
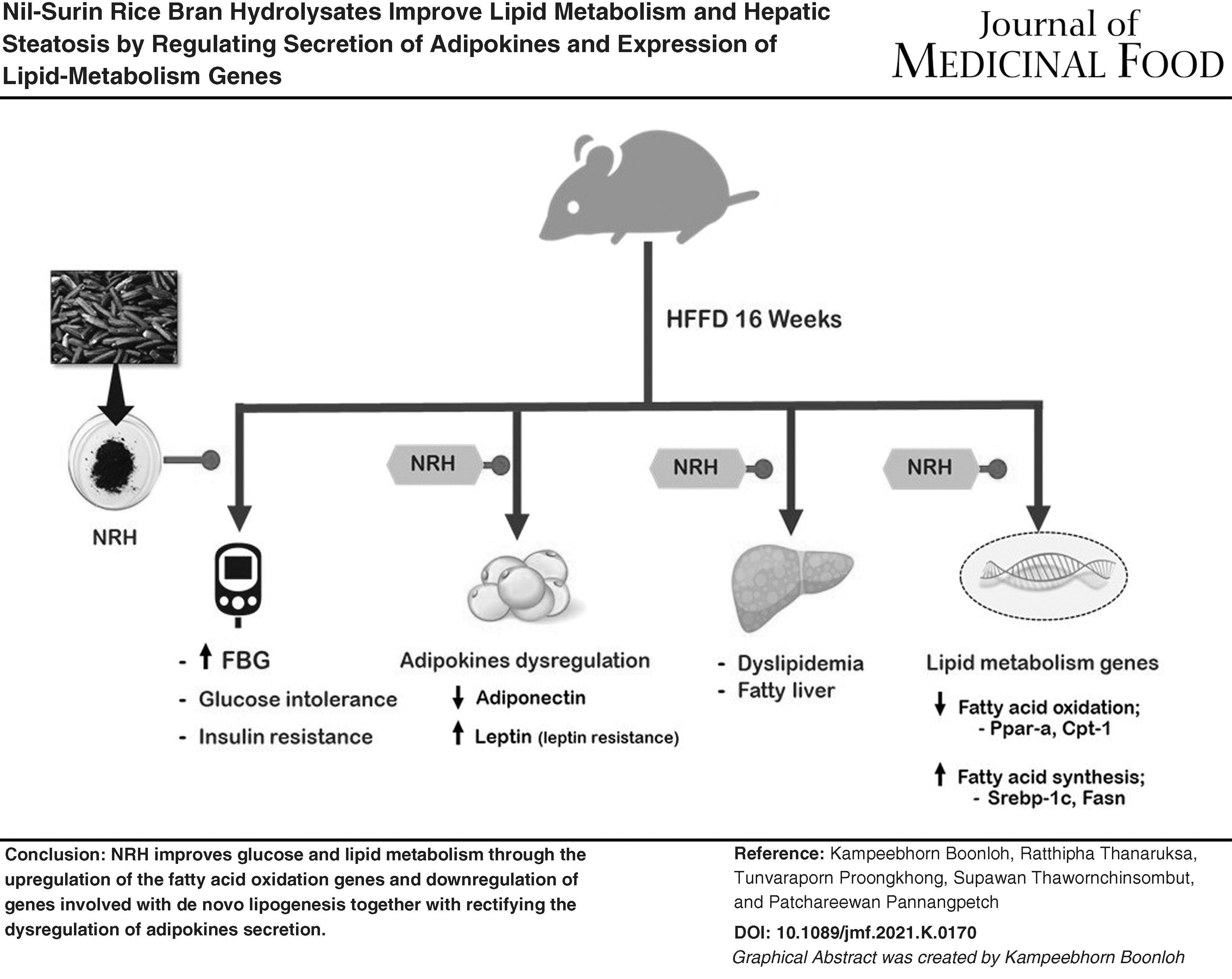
Abstract
Overconsumption of a high caloric diet is associated with metabolic disorders and a heightened risk of diabetes mellitus (DM), hepatic steatosis, and cardiovascular complications. The use of functional food has received much attention as a strategy in the prevention and treatment of metabolic disorders. This present study investigated whether Nil-Surin rice bran hydrolysates (NRH) could prevent or ameliorate the progression of metabolic disorders in rats in which insulin resistance (IR) was induced by a high fat-high fructose diet (HFFD). After 10 weeks of the HFFD, the rats showed elevated fasting blood glucose (FBG), impaired glucose tolerance, dysregulation of adipokine secretion, distorted lipid metabolism such as dyslipidemia, and increased intrahepatic fat accumulation. The IR was significantly attenuated by a daily dose of NRH (100 or 300 mg/kg/day). Doses of NRH rectified adipokine dysregulation by increasing serum adiponectin and improving hyperleptinemia. Interestingly, NRH decreased intrahepatic fat accumulation and improved dyslipidemia as shown by decreased levels of hepatic triglyceride (TG) and serum TG, total cholesterol and low-density lipoprotein cholesterol, and increased high-density lipoprotein cholesterol. In addition, a modulation of expression of lipid metabolism genes was observed: NRH prevented upregulation of the lipogenesis genes Srebf1 and Fasn. In addition, NRH enhanced the expression of fatty-acid oxidation genes, as evidenced by an increase of Ppara and Cpt1a when compared with the HFFD control group. The activities of NRH in the modulation of lipid metabolism and rectifying the dysregulation of adipokines may result in a decreased risk of DM and hepatic steatosis. Therefore, NRH may be beneficial in ameliorating metabolic disorders in the HFFD model.
Color images are available online.
INTRODUCTION
Insulin resistance (IR) plays an essential role in the development of metabolic syndromes (MS), dyslipidemia, type 2 diabetes mellitus (T2DM), non-alcoholic fatty liver disease (NAFLD), and cardiovascular complications. 1 –3 At the initial stage of IR, the body tries to maintain glucose homeostasis by requiring pancreatic β cells to produce more insulin, which leads to the condition of compensatory hyperinsulinemia.
