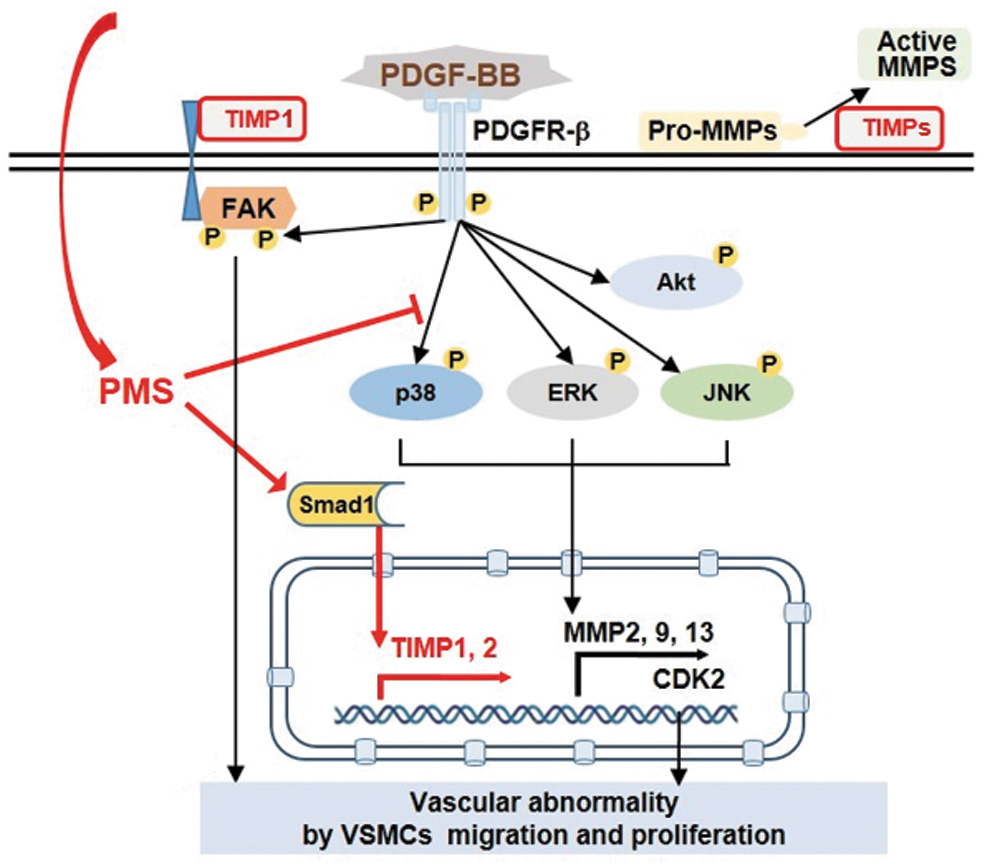
Proposed scheme for the inhibitory effects of PMS on the proliferation and migration of PDGF-BB-induced VSMCs.
Several chemicals inhibit PDGF-BB-induced behavioral changes of VSMCs through the regulation of signaling molecules such as MAPK family members, including p38, ERK1/2, c-Jun N-terminal kinase (JNK), and Akt. Ulinastatin, a urinary trypsin inhibitor, inhibited PDGF-BB-induced VSMC abnormal behaviors by downregulating Akt-related pathways, 20 whereas rubiarbonone C suppressed the activation of all three types of MAPKs. 16 Consistent with previous studies, our results show that PDGF-BB induced the activation of Akt, ERK1/2, p38, and JNK; however, PMS specifically decreased the phosphorylated level of p38, but not other activated signaling molecules (Fig. 5B).
In addition, PMS repressed the growth and metastasis of malignant melanoma cells through the inhibition of the phosphorylation of Akt, thus contradicting our result that PMS had no effect on phosphorylated Akt level induced by PDGF-BB in VSMCs. This discrepancy between previous reports and our results may be primarily owing to different cell types and experimental conditions. It would be interesting to investigate the specific effects of PMS on different cell types in future studies to expand its feasibility and applicability to various diseases.
