
Abstract
Glioblastoma (GBM) is one of the most dangerous brain tumors in humans. The median survival of patients with GBM is <18 months. Glioma stem-like cells (GSCs), a small subpopulation of cells with stem cell-like characteristics found within GBM, are regarded as the main cause of GBM malignancy. Therefore, targeting GSCs presents an important therapeutic strategy for reducing the aggressiveness of tumors. In this study, we examined effects of (9Z,16S)-16-O-acetyl-9,17-octadecadiene-12,14-diynoic acid (AODA), a diacetylenic carboxylic acid isolated from leaves of Dendropanax morbiferus, on viability and self-renewal activity of GSCs. AODA substantially decreased GSC growth, causing apoptotic cell death as assessed by Annexin V/PI staining and morphological alterations by optical diffraction tomography. Interestingly, treatment with AODA suppressed ‘‘stem-like features’’ in vitro by limiting dilution assays and real-time polymerase chain reaction analysis. In addition, Western blotting revealed that AODA treatment decreased expression levels of phosphorylated AKT and phosphorylated ERK in GSC11 cells. Taken together, our results indicate that AODA could be considered a new therapeutic candidate to target GSCs.
INTRODUCTION
Glioblastoma (GBM) is one of the most malignant tumors, accounting for 12–15% of all brain tumors. 1 Despite conventional treatments of GBM such as surgeries, chemotherapy, and radiotherapy, the median survival time of patients is 12–18 months. 1,2 GBM is histologically characterized by high invasiveness in surrounding brain areas, pseudopalisading necrosis, and angiogenesis. 1 GBM has glioma stem-like cells (GSCs) as a specific cell population. GSCs have some capabilities of tumor initiation, resistance to anticancer therapies, and invasion. They could induce intratumoral heterogeneity. 3 –5 Therefore, targeting GSCs has been considered a major therapeutic strategy for GBM.
Recently, natural products have gained attention as potential sources of cancer chemotherapeutic agents. Since existing chemotherapy drugs target rapidly dividing cells and their nonspecific effects, some normal human cells are also affected, leading to undesired side effects. However, a rich source of bioactive natural compounds is relatively less toxic than existing anticancer drugs. In addition, their anticancer effects have been reported in many human malignant tumors, including breast cancer, liver cancer, and colorectal cancer. 6 –8
Dendropanax morbifera (Araliaceae) is a subtropical broad-leaved evergreen tree native to Korea. It can be found across the country's southern regions. This shrub's stems and roots are used in traditional medicine to cure a variety of ailments. 9 Extracts of D. morbifera have been recently demonstrated to possess several biological effects, including antiosteoclastogenic, antiamnesic, neuroprotective, antimicrobial, antidiabetic, antiplasmodial, and hepatoprotective activities. 9 –14 In addition, D. morbifera extracts can inhibit lung cancer cell migration and induce apoptosis or senescence in human hepatoma cell line. 15,16
Diacetylenes are chemical elements found in D. morbifera. They possess anti-inflammatory, anticomplement, anticancer, and antiosteoclastogenic properties. 10 –16 In this study, we isolated (9Z,16S)-16-O-acetyl-9,17-octadecadiene-12,14-diynoic acid (AODA), and a diacetylene carboxylic acid from the leaves of D. morbifera. However, few studies have evaluated anticancer effects of AODA in brain cancer. Thus, the objective of this study was to determine whether AODA isolated from D. morbifera could exert anticancer effects on brain cancer cells. We report that AODA can induce apoptotic cell death in GSCs and modulate extracellular-signal-regulated kinase (ERK) and protein kinase B (AKT) signaling.
MATERIALS AND METHODS
Materials
Leaves of D. morbiferus were harvested in May 2020 in Bosung county, South Korea. A voucher sample has been deposited in the Laboratory of Food and Natural Product Chemistry at Chonnam National University. These leaves were dried by hot air drying at 50°C for 48 h and ground to powder. Leaf powder was kept at −20°C until used.