However, this is still insufficient to control the blood glucose concentrations, which is indicated by an increase in homeostasis model assessment of insulin resistance (HOMA-IR) scores. 4 A chronic hypersecretion of insulin may cause β cell malfunction, which finally contributes to T2DM. 5,6
Chronic consumption of diets high in fats and carbohydrates can cause IR. 5,7 Carbohydrates, especially fructose, a monosaccharide mostly found in honey, soft drinks, and many fruits, as well as its derivative fructose-1-phosphate, are believed to be an important cause of lipogenesis and IR. 8 Fructose can produce metabolic disorders because it is metabolized in the liver, promoting the biogenesis of very low-density lipoprotein (VLDL) and de novo lipogenesis. 9
Over-consumption of highly saturated fats causes an increase in the amounts of lipid intermediates such as ceramides and diacylglycerol, which further activates proinflammatory signaling pathways, inducing oxidative stress and/or endoplasmic reticulum stress that contributes to systemic IR. 10 –12
It is well established that nutritional status can influence the regulation of sterol regulatory element-binding protein-1c (SREBP-1c); the key transcription factor encoded by Srebf1 is liable for the regulation of the genes involved with de novo lipogenesis. 13 In addition, as previously reported by our group, rats fed for long periods with a high fat-high fructose diet (HFFD) exhibited an increased expression of Srebf1 in the liver. 14,15
The upregulation of Srebf1 and its downstream targets gene such as fatty acid synthase (Fasn) leads to an expeditious stimulation of lipogenesis together with an increase of triglyceride (TG) concentration, hepatic IR, 16 and intrahepatic lipid accumulation (fatty liver), which could further develop into non-alcoholic steatohepatitis and other complications. 14,17
Peroxisome proliferator-activated receptor-α (PPAR-α), an essential transcription factor required for lipid and glucose homeostasis, is strongly expressed in the liver, skeletal muscle, and heart, which have high capacities for fatty acid oxidation. 18 It is notable that downregulation of the Ppara gene is associated with excessive fructose consumption. 19 Interestingly, PPAR-α agonists improve dyslipidemia and insulin sensitivity in both humans 20 and animal models. 21,22 Therefore, increased PPAR-α activity may be a potential therapeutic target for the management of MS.
Adiponectin is one of the influential adipokines on energy metabolism that is produced and secreted by adipocytes. It maintains energy homeostasis by the stimulation of fatty acid oxidation, promotion of glucose uptake, and inhibition of gluconeogenesis. 10,23 Moreover, adiponectin can produce anti-inflammatory effects by abolishment of the NF-κB signaling pathway. 24 Therefore, the level of adiponectin is conversely correlated to the inflammatory status of adipose tissues and IR condition. 25
Important strategies to alleviate MS consist of (a) lifestyle modifications, that is, caloric restriction, increased physical activity or (b) improving insulin sensitivity by using a medical approach, nutraceuticals, and eventually food supplements. Thus, it is important to identify new agents with insulin-sensitizing activity that might help to alleviate MS. Nil-Surin rice is a famous dark-purple local Thai rice cultivated mostly in Surin Province in Northeastern Thailand. Nil-Surin rice is rich in phenolic compounds, especially anthocyanins, vitamin E (α-tocopherol), and γ-oryzanol (
Nil-Surin rice bran is derived from the pericarp, which may contain numerous biologically active compounds. Interestingly, our previous studies revealed the antidyslipidemic, antidiabetic, and especially IR-improving activities of bran from white jasmine rice and light red or ruby-colored rice. 15,26,27 Therefore, bran of Nil-Surin rice may be a significant food supplement or nutraceutical for patients with MS or those at a high risk of T2DM. However, there is still a lack of evidence to support the therapeutic effects of Nil-Surin rice bran.
Thus, we explored the effect of Nil-Surin rice bran hydrolysates (NRH) on IR, glucose and lipid dysregulation, intrahepatic lipid accumulation, and adipokine secretion. For this, we used the HFFD-induced insulin-resistant animal model. The effects of NRH on the expression of genes regulating fat metabolism were especially investigated.
MATERIALS AND METHODS
Preparation of hydrolysates of Mali-Nil Surin rice bran
Nil-Surin rice, a dark-purple rice (Oryza sativa L. strain SRNC05053-6-2), is locally cultivated in northeast Thailand. Nil-Surin rice bran was kindly supplied by Surin Rice Research Center, Thailand. NRH was prepared according to a previous report. 28 Briefly, the protein fraction was extracted from defatted rice bran by alkaline solubilization (pH 9.5) for 1 h, and it was then heated at 127°C for 40 min, followed by pI precipitation (pH 4.5).
Proteolysis using a commercial enzyme (Protease G6; Genencor International Inc., Rochester, NY, USA) was performed at 0.5% (E/S) at pH 8.0, 55°C for 4 h, and then using Protease GN at 0.5% (E/S) at pH 8.0, 55°C for 2 h. The protease was then inactivated at 90°C for 15 min. After centrifugation, the protein hydrolysates were freeze-dried to obtain NRH powder. A yield of 23% NRH powder (based on weight of defatted rice bran) was achieved. The powdered NRH was stored at −20°C until it was used.
The NRH powder consists of ∼16% protein, 1.28% fat, and 65.98% carbohydrate and fiber. The anthocyanin content, measured as cyanidin-3-glucoside (C3G) described by Sutharut and Sudarat, 29 was 6.43 ± 0.24 mg/L extract.
An assay based on reverse-phase column and ultraviolet-vis diode-array detector high-performance liquid chromatography (HPLC-DAD) was used for the characterization and quantification of some phenolic compounds in NRH that was done by Central Laboratory (Thailand) Co., Ltd, Chiang Mai, Thailand. The amount of apigenin, catechin, gallic acid, isoquercetin, quercetin, tannic acid, and rutin were evaluated by using standard calibration curves of the pure chemicals.
Animals and experimental protocols
Male Sprague-Dawley rats (weighing 230–250 g) were purchased from Nomura Siam International Co., Ltd (Bangkok, Thailand). The rats were housed under 12 h light/dark cycles at 25 ± 1°C, fed ad libitum with a standard chow diet, and provided with sterile tap drinking water. All animal experimental protocols were conducted in accordance with the guidelines of the National Research Council of Thailand. Approval was received from the Animal Ethics Committee of Khon Kaen University (IACUC-KKU-8/62).
A total of 40 rats were subjected to the experiment. After a week of acclimatization, the normal control rats (n = 8; referred to as Group 1) were fed standard chow (Table 1) and given distilled water to drink until the end of the study. The remaining 32 rats were fed an HFFD (40% lard and 20% fructose) for 10 weeks to induce IR. At 10 weeks, the HFFD-fed rats were randomly allocated into four groups with eight rats per group and the HFFD diet was continued for a further 6 weeks; Group 2: HFFD rats given distilled water (as a negative control group); Groups 3 and 4: HFFD rats given NRH at 100 and 300 mg/kg/day; and Group 5: HFFD rats given pioglitazone, an insulin-sensitizing agent at 10 mg/kg/day (as a positive control treatment).
Composition of the Animal Food
The powdered rat food (CP Mice food, CP company, Samut Prakan Province, Thailand) is composed of 24% protein, 4.5% fat, 47% carbohydrate, 5% fiber, and 12% moisture supplemented with vitamins and minerals.