We show, for the first time, that PMS significantly increased the expression levels of TIMP1 and TIMP2 through Smad1. In a previous study, ruxolitinib attenuated intimal hyperplasia by inhibiting the activation of signal transducer and activator of transcription 3 (STAT3), known as a proliferating TF, in VSMCs. 26 In addition, rubiarbonone C also inhibited STAT3 activation. 16 We observed the significant activation of STAT3 in PDGF-BB-induced VSMCs; however, PMS failed to alter STAT3 activation, instead significantly increased Smad1 activation (Table 2 and Fig. 4D). Our result that PMS did not increase the levels of TIMP1 and TIMP2 in Smad1-specific siRNA-transfected VSMCs demonstrates that Smad1 is a key TF regulating TIMP levels in PMS-treated VSMCs (Fig. 4E).
TIMPs are key regulators of ECM turnover, cellular behavior, and tissue remodeling and, thus, participate in various physiological and pathophysiological processes, such as angiogenesis, proliferation, migration, and apoptosis. 20 It is known that TIMPs inhibit the proteolytic activity of MMPs within the ECM. There are four kinds of TIMPs, namely TIMP1, TIMP2, TIMP3, and TIMP4; however, each TIMP inhibits different MMPs better than others. In addition, TIMP2 acts as an MMP activator. 27 Our results clearly show that the elevated levels of TIMP1 and TIMP2 after PMS treatment suppressed the activities of MMP2, MMP9, and MMP13 in VSMCs due to Smad1-mediated regulation.
This result is supported by a previous report that Smad factors participate in the regulation of TIMP1 expression by transforming growth factor-β1 and that TIMP1 expression is induced in Smad2, Smad3, or Smad4 knockout cell lines. 28 Although these authors did not provide any evidence regarding the underlying role of Smad1, our results provide a clear explanation (Fig. 4E). Furthermore, Smad1 is reported to be a key regulator promoting the contractile phenotype of VSMCs, 29 and PDGF-BB-induced phenotypic change of VSMCs was induced by Smad1 suppression, 30 which strongly supports our results.
Previous studies have shown that FAK participates in the process of cellular adhesion and movement and was deemed necessary for hepatic stellate cell (HSC) migration. 31 TIMP1 can attenuate cell migration by inhibiting the FAK activity. 32 Adiponectin reduced HSC migration by promoting TIMP1 secretion, which was mediated by TIMP1 binding to the integrin-β1/FAK complex, followed by inhibition of FAK phosphorylation. 22 Interestingly, we could not observe any alteration in the phosphorylation level of FAK until 12 h after PMS treatment (data not shown); however, we noted a significant decrease in the phosphorylation of FAK at both Y397 and Y925 sites (Fig. 5A). This decrease in the phosphorylated level of FAK was not observed in Smad1-specific siRNA-transfected VSMCs, suggesting that the elevated TIMP1 expression by Smad1 inhibits FAK phosphorylation.
Studies have suggested the therapeutic potential of PMS in various diseases. For instance, PMS attenuated isoproterenol-induced cardiac hypertrophy 11 and protected H9c2 cells against hypoxia/reoxygenation-induced injury. 12 In addition, PMS inhibited the growth and metastasis of tumor cells by decreasing the activity of MMP2 and MMP9. 13,14 Our results show that PMS significantly inhibited neointima formation induced by balloon injury. These effects were obviously associated with inhibitory effects of PMS on proliferation and migration, which were mediated by the suppression of p38 and activation of Smad1 followed by TIMP expression, respectively. In addition to the therapeutic potential, PMS or the extract of P. asiatica can be used as ingredients for food or health products. Indeed, no chronic toxicity of PMS concentrate was observed even at an intragastric dose of 2000 mg/kg. 33
This study has some limitations. One is that the long-term effects of PMS could not be evaluated in animal models. Second, the effects of various PMS concentrations have not been tested in animal models. Even though there are limitations for this study, our results provide strong evidence that PMS inhibits neointima formation in BI rat carotid arteries through downregulation of VSMC proliferation and migration. Therefore, PMS may serve as a potential therapeutic candidate for the treatment and prevention of atherosclerosis and restenosis. Further investigations on the signaling pathways involved in the antiproliferative and anti-migratory process of VSMCs are warranted to develop PMS-based treatment for vascular diseases.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by the Basic Science Research Program of the National Research Foundation of Korea (NRF), funded by the Ministry of Science, ICT and Future Planning, Grant/Award No.: 2021R1I1A3051564 and a grant from the Chosun University (2021).