
Abstract
Many studies have demonstrated that adipogenesis is associated with obesity, and the Hedgehog (Hh) signaling pathway regulates adipogenesis and obesity. Following the screening study of the chemical library evaluating the effect of vitexin on Gli1 transcriptional activity, vitexin was chosen as a candidate for antiadipogenic efficacy. Vitexin significantly reduced lipid accumulation and suppressed C/EBPα (CCAAT/enhancer-binding protein α) and PPARγ (peroxisome proliferator-activated receptor γ) expression, which are known as key adipogenic factors in the early stages of adipogenesis by activating Hh signaling. Furthermore, Hh inhibitor GANT61 reversed the effect of AMP-activated protein kinase (AMPK) activator AICAR (5-aminoimidazole-4-carboxamide ribonucleotide), indicating that Hh signaling is an upstream regulator of AMPK in 3T3-L1 cells. Vitexin suppressed adipogenesis by regulating Hh signaling and phosphorylation of AMPK, leading to the inhibition of fat formation. These results suggest that vitexin can be considered a potent dietary agent in alleviating lipid accumulation and obesity.
Introduction
Hedgehog (Hh)
Aberrant regulation of the Hh signaling pathway triggers developmental malformations and various cancers. 8,9 In addition, Hh signaling is closely associated with regulating adipogenesis and obesity. 10 It was reported that Hh signaling suppressed fly fat formation and the early stage of adipogenesis in mammals, 11 diverting preadipocytes and mesenchymal precursors toward osteogenesis. Also, the ectopic activation of Hh signaling by truncated Ptch1 decreased white adipose tissues during postnatal development 12 and knockdown of Hh-inhibiting transcription factor Gli3 reversed the early block in adipogenesis, 13 indicating the conserved role of Hh signaling in adipogenesis and obesity.
Obesity is caused by energy imbalance where energy intake is higher than expenditure and has a negative impact on human health. Obesity is one of the major health concerns worldwide. People with obesity are more likely to develop different chronic diseases including cardiovascular diseases, 14 type-2 diabetes, 15 and cancer. 16 Obesity is associated with hypertrophy (size increase of adipocytes) and hyperplasia (generating new adipocytes by preadipocyte differentiation) of the adipose tissue, which controls lipid storage and energy metabolism. 17 Adipocyte differentiation, having significant hypertrophic potential to increase adipocyte size, contributes to adipose tissue expansion. 18
Therefore, obesity can be prevented by blocking the differentiation process producing the mature adipocytes, accompanied by intracellular lipid accumulation, known as adipogenesis. 19 Adipogenesis is regulated by key adipogenic transcriptional factors, peroxisome proliferator-activated receptor γ (PPARγ) and CCAAT/enhancer-binding protein α (C/EBPα). 20
Considering the close association between obesity and Hh signaling, we previously screened various natural and dietary compounds to evaluate their regulatory effect on Hh signaling using the Shh-Light-II cell line having Gli-responsive element and luciferase reporter genes. Among them, vitexin, a bioactive flavonoid compound isolated from various dietary and medicinal plants such as mung bean and fenugreek, 21 shows significant dose-dependent stimulatory effects on the transcriptional activity of Gli1.
Many studies have reported the health-beneficial efficacies of vitexin on different chronic diseases, including cancers 22 and nervous system disease. 23,24 For example, vitexin exerts antitumor effects by suppressing the NF-κB pathway in nasopharyngeal carcinoma 22 and neuroprotective effect on pentylenetetrazole-induced seizure in male Wistar rats, as well as antidepressant-like effect through an increase of catecholamine level in BALB/c mice. 23,24 However, the role of vitexin on adipogenesis through the Hh signaling pathway has not been investigated.
In this study, the effect of vitexin on adipogenesis and the association of the Hh signaling pathway were investigated, and a novel underlying mechanism of this process in 3T3-L1 adipocytes was demonstrated. We also determined the intracellular communication between Hh signaling and AMP-activated protein kinase (AMPK) by using an Hh activator/inhibitor and AMPK activator during adipogenesis in 3T3-L1 adipocytes.
Materials and Methods
Materials
Insulin was provided from Gibco (Grand Island, NY, USA), and
Primary antibodies against Gli1, AMPKα, phosphorylated AMPKα (Thr172), and β-actin were from Cell Signaling Technology (Beverly, MA, USA). Those against PPARγ and C/EBPα were from Santa Cruz Biotechnology (Dallas, TX, USA), and the primary antibody against Smo was from Abcam (San Francisco, CA, USA). The secondary anti-mouse antibody was supplied by Cell Signaling Technology, and anti-rabbit secondary antibody was from Sigma-Aldrich.