HFFD, high fat-high fructose diet.
The NRH and pioglitazone (Takeda Pharmaceutical, Osaka, Japan) were dissolved in distilled water. All treatments were administered by oral gavage. The FBG, oral glucose tolerance tests (OGTT), lipid profiles, insulin, leptin, and adiponectin levels were determined at the end of all treatments. After the rats were sacrificed by using the anesthesia drug, 5% isoflurane, the livers were collected for the determination of TG content, for the expression levels of lipid metabolism genes, and for the evaluation of histological changes.
Measurement of FBG, OGTT, IR, lipid profiles, and adipokine secretion
After completing the treatment, rats were fasted overnight (10–12 h) but allowed free access to water. Blood was collected from the lateral tail vein for the determination of FBG, which was done by using a Glucometer (Accu-Chek Performa, Roche Diagnostics, Mannheim, Germany). The rats were then orally given glucose solution at 2 g/kg body weight and blood-glucose levels were subsequently measured after glucose loading at 30, 90, and 120 min for the OGTT and to assess the state of IR. The area under the curve (AUC), from 0 to 120 min, was calculated by using the trapezoidal method.
Fasting serum was also collected for the determination of total cholesterol (TC), TG, low-density lipoprotein cholesterol (LDL-C), and high-density lipoprotein cholesterol (HDL-C) by enzymatic and colorimetric methods by using the Wako® diagnostics reagents (Wako Chemicals, Osaka, Japan).
Fasting serum insulin, adiponectin, and leptin levels were measured according to the manufacturer's protocols for the enzyme-linked immunosorbent assay (ELISA) kits for mouse/rat insulin, leptin, and adiponectin (ELISA Kit, Millipore, MA, USA). To evaluate the IR status, the HOMA-IR index was calculated as follows: HOMA-IR = Fasting glucose (mmol/L)/22.5 × Fasting insulin (μIU/mL). 30 Insulin 1 μIU/mL is equal to 6.945 pmol/L.
Determination of hepatic TG levels and histological changes
To determine whether NRH prevents hepatic lipid accumulation, hepatic TG levels were determined, and liver histological changes were assessed. The hepatic TG content was determined as previously reported. 31 Briefly, liver specimens were homogenized and dissolved in isopropanol. The liver homogenates were centrifuged, and supernatants were collected for the analysis of TG level by using a Wako Assay kit. The amount of TG was expressed as milligram per gram tissue.
To examine hepatic histological changes, the liver tissues were fixed in buffered neutral formalin (10%) and processed to be stained with hematoxylin and eosin for microscopic examination.
Determination of mRNA expression by reverse-transcription polymerase chain reaction
The liver tissues were stored at −80°C until subsequent RNA extraction. The expression levels of lipid-metabolism genes (Srebf1, FASN, PPAR-α, and CPT-1a) were evaluated. TRIzol® reagent obtained from Life Technologies (Carlsbad, CA, USA) was used for the RNA extraction according to the manufacturer's instructions. One microgram of total RNA was used for a synthesis of single-strand complementary DNA (cDNA) by using iScript™ reverse transcriptase (Bio-Rad, Hercules, CA, USA) at 25°C for 5 min, followed by 42°C for 30 min, with a final step of 5 min at 85°C in a C1000 Thermal cycler instrument obtained from Bio-Rad.
The real-time quantitative reverse-transcription polymerase chain reaction analysis was performed by Realtime QuantStudio™ 6 Flex instrument (Thermo Fisher Scientific, Waltham, MA, USA) using cDNA template, 0.5 μM of each primers, and 7.5 μL of SsoFast™ EvaGreen Supermix with low Rox (Bio-Rad) in a final reaction volume of 15 μL under the following conditions: initial denaturation at 95°C for 3 min followed by the amplification step with 40 cycles of 95°C for 15 s and 60°C for 31 s. The specific primers used are shown in Table 2. The expression of specific mRNA was calculated relative to the housekeeping β-actin (Actb).
Primer Sequences for Amplification of Lipid Metabolism Genes
bp, base pairs; PCR, polymerase chain reaction.
Statistical analysis
All values are presented as means ± standard errors of the means. One-way analysis of variance was used for the assessment of the differences between mean values of normally distributed data, followed by Tukey test for all pairwise multiple comparison to show specific group differences by using a computer-based software SigmaStat (Systat Software, San Jose, CA, USA). A P < .05 was considered statistically significant.
RESULTS
Phenolic content of NRH
High amounts of some phenolic compounds were found in NRH, including tannic acid, rutin, isoquercetin, and gallic acid. A moderate concentration of quercetin, catechin and apigenin were also found as shown in Table 3. Some phenolic compounds (i.e., kaempferol, eriodictyol, and hydroquinone) were under the limit of detection (<5 mg/kg).
Phenolic Compounds of Nil-Surin Rice Bran Hydrolysates
Effect of NRH on body weight
Feeding the rats with HFFD for the first 10 weeks did not cause any differences in body weight relative to controls. After continued feeding for a further 6 weeks, the rats fed the HFFD diet had higher body weight gains than did normal control rats. NRH 300 mg/kg was the most effective treatment in the prevention of body weight gaining of HFFD rats (Table 4). Pioglitazone group showed an increased body weight comparable with that of the HFFD control group.
Effect of Nil-Surin Rice Bran Hydrolysates on Body Weight and Percentage Increase in Body Weight in Each Experimental Group
All values are expressed as means ± SEM.
P < .05, significantly higher than that of normal controls.
P < .05, significantly lower than that of the HFFD control group.
BW, body weight; DW, distilled water; NRH, Nil-Surin rice bran hydrolysates; Pio, pioglitazone; SEM, standard error of the mean.
Effect of NRH on FBG and OGTT
Feeding the rats with HFFD for 10 weeks caused a notable increase in FBG (normal rats 79.33 ± 3.31; HFFD rats 87.25 ± 1.35 mg/dL), and impaired or defective glucose management as determined by OGTT and higher AUC of glucose (normal rats 14,931 ± 629; HFFD rats 15,837 ± 207). Subsequently, the HFFD rats were divided into four groups, as described earlier, and given daily administration of distilled water or NRH or pioglitazone for a further 6 weeks concurrent with the HFFD diet.
The NRH at both doses (100 and 300 mg/kg), or pioglitazone (10 mg/kg) remarkably diminished FBG level and AUC of HFFD rats as compared with that of HFFD control rats. This finding indicated that NRH and pioglitazone improved glucose tolerance in HFFD rats (Fig. 1).