Preparation of AODA from leaves of D. morbiferus
D. morbiferus leaf powder (1.5 g) was extracted with methanol (MeOH, 15 L) at room temperature for 24 h and filtered through No. 2 filter paper (Whatman, Maidstone, England). Residues were then extracted with 6 L MeOH and filtered by the same method mentioned above. MeOH extract filtrates were then combined and evaporated in vacuo at 40°C. After the MeOH extract (152.5 g) was suspended in distilled water (1 L) was partitioned with n-hexane (1 L) three times. The aqueous layer was partitioned successively with chloroform, ethyl acetate, and water-saturated n-butanol using the same method described above.
Ethyl acetate (EtOAc) layers were combined and concentrated under a vacuum at 38°C. The EtOAc layer (13.8 g) was purified by a medium-performance column chromatography system (MPLC; Isolera one, Biotage, Korea) equipped with a octadecylsilane (ODS) column (SNAP C18 120 g, 25 μm, Biotage). The mobile phase consisted of distilled water (solvent A) and acetonitrile (MeCN; solution B) using gradient elution (0 − 18 min 10% B; 18 − 130 min 10 − 100% B; 130 − 160 min 100% B) at a flow rate of 50 mL/min with UV detection at 210 nm and 254 nm. ODS-MPLC chromatogram revealed 29 compounds (D1−D29). Fraction D21 [retention time (tR ) 88–90 min, 345.6 mg] containing AODA was purified again with an ODS-MPLC system connected with SNAP C18 30 g (25 μm, Biotage).
The mobile phase consisted of 40% MeCN (solvent C) and 60% MeCN (solution D) using a gradient elution (0 − 6 min 100% C; 6 − 22 min 10 − 100% D; and 22 − 28 min 100% D) at a flow rate of 50 mL/min with UV detection at 210 and 254 nm. AODA (204.5 mg) was monitored at tR 21–22 min on the ODS-MPLC chromatogram. The molecular weight of AODA was determined using an electrospray ionization source-hybrid ion-trap time-of-flight mass spectrometer (Xevo G2-XS QTOF, Waters, Milford, MA, USA). The structure of AODA was also confirmed by 1D- ( 1 H and 13 C) and 2D- ( 1 H- 1 H correlation spectroscopy [COSY], heteronuclear single quantum coherence [HSQC], and heteronuclear multiple bond correlation [HMBC]) nuclear magnetic resonance (NMR) analyses using an unityINOVA 500 spectrometer (Varian, Walnut Creek, CA, USA).
Cell culture and reagents
Normal Human Astrocyte (NHA) were maintained in Astrocyte Medium (ScienCell Research Laboratories, CA, USA) supplemented with 10% fetal bovine serum (Gibco, Thermo Fisher Scientific, Waltham, MA, USA) and 1% penicillin/streptomycin (Welgene, Gyeongsangbuk-do, Korea). GSC11 and GSC23 cells were acquired from GBM patients at the University of Texas MD Anderson Cancer Center. 17 Neurobasal medium (NBE, serum-free neurobasal media supplemented with basic fibroblast growth factor [bFGF] and epidermal growth factor [EGF]) was used to maintain GSCs. NBE was prepared by adding 2% B27 (50) (Gibco, Grand Island, NY, USA), 1% penicillin/streptomycin (Welgene), EGF (20 ng/mL; R&D Systems, Minneapolis, MN, USA), and bFGF (20 ng/ml; R&D Systems) to Dulbecco's modified Eagle's medium/F12 (Welgene).
Cell viability
Cell viability was conducted with an alamarBlue® solution (Invitrogen). NHA and GSCs were seeded at a density of 3000 cells per well into 96-multiwell plates (n = 6). These cells were then treated with or without AODA at different concentrations. After incubation for 48 h, alamarBlue solution was added to each well followed by incubation for 6 h. The fluorescence of the solution was measured at wavelengths of 570 and 600 nm using a Synergy HTX Multi-Mode Reader (BioTek Instruments Inc., VT, USA).
Annexin-V and propidium iodide staining
NHA, GSC11, and GSC23 were treated with dimethyl sulfoxide or AODA (10 and 25 μM) for 24 h. These cells were then harvested, washed with cold phosphate-buffered saline (PBS), and incubated with Annexin-V-FITC and propidium iodide (Invitrogen) at room temperature for 15 min. Incubated cells were then analyzed using a flow cytometer (Beckman coulter).