Cell culture and adipocyte differentiation
The 3T3-L1 cell line was provided by American Type Culture Collection (Rockville, MD, USA) and was cultured in the media (Dulbecco's modified Eagle's medium [DMEM]) added with 10% (v/v) calf serum (Thermo Fisher Scientific, Wattham, MA, USA) and 1% penicillin/streptomycin (Gibco) in the incubator at the condition of 37°C and 5% CO2. Preadipocyte was differentiated by DMEM supplemented with MDI (insulin, 10 μg/mL; Dex, 1 μM; IBMX, 0.5 mM) treated with or without various concentrations of vitexin (1–20 μM). The medium was replaced by DMEM containing insulin (10 μg/mL) every 2 days during 10 days of incubation.
Cell viability assay
Cell viability was tested with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) reagent (Sigma-Aldrich). After seeding in 96-well plate (3000 cells/well) for 24 h, 3T3-L1 cells were treated with vitexin (1, 5, 10, and 20 μM) for 72 h before incubating with 20 μL MTT (5 mg/mL) for 4 h. Then, formed formazan crystal was dissolved by DMSO, and the final absorbance was detected by a microplate reader at 570 nm (Molecular Devices, Sunnyvale, CA, USA).
Oil Red O staining and BODIPY staining procedures
For Oil Red O staining, 3T3-L1 cells were fixed with 10% formalin, followed by washing with 60% isopropanol and staining with Oil Red O solution (Sigma-Aldrich) at room temperature (RT). Furthermore, stained cells were taken using a microscope (Olympus, Tokyo, Japan). Finally, the lipid droplets were dissolved with 100% isopropanol, and the absorbance was measured at 518 nm by a microplate reader (Molecular Devices).
For fluorescent staining, the cells were washed in phosphate-buffered saline and stained with boron-dipyrromethene (BODIPY) staining solution (500 μM) for 30 min at RT. Next, they were dyed by 4′,6-diamidino-2-phenylindole (DAPI) after 10% formalin fixation, and the picture were taken under a fluorescence microscope (Olympus). The fluorescent density of BODIPY and DAPI was evaluated using ImageJ software (Version 1.53c; National Institutes of Health, MD, USA).
Quantitative reverse transcription polymerase chain reaction assay
Total RNA was separated with TRIzol (Invitrogen, Carlsbad, CA, USA), and cDNA was made from RNA (1 μg) using PrimeScript RT Reagent Kit (Thermo Fisher Scientific). Primers (Table 1) and cDNA were applied for further quantitative RT-PCR (Q-RT-PCR). The target gene expression levels were normalized to the level of the glyceraldehyde 3-phosphate dehydrogenase, and relative quantification was performed using the comparative 2−ΔΔCT method. Forty cycles included 15-sec denaturation at 95°C, 60-sec annealing at 55°C, and 50-sec elongation at 72°C.
Primer Sequences for Quantitative Reverse Transcritption Polymerase Chain Reaction
C/EBPα, CCAAT/enhancer-binding protein α; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; PPARγ, peroxisome proliferator-activated receptor γ; Smo, smoothened.
Western blotting
After 24 h of treatment, cells pellet was collected in a 1.5 mL tube by scrapers and lysed in radioimmunoprecipitation assay buffer along with a cocktail protease inhibitor mixture for 30 min. Then, cell supernatants were saved after 15 min of centrifugation at 16,100 × g at 4°C. Protein quantification was performed by the bicinchoninic acid method. 25 Proteins were loaded in SDS-PAGE (sodium dodecyl sulfate–polyacrylamide gel electrophoresis) and further transferred to the polyvinylidene fluoride membrane (Millipore, Billerica, MA, USA). The primary antibody, that is, anti-Gli1 (1:1000), anti-PPARγ (1:1000), and anti-β-actin (1:2000), was incubated overnight. Then, they were further incubated with suitable secondary antibody. The immunoreactive signals were measured by a chemiluminescence imaging system (EZ Capture MG; ATTO, Tokyo, Japan).
Statistical analysis
The data are presented as means ± standard deviations. Group differences were conducted by one-way ANOVA (analysis of variance) and significant difference was determined at P < .05.
Results
Hh signaling activation by vitexin in MDI-induced 3T3-L1 adipocytes
We first screened the chemical library to confirm the effect of vitexin on the activation of Hh signaling in Shh-Light II cells and selected vitexin as a candidate molecule for Hh activation and adipogenesis inhibition. We showed that vitexin treatment at 0.2, 2.0, and 20 μM increased the Gli1 transcriptional activity by 1.3-, 2.5-, and 3.3-fold relatively compared to the control, respectively (Supplementary Fig. S1). The cell viability was determined by the MTT assay to assess the cytotoxicity of vitexin on 3T3-L1 adipocytes. As shown in Figure 1A, vitexin showed no significant inhibition in the cell viability of 3T3-L1 cells up to 20 μM.