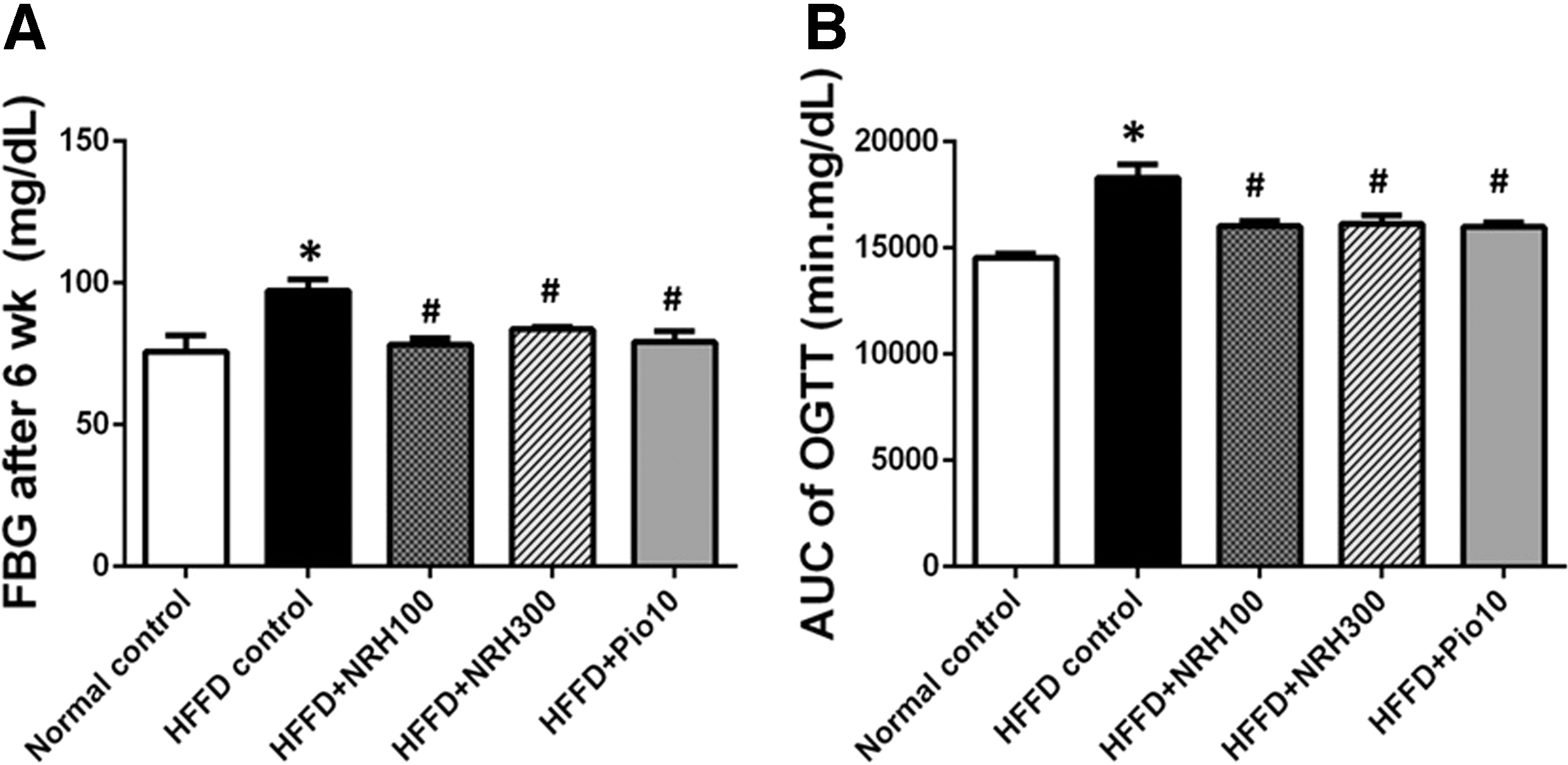
Effect of NRH on FBG
Effect of NRH on serum insulin and HOMA-IR scores
A marked increase in serum insulin and HOMA-IR scores in the HFFD control group was observed (Fig. 2A, B). The NRH (100 or 300 mg/kg) or pioglitazone 10 mg/kg significantly mitigated both serum insulin concentrations and the HOMA-IR score (Fig. 2A, B). This evidence suggested that NRH and pioglitazone could improve the body's glucose homeostasis.
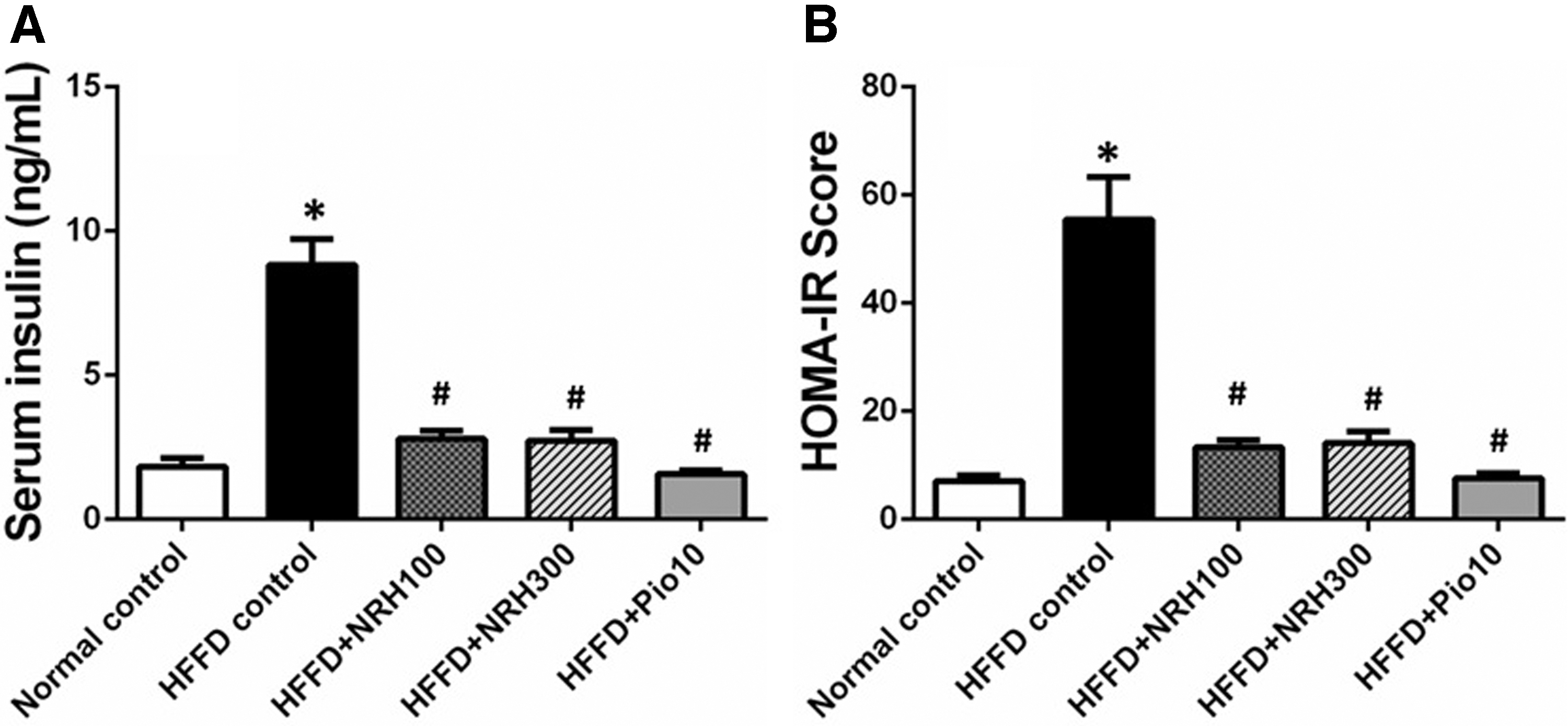
The effect of NRH on serum insulin level
Effect of NRH on adipokine secretion
To obtain more details about the effect of NRH on IR, we determined the levels of the two major adipokines; adiponectin and leptin, that function in lipid homeostasis and together with insulin in the regulation of glucose. The level of serum adiponectin in the HFFD control group was comparable to that of normal controls. However, NRH (100 or 300 mg/kg) or pioglitazone significantly raised the level of adiponectin in HFFD rats (Fig. 3A). The HFFD control group displayed a remarkable rise in serum leptin relative to normal controls, implying the state of leptin resistance in HFFD rats. Interestingly, NRH (100 or 300 mg/kg) or pioglitazone notably reduced the elevated serum leptin level of HFFD rats (Fig. 3B).
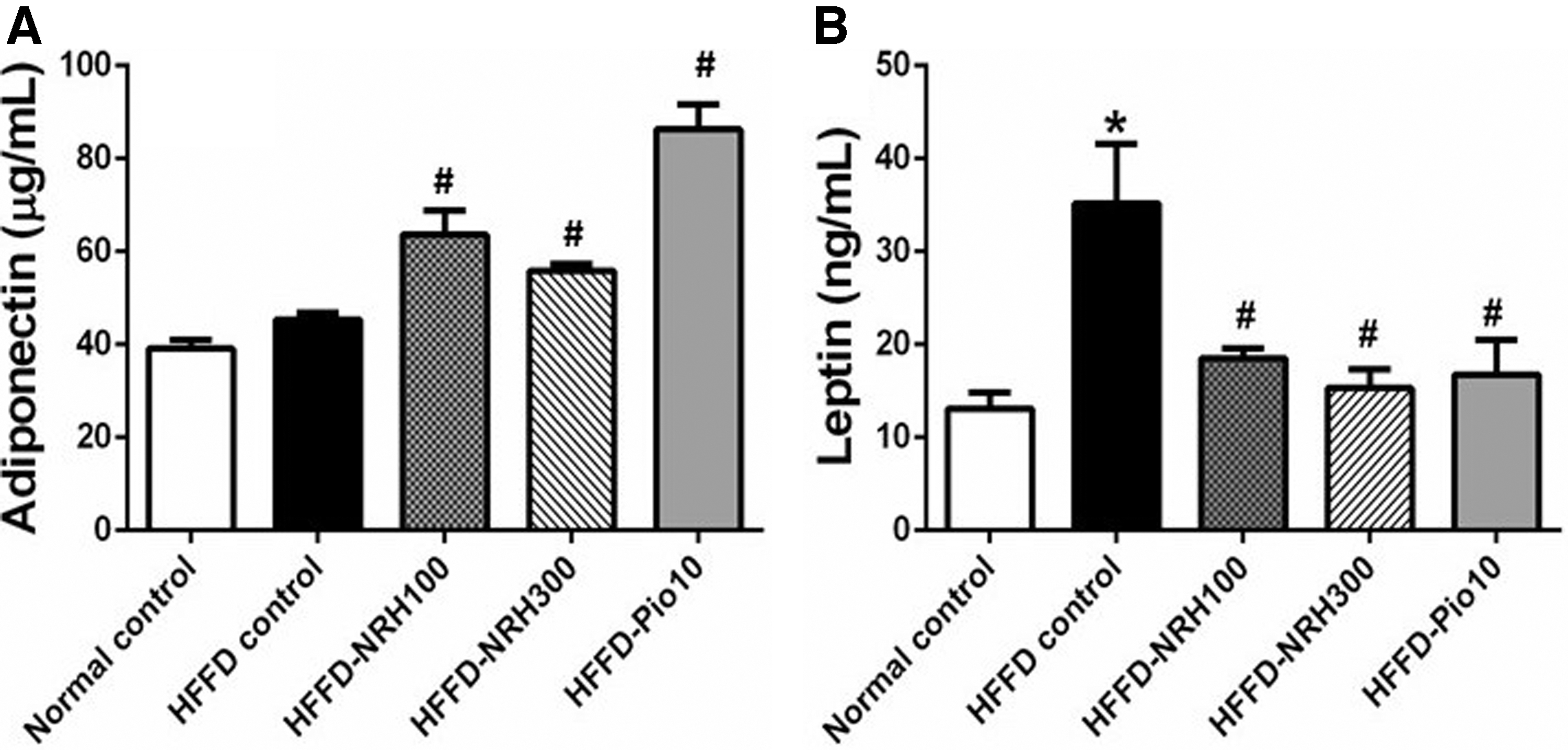
The effects of NRH on levels of serum adipokines; adiponectin
Effect of NRH on lipid profiles and hepatic TG concentrations
The HFFD control rats exhibited a dyslipidemic state; an increase in serum TG, TC, and LDL-C but a decline in HDL-C levels approximately by 25.13%, 45.55%, 89.83%, and 42.03%, respectively, as compared with normal control rats (Fig. 4A). The NRH (100 or 300 mg/kg) significantly reduced the elevated serum TG, TC, and LDL-C levels, but it increased the HDL-C level, indicating that NRH improved dyslipidemia. Pioglitazone decreased the levels of LDL-C and TG to an extent comparable to NRH but caused a substantial increase in HDL-C.
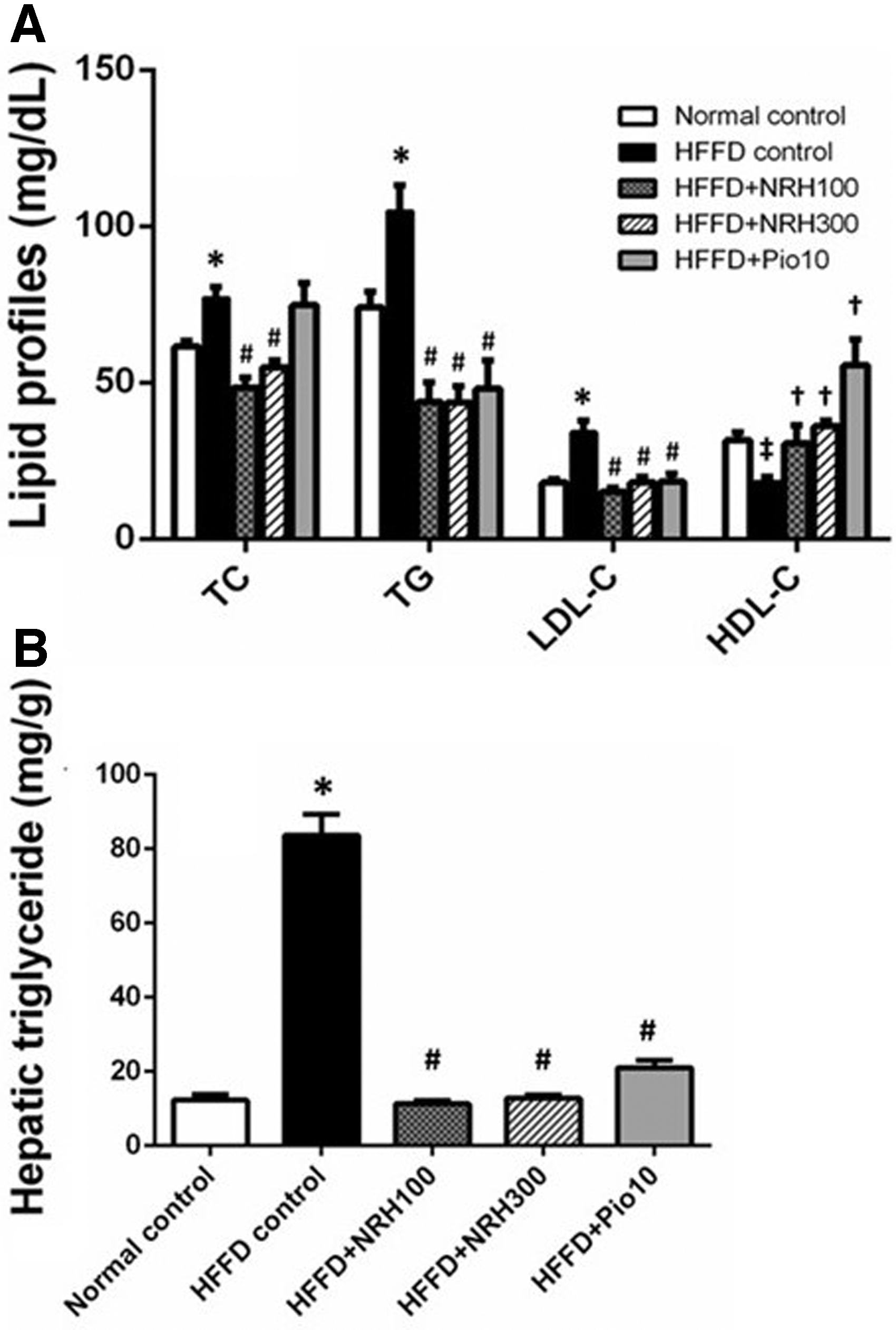
Effect of NRH on lipid profiles in HFFD rats
Analysis of hepatic TG contents demonstrated a high concentration of TG in the liver of the HFFD control group (Fig. 4B). The NRH and pioglitazone could substantially diminish TG accumulation in the liver of HFFD rats (Fig. 4B).
Effect of NRH on histology of the liver
Microscopic examination of liver tissue demonstrated that the numbers and sizes of lipid droplets were greatly increased in the HFFD controls relative to normal controls (Fig. 5A, B). The lipid droplets displaced the nuclei of hepatocytes. The NRH (100 or 300 mg/kg) and pioglitazone appeared to decrease both numbers and sizes of lipid droplets (Fig. 5B–E). This finding was consistent with the amount of TG found in the liver (Fig. 4B).
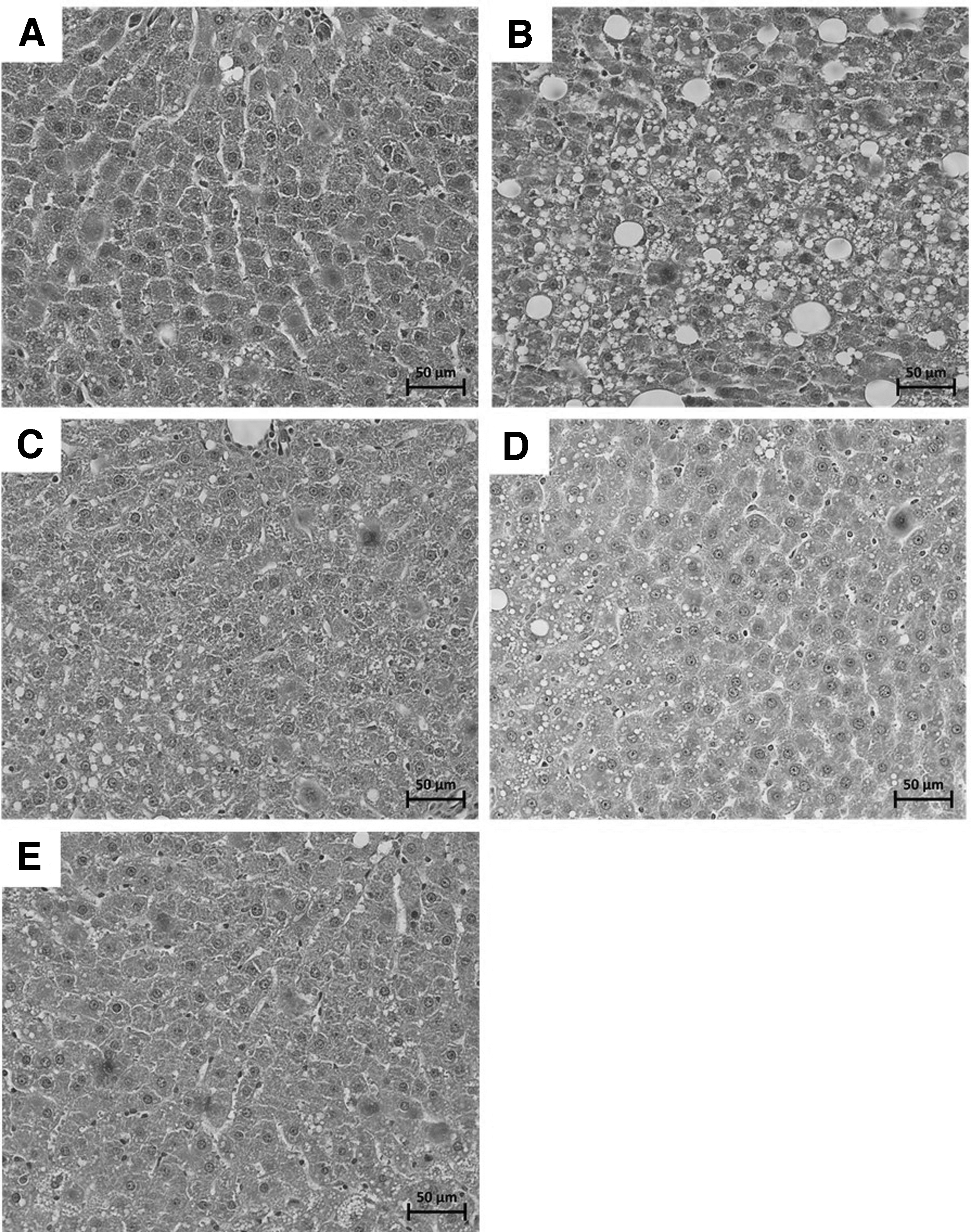
H&E staining for histological investigation of lipid accumulation in liver with original magnification of 400 × . Normal control group
Effect of NRH on the expression of lipid metabolism genes
To obtain more information about the mechanism of action of NRH involved in its lipid-lowering, and insulin sensitivity improving effects, we investigated the expression levels of genes involved with lipid metabolism; Srebf1 (a transcription factor involved with de novo lipogenesis) and its downstream signaling molecule, fatty acid synthase (FASN) in liver tissues. The HFFD rats had greater expression levels of Srebf1 (about 12-fold) than that of normal control rats (Fig. 6A).
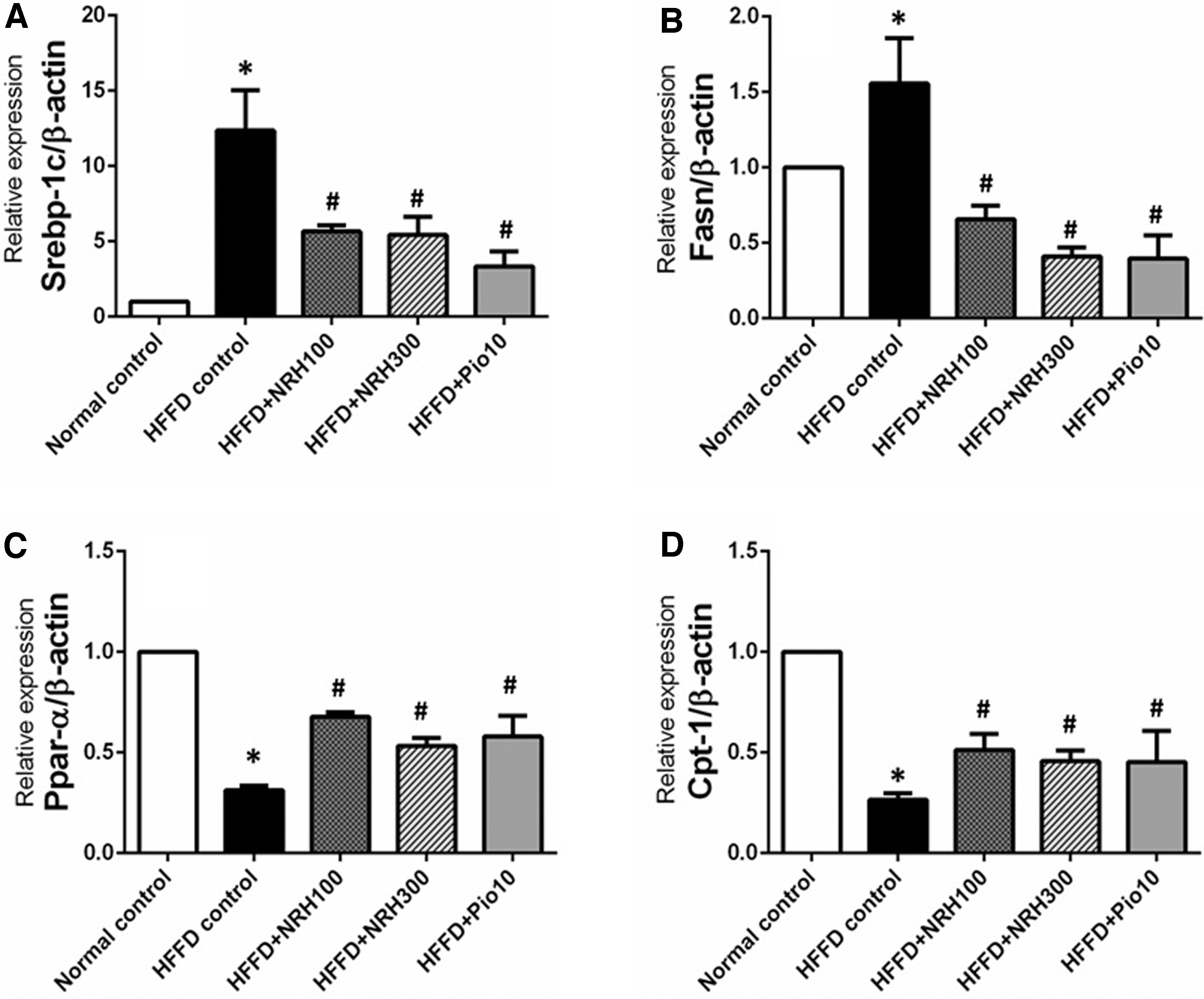
The effects of NRH on lipid metabolism genes in liver tissue; Srebp-1c
Interestingly, the elevated mRNA expression of Srebf1 was partially suppressed by the oral administration of NRH (100 or 300 mg/kg) or pioglitazone (by 54.2.2%, 56.1%, and 72.9% respectively). Consistent with this, the expression of its downstream gene, FASN, was also remarkably increased in HFFD rats, but this effect was reversed by treatment with NRH or pioglitazone (Fig. 6B).
The mRNA expression levels of the fatty-acid oxidation gene PPAR-α and its downstream target gene, carnitine palmitoyl transferase 1A (CPT-1a) in liver tissues were also analyzed. Relative to normal control rats, the HFFD control rats had lower expression levels of PPAR-α and CPT-1a by 68.7%, and 73.6%, respectively (Fig. 6C, D). The NRH (100 or 300 mg/kg) or pioglitazone produced a significant increase of PPAR-α expression in the HFFD rats by 54%, 41.3%, and 46%, respectively (Fig. 6C).
Similarly, the treatment with NRH at 100 or 300 mg/kg or pioglitazone significantly increased CPT-1a expression by 48.6%, 42.1%, and 42% respectively, compared with that of the HFFD control group (Fig. 6D). Taken together, this suggests that NRH or pioglitazone can modulate lipid metabolism by promoting fatty-acid oxidation and diminishing lipogenesis.
DISCUSSION
It is well established that excessive consumption of a high-calorie diet is associated with an increased body weight and contributed to hyperglycemia and hyperinsulinemia in both humans and animal models. 32,33 Our findings are consistent with the previously observed metabolic effects of high-energy diets. We demonstrated that over-consumption of an HFFD caused a series of metabolic changes as evidenced by significantly impaired glucose tolerance, increased blood glucose level together with compensatory hyperinsulinemia.
The results clearly suggest an insulin-resistant state, a crucial factor for MS that could further contribute to more progressive diseases such as T2DM, hepatic steatosis, and cardiovascular complications. The degree of metabolic disorder in HFFD control rats was quite high, as illustrated by a distinctly increased HOMA-IR score. The high insulin levels and HOMA-IR scores, together with the impaired OGTT, strongly suggested an insulin-resistant condition in HFFD-fed rats. We found that oral administration of NRH ameliorated IR in the HFFD rat model, as shown by decreased levels of FBG and serum insulin levels, improved glucose tolerance, and HOMA-IR score.
Chronic consumption of HFFD also causes a stress condition associated with dysregulation of adipokines, as indicated by an increase of serum leptin. Leptin is important in the modulation of insulin sensitivity via the suppression of gluconeogenesis and lipogenesis. 34 The increased serum leptin levels we observed after prolonged feeding with HFFD may indicate that the animals developed a compensatory hyperleptinemia or leptin resistance, a condition that has been linked to IR. 35,36 However, this situation was ameliorated by the administration of NRH or pioglitazone.
Adiponectin is another important adipokine that plays a crucial role in improving insulin sensitivity. 37 It regulates energy homeostasis by enhancing fatty acid oxidation and promoting glucose uptake but suppressing hepatic gluconeogenesis. In our study, the level of serum adiponectin in HFFD rats was similar to those of normal control rats. However, NRH increased serum adiponectin levels in the HFFD rats, which may facilitate an improvement of insulin sensitivity and help to rectify blood glucose levels.
Consistent with other reports, we demonstrated that inappropriate consumption of a high-energy diet can generate dyslipidemia as indicated by a marked increase of both serum and hepatic TG levels, as well as of serum TC and LDL-C, but a decrease in HDL-C levels. 38 –40 The IR provokes an imbalance between fatty acid oxidation and secretion as VLDL TGs and the influx of fatty acids, resulting in lipid deposition in the liver, and finally negotiates the hepatic steatosis (that so-called “nonalcoholic fatty liver disease”). In addition, free fatty acids can also induce oxidative stress and lipid peroxidation, which may facilitate hepatocellular inflammation and activate stellate cells, leading to liver fibrosis, cirrhosis, and may eventually result in the development of liver cancer. 41,42
Consistent with the biochemical finding of a high TG concentration, we microscopically observed an escalation in both size and number of lipid droplets accumulated in liver tissues in the HFFD control group. These changes were alleviated in HFFD rats given NRH or pioglitazone, as evidenced by significant decreases in hepatic TG concentrations and marked α decreases in lipid droplets in hepatocytes.
The impairment of hepatic lipid metabolism is closely associated with IR. Our study indicated that NRH could improve dyslipidemia and lessen hepatic lipid accumulation (Fig. 4). PPAR-α and its downstream target, CPT-1a, are essential for fatty acid homeostasis. Increases of PPAR-α and CPT-1 activity influence mitochondrial, peroxisomal, and microsomal oxidation, resulting in the enhancement of fatty acid catabolism and the prevention of hepatic lipid accumulation. 43 Interestingly, along with hepatic lipid metabolism, PPAR-α also regulates glucose homeostasis. 44
Activation of the PPAR-α receptor has been disclosed to improve inflammation, hepatic steatosis, and fibrosis in NAFLD in animal models, suggesting that PPAR-α is a potential target for both prophylaxis and therapeutic handling of MS and NAFLD. We demonstrated that prolonged feeding of rats with HFFD caused a decrease of both PPAR-α and CPT-1a mRNA levels. Interestingly, the administration of NRH could prevent the downregulation of those fatty acid oxidation genes.
Srebf1, a transcription factor gene and its downstream target, FASN, are responsible for lipogenesis. Srebf1 and FASN expression has been reported to increase in both humans and animals with excessive caloric intake. Animals fed a high-fat diet show an upregulation of Srebf1 and have elevated blood cholesterol and TG levels. 13 Here, we showed that Srebf1 and FASN were markedly upregulated after prolonged feeding with HFFD. Treatment with NRH could decrease the over-expression of these lipogenic genes. This is evidence that NRH can improve dyslipidemia by upregulating fatty acid oxidation and suppression of lipogenesis.
Taken together, our findings suggest that NRH improves glucose and lipid metabolism in HFFD-induced insulin resistant rats. NRH, anthocyanin-rich rice-bran hydrolysate, may rectify IR through modulating the dysregulation of adipokines. Remarkably, NRH promotes the upregulation of the fatty acid oxidation genes PPAR-α and CPT-1a, and the downregulation of genes involved with de novo lipogenesis (Srebf1 and FASN), resulting in a decrease in blood cholesterol, TGs, and intrahepatic lipid accumulation. Our findings provide evidence that NRH could be a nutraceutical for the management of diseases associated with IR, such as MS, and for the prevention of NAFLD.
Footnotes
ACKNOWLEDGMENT
The authors would like to acknowledge Prof. David Blair for editing the article through Publication Clinic KKU, Thailand.
AUTHORs' CONTRIBUTIONS
The authors declare that this work was done by the authors named in this article. K.B. and P.P. conceived and designed the experiments; K.B. performed the experiments; R.T. and T.P. provided Nil-Surin rice bran; S.T. prepared Nil-Surin rice bran hydrolysates; and K.B. and P.P. analyzed data and prepared the article.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This work was supported by the Rice Department, Ministry of Agriculture and Cooperatives of Thailand (Grant No. KKU 3/2562), and Cardiovascular Research Group, Khon Kaen University, Thailand.