In vitro limiting dilution assay
For a limiting dilution assay, cells that decreased twice per well (50, 25, 12, 6, 3, and 1) were seeded on a 96-multiwell plate. The tumorsphere formation in each well was observed. The calculation of stem cell frequency was conducted with a website available at
Quantitative reverse transcription-polymerase chain reaction
Total RNA was extracted with RiboEx reagent (GeneAll, Seoul, Korea) and purified with a Hybrid-R kit (GeneAll) as directed by the manufacturer. A Revert Aid First-Strand cDNA Synthesis kit mRNA (Thermo Fisher Scientific) was used to synthesize cDNA from 500 ng of mRNA. TB Green Premix Ex Taq I™ (Takara Bio, Shiga, Japan) was used in a real-time polymerase chain reaction (PCR) on a Qtower3 real-time PCR thermal cycler (analytikjena, Jena, Germany). Results of quantitative reverse transcription-PCR (qRT-PCR) were evaluated as Ct values and quantified using the 2-ΔΔCt method. Primer sequences used for RT-qPCR amplification were as follows (5’ to 3’): human 18S rRNA forward, 5′-CAGCCACCCGAGATTGAGCA-3′; human 18S rRNA reverse: 5′-TAGTAGCGACGGGCGGTGTG-3′; human CD133 forward, 5′-CAGGTAAGAACCCGGATCAA-3′; and human CD133 reverse:5′-TCAGATCTGTGAACGCCTTG-3′.
Neurosphere assay
GSCs treated with AODA as described above were seeded at a density of 1 × 104cells/well into 24-well plates and incubated for 7 days at and 37°C with 5% carbon dioxide without disturbing the plates or refilling the media. Using a digital microscope (Logos Biosystems, Anyang-si, Gyeonggi-do, Korea), photographs of neurospheres were obtained at the end of the 7-day incubation period to measure sizes of neurospheres.
Apoptosis imaging using three-dimensional optical diffraction tomography
Apoptotic morphologies of GSCs were monitored using three-dimensional (3D) ODT (Tomocube, Daejeon-si, Korea) according to the method presented by Park et al. 18 To observe live cells with ODT, GSCs (1 × 103 cells) were seeded into the central glass-bottom Tomodish. For apoptosis imaging, GSCs (GSC11 and GSC23) were treated with AODA (20 μM) for 24 h. They were then washed with PBS (pH 7.4) and imaged using 3D ODT.
Western blot analysis
GSCs treated with AODA were lysed with radio-immunoprecipitation assay buffer supplemented with phosphatase inhibitor cocktail 2 (ApexBio). A bicinchoninic acid Protein Assay Kit was used to determine protein content (Thermo Fisher Scientific). Proteins were separated on 10% and 15% sodium dodecyl sulfate–polyacrylamide gel and then transferred to polyvinylidene difluoride membranes. These membranes were blocked with 5% skim milk in phosphate buffered saline with Tween®20 (PBST) at room temperature for 1 hour and then incubated with corresponding primary antibodies at 4°C overnight with moderate shaking. After washing with PBST, membranes were incubated with an appropriate secondary antibody at room temperature for 1 hour. Membranes were then visualized using chemiluminescence (Invitrogen) following the manufacturer's protocol after they were washed with PBST.
Statistical analyses
GraphPad Prism 8 (GraphPad Inc.) and Microsoft Excel (Microsoft Inc.) were used for statistical analyses. Statistical significance between and among groups was determined by a two-tailed t-test and one-way analysis of variance, respectively, followed by Tukey's multiple comparison test.
RESULTS
AODA isolation from D. morbiferus
AODA (204.5 mg) was isolated from the leaf powder (1.5 g) of D. morbiferus. AODA high-resolution electrospray ionization-mass spectrometry (positive) data exhibited a sodiated molecular ion peak at m/z 353.1725 [M+Na]+ (calculated for C20H26O4Na, m/z 353.1729, −0.4 mDa), indicating the molecular formula (C20H25O4) and molecular weight (330 Da) of AODA. 1 H-NMR (500 MHz, CD3OD) spectra