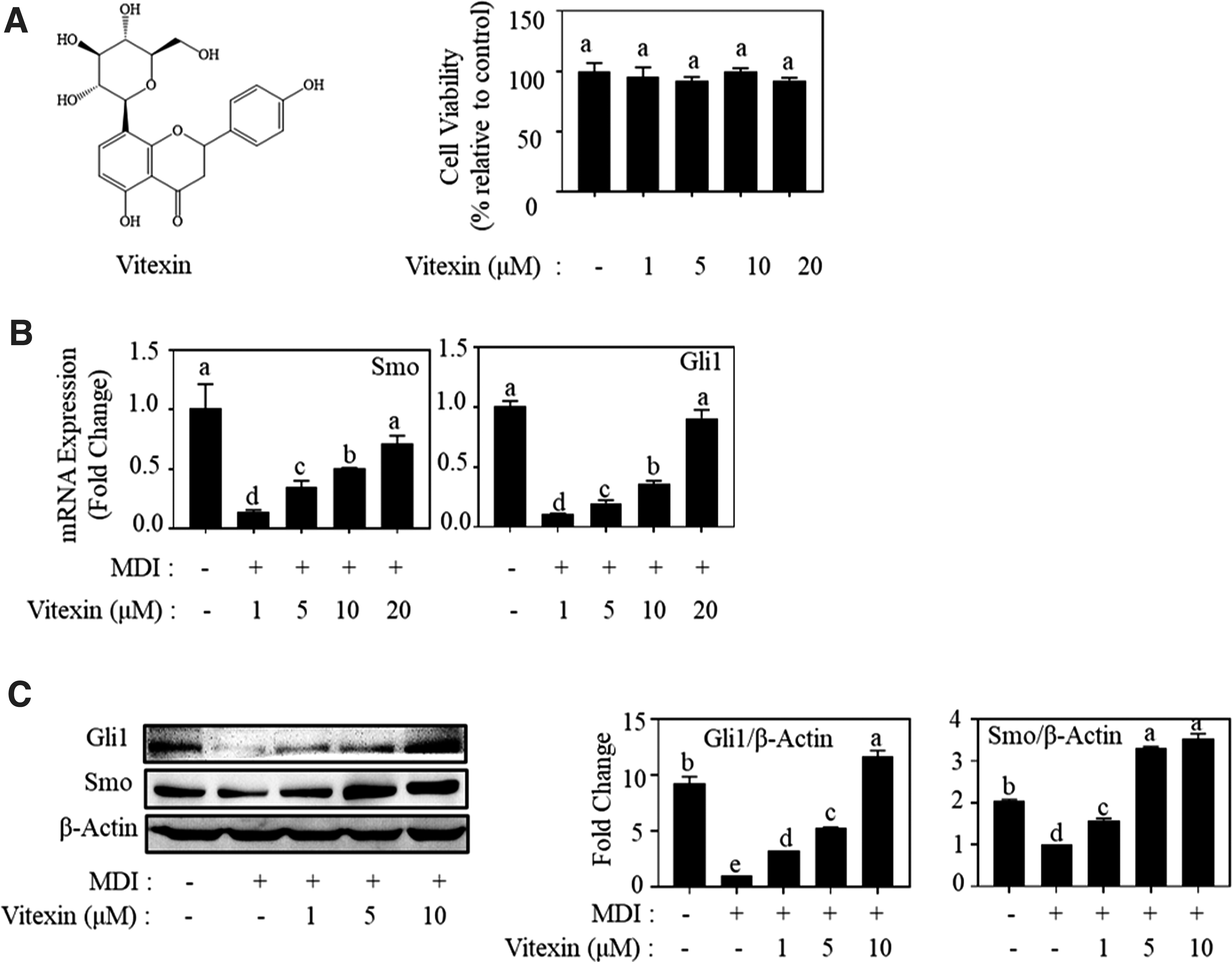
The stimulatory effect of vitexin on hedgehog signaling.
We found that vitexin enhanced the mRNA expression of Smo and Gli1 in MDI-induced 3T3-L1 adipocytes dose dependently (Fig. 1B). Vitexin treatment at 10 μM also increased the protein expression of Gli1 and Smo by 11.7- and 3.4-fold, respectively, compared to that of MDI-treated cells (Fig. 1C). These results indicate that vitexin activates Hh signaling in MDI-induced adipocytes.
Inhibitory effect on fat accumulation by vitexin in differentiated 3T3-L1 adipocytes
The stimulatory effect of vitexin on Hh signaling is shown in Figure 1. The effect of vitexin on fat accumulation was also investigated. Vitexin decreased the number of lipid droplets by 36.2%, 49.9%, 65.3%, and 77.1% at 1, 5, 10, and 20 μM, respectively (Fig. 2A). In the BODIPY staining experiment, the number of lipid droplets was represented by comparative green fluorescent intensity based on the cell number assessed by DAPI staining. The relative fluorescence intensities dropped to 85.1%, 71.5%, and 64.5% after treatment with vitexin at 1, 5, and 10 μM, respectively (Fig. 2B).

Inhibitory effect of vitexin on lipid accumulation and adipogenic markers in MDI-induced adipocytes.
Owing to their vital roles in regulating adipocyte differentiation, both C/EBPα and PPARγ expressions were analyzed in their mRNA and protein level. Vitexin at 1, 5, and 10 μM significantly reduced C/EBPα and PPARγ in both mRNA and protein levels dose-dependently (Fig. 2C, D) in MDI-induced 3T3-L1 cells. These results indicate that vitexin inhibits lipid accumulation through regulating the adipogenic mediators, C/EBPα and PPARγ.
Similar effect of vitexin and Hh activator on adipogenesis in MDI-induced 3T3-L1 adipocytes
To assess the effect of vitexin on lipid accumulation through the activation of Hh signaling, we compared the effect of vitexin (10 μM) and SAG (5 μM, a type of well-known Smo agonist 26 ) on adipogenesis. We discovered that 5 μM SAG suppressed lipid accumulation in 3T3-L1 adipocytes to 33.4% of the control group (Fig. 3A). SAG also enhanced the Smo and Gli1 protein expression levels, while inhibiting PPARγ and C/EBPα protein levels (Fig. 3B). Moreover, 10 μM vitexin demonstrated effects similar to those of SAG on the regulation of lipid accumulation and expression level of Smo, Gli1, C/EBPα, and PPARγ (Fig. 3A, B). These results reveal that vitexin inhibits adipogenesis by activating Hh signaling.
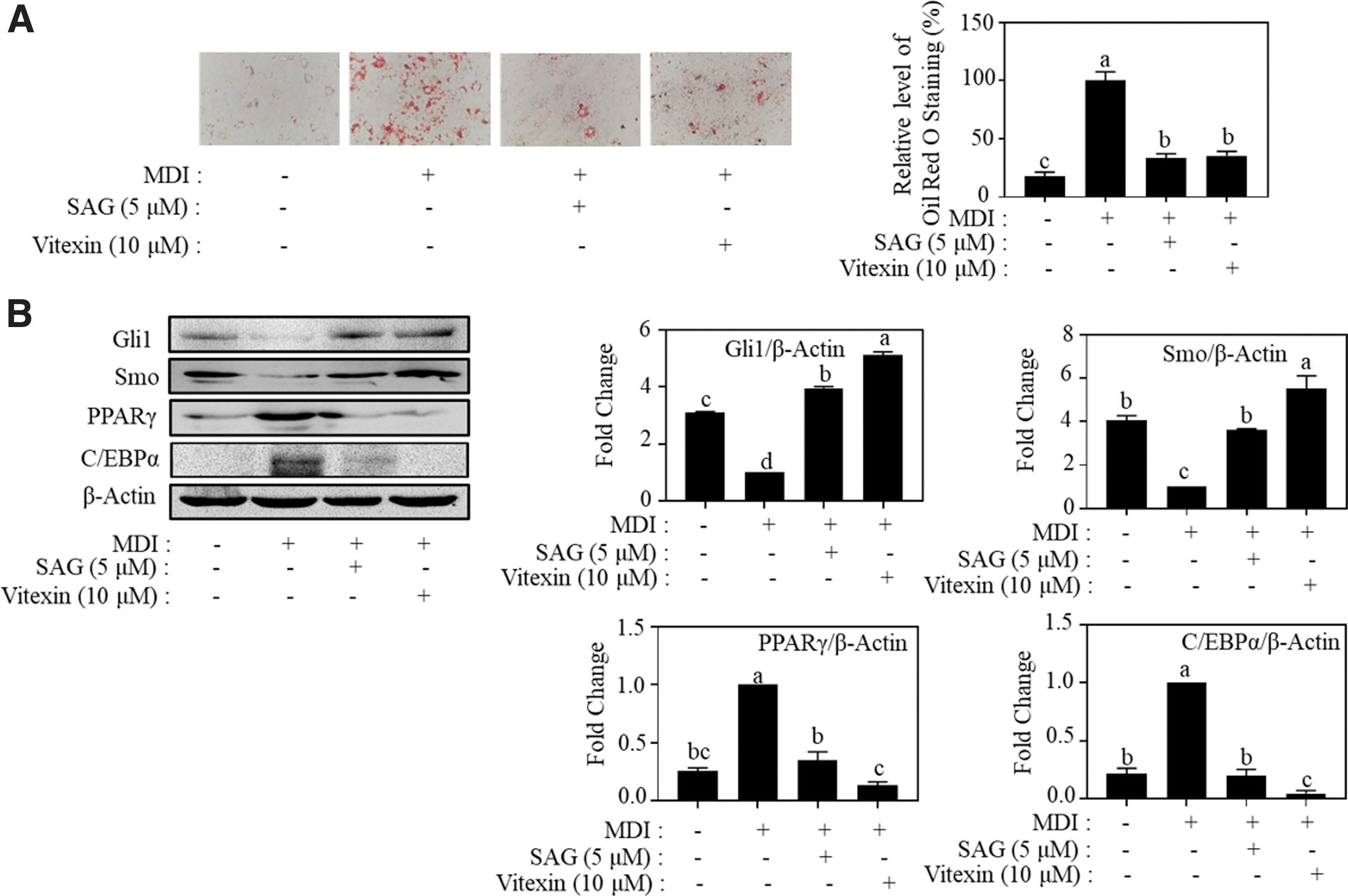
Effect of vitexin and SAG on adipogenesis in MDI-induced adipocytes. 3T3-L1 cells (1 × 105 cells/6 cm dish) were treated with vitexin (1, 5, and 10 μM) for 8 days.
Reversal of vitexin effect on adipogenesis by suppression of Hh signaling in 3T3-L1 adipocytes
To further confirm that vitexin inhibits adipogenesis through the Hh signaling pathway, we introduced GANT61 (Gli1 inhibitor) 27 and vismodegib (Smo inhibitor) 28 with vitexin to treat MDI-induced adipocytes. Treatment with GANT61 and vismodegib slightly promoted fat accumulation in 3T3-L1 adipocytes relative to that of the MDI-treated group (Fig. 4A, C). In addition, both Hh inhibitors reversed the suppressive effect of vitexin on lipid accumulation. Consistently, the activation of Gli1 and Smo and the inhibition of PPARγ and C/EBPα by vitexin were reversed by GANT61 or vismodegib (Fig. 4B, D). These results demonstrate that vitexin suppresses adipogenesis by Hh signaling activation.

Inhibitory effect of vitexin on adipogenesis was reversed by suppression of Hh signaling.
Inhibitory effects of vitexin on fat droplet formation at early stages of differentiation of adipocytes
3T3-L1 adipocytes undergo differentiation in the presence of adipogenic cocktail, following three different stages, early stage, postmitotic intermediate stage, and terminal stage, during days 0–2, 2–4, and 4–8, respectively. To determine the stages of adipocyte differentiation affected by vitexin, we used 10 μM vitexin at different stages of 3T3-L1 adipocyte differentiation (Fig. 5A). Treatment with vitexin at early stages (#III and #IV) significantly inhibited adipogenesis. However, vitexin showed a weak inhibitory effect when it was used after 4 days (Fig. 5B).
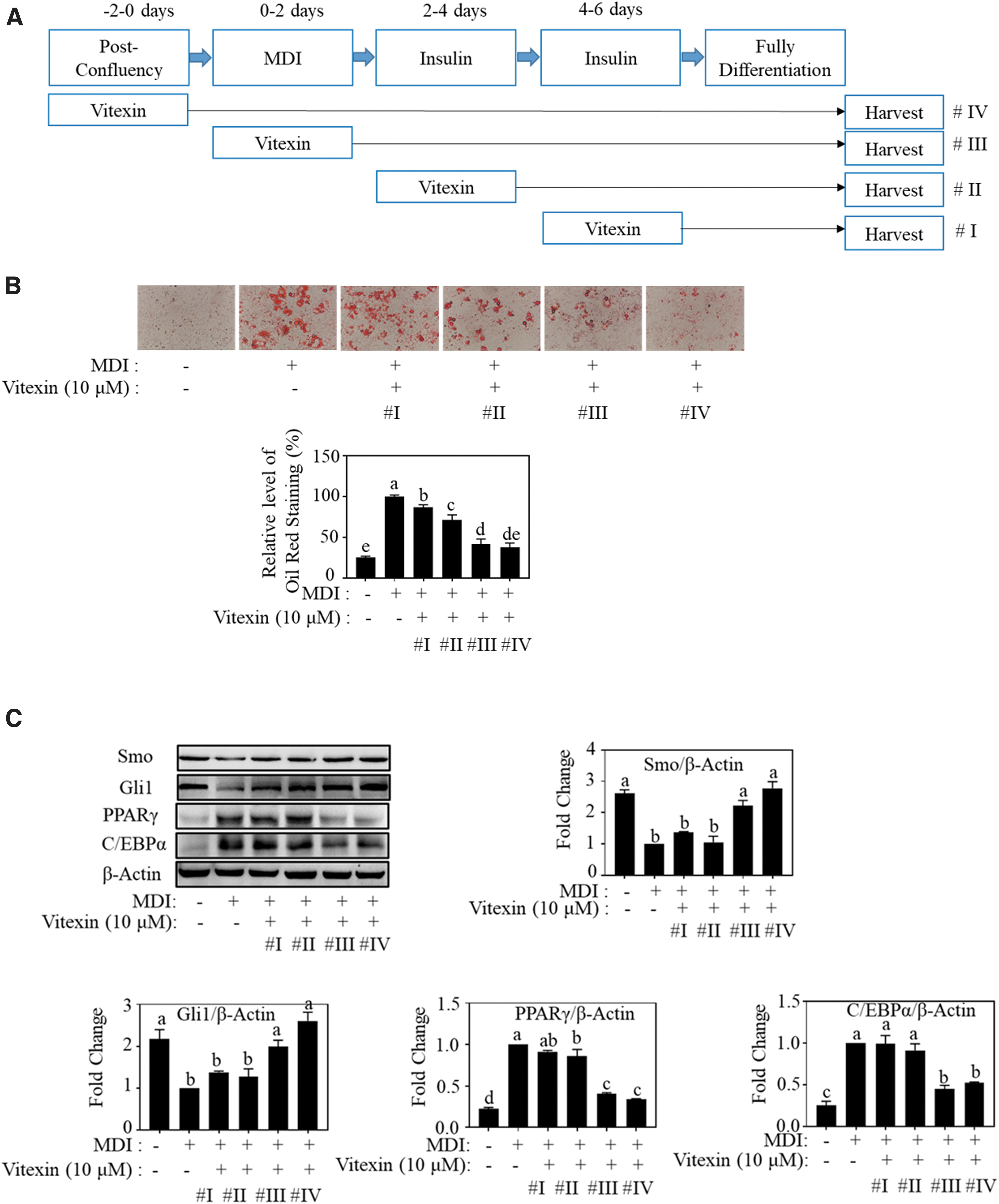
Vitexin exerts inhibitory effects on lipid accumulation at early stages of differentiation of adipocytes.
Consistent with these results, Gli1 and Smo protein levels were gradually enhanced by vitexin treatment from #I to #IV, and PPARγ and C/EBPα protein levels were gradually suppressed by vitexin during the differentiation process (Fig. 5C). These results indicate that vitexin exerts antiadipogenic effects during the early stages of 3T3-L1 adipocyte differentiation.
Blockage of AICAR effect on inhibition of fat accumulation by Hh signaling inactivation in adipocytes
Vitexin was reported to reduce obesity induced by high-fat diet (HFD) in C57BL/6J mice and suppress adipogenesis in 3T3-L1 cells by activating the AMPK signaling pathway. 29 We found that vitexin reduces adipogenesis in MDI-induced 3T3-L1 adipocytes by targeting Hh signaling. Therefore, we assume that intracellular cross-talk of Hh signaling and AMPK can affect adipogenesis. As shown in Figure 6, treatment with AICAR, a known AMPK activator, 30 decreased the number of lipid droplets by 43.5% relative to that of the positive control group. However, its effect was blocked by Hh inhibitor GANT61.
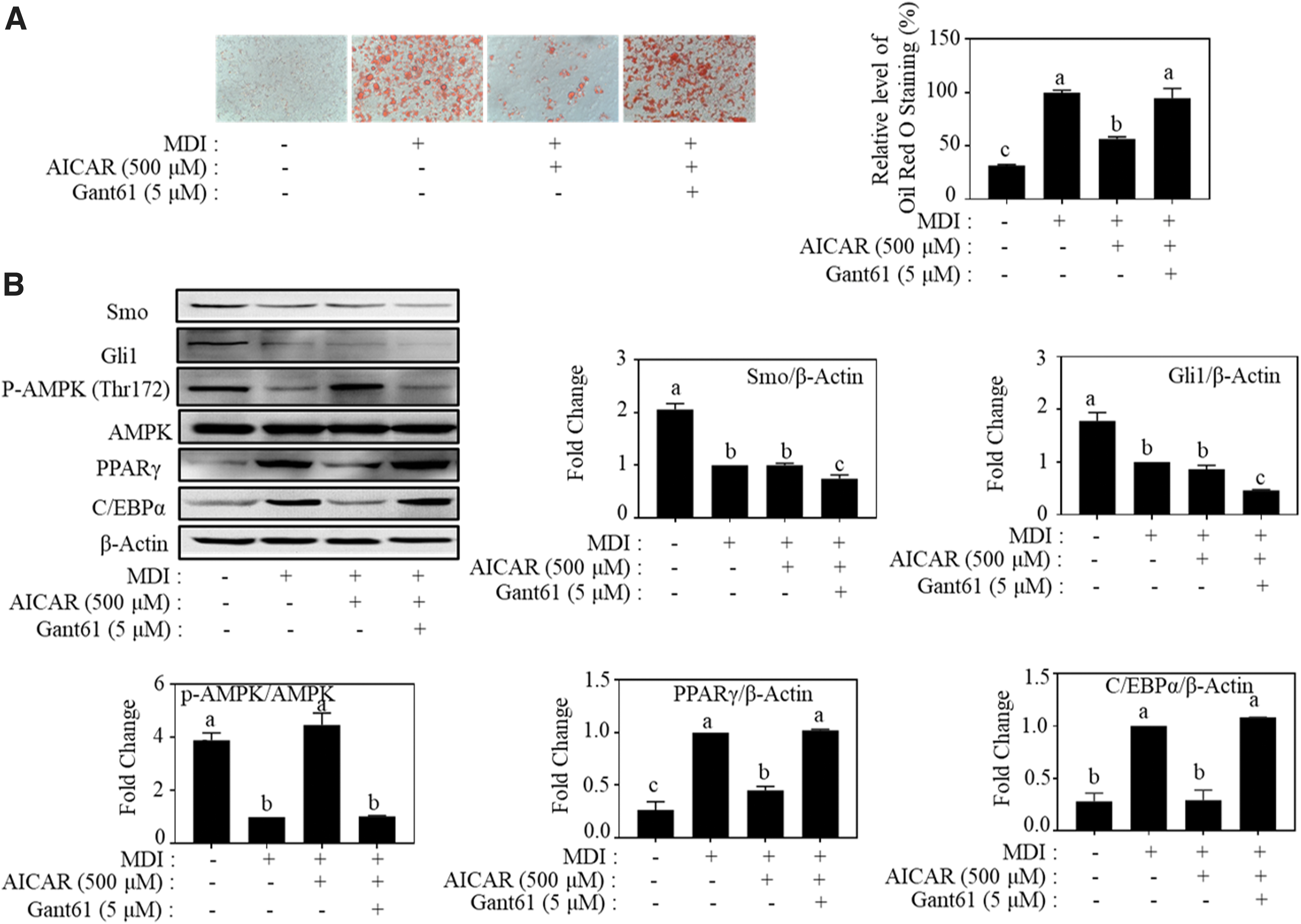
Inhibitory effect of AICAR on lipid accumulation was reversed by hedgehog inhibitor (GANT61) in MDI-induced adipocytes.
Besides, the upregulated expression level of p-AMPKα (Thr172) and downregulated expression level of PPARγ and C/EBPα by AICAR were reversed by GANT61. AICAR had no evident role in regulating Gli1 and Smo compared to the control. These results indicate that Hh signaling is an upstream regulator to AMPK and leads to the inhibition of adipogenesis.
Discussion
Adipocyte differentiation is a chronological process of growth arrest, mitotic clonal expansion, and early and terminal differentiation. 31 In preadipocytes, 3T3-L1, a widely used cell line for studying adipogenesis, differentiation can be initiated by treatment with appropriate inducers, including fetal bovine serum, glucocorticoids, cAMP agonist, and insulin. 32 After the growth arrest by full confluence of the cells, an adipogenic cocktail containing MDI regulates the expression of different transcription factors; C/EBPδ and C/EBPβ are first induced at the mitotic clonal expansion stage, peaked at the early differentiation stage, and reduced at the terminal differentiation stage. 33 The second wave of transcription factor PPARγ and C/EBPα expression follows; C/EBPα −/− fibroblasts decrease PPARγ expression and adipogenic capacity, and the restoration of PPARγ expression recovers the capacity. 34
These results suggest that PPARγ and C/EBPα are key adipogenic markers during adipocyte differentiation. We showed that MDI induced the expression of PPARγ and C/EBPα, as well as fat accumulation in 3T3-L1 adipocytes, which were significantly suppressed by vitexin dose dependently (Fig. 2).
To determine the adipocyte differentiation stages, which can be modulated by natural components, a recent study reported that sulforaphane, an isothiocyanate highly present in cruciferous vegetables, effectively suppressed the mitotic clonal expansion stage of differentiation by initiating the G0/G1 arrest in the cell cycle. 35 In addition, when quercetin, one of the most common flavonoids, was used as a treatment at physiologically achievable serum concentrations, it showed delipidating activity by acting at the early phase of adipogenesis, but it was not effective at lower concentration in mature adipocytes. 36 In a mechanistic study, the upstream signaling pathways such as Hh or Wnt/β-catenin signaling were regulated by a single molecule or extract, leading to the suppression of adipogenesis. 37,38
In this study, we divided the differentiation process into four levels to mimic the four stages of adipogenesis, and treated with vitexin at different time points. As shown in Figure 5, vitexin treatment at the growth arrest stage (#IV) and mitotic clonal expansion stage (#III) strongly inhibited fat accumulation, suppressed the protein levels of adipogenic markers, PPARγ and C/EBPα, and fully recovered Hh signaling mediators, Smo and Gli1. This result indicates that early exposure of vitexin is necessary for activating Hh signaling.
Mesenchymal stem cells (MSCs) in bone marrow are spindle shaped, and nonhematopoietic stem cells are known to be pluripotent cells that can differentiate into mature cells, including adipocytes and osteoblasts. 39 Hh signaling is suppressed during adipocyte differentiation of MSCs and plays a positive role during osteogenic differentiation, which indicates that Hh signaling is antiadipogenic and pro-osteogenic. 40 In addition, Hh signaling initiation by the fat-specific knockout of the Hh inhibitor, Sufu, in mice resulted in lean phenotypes, 41 and that by constitutively active Smo and Gli2 expression in the adipocyte lineage of the postnatal mice suppressed HFD-induced obesity, as well as the levels of adipogenic markers, PPARγ and C/EBPα. 42
Constitutive activation of Hh signaling did not alter glucose homeostasis, but improved insulin sensitivity. 41,42 We also showed that Hh activator SAG restored the expression of Smo and Gli1 and suppressed fat accumulation and the expression of PPARγ and C/EBPα at a similar level to that of vitexin treatment (Fig. 3). Also, blocking Hh signaling by using vismodegib and GANT61 targeting Smo and Gli1, respectively, significantly abolished vitexin's effect on fat accumulation and the expression of adipogenic markers (Fig. 4). These results strongly support that vitexin activates Hh signaling pathway to inhibit adipogenesis.
Vitexin has been identified as a major component of different plants such as Sasa coreana Nakai and Spirodela polyrhiza, and has been reported to exert regulatory effects on fat accumulation and protein levels such as PPARγ and C/EBPα in 3T3-L1 cells. 43,44 In the HFD-induced mouse model, the oral administration of vitexin (10 mg/kg BW) significantly reduced body weight (BW) and retroperitoneal fat. 45 However, these studies mostly showed the phenotypic effects of vitexin. Recently, Peng et al. 29 reported that vitexin alleviated obesity in HFD-fed male C57BL/6J mice through the regulation of the AMPK pathway.
Although a previous study showed that vitexin induces the phosphorylation of AMPK in vivo and in vitro, studies elucidating the underlying mechanism of this process are limited. In our study, we showed that the activation of AMPK activity by AICAR, which is known to phosphorylate AMPK at Thr172 by binding the γ-subunit of AMPK, 46 suppressed the expression of PPARγ and C/EBPα and fat accumulation in 3T3-L1 adipocytes (Fig. 6). However, AICAR did not affect the expression of Hh mediators, Smo and Gli1, and co-treatment with GANT61 counteracted the effect of AICAR (Fig. 6), indicating that Hh signaling is the upstream regulator of AMPK during 3T3-L1 adipocyte adipogenesis.
In conclusion, vitexin inhibits adipogenesis by downregulating PPARγ and C/EBPα and reduces fat accumulation in 3T3-L1 adipocytes. Vitexin activates the Hh signaling pathway by acting as an upstream mediator in the phosphorylation of AMPK. These results suggest that vitexin is a potent dietary agent that might be useful to prevent obesity if the results can be confirmed in human trials.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by Korea National Research Foundation (NRF-2021R1F1A1063279) and by the Bio-Synergy Research Project (NRF-2013M3A9C4078156) of the Ministry of Science, ICT, and Future Planning. Besides, this research was supported by the Academic scholarship for College of Biotechnology and Natural Resources provided by Chung-Ang University Graduate Research Scholarship in 2021.
Supplementary Material
Supplementary Figure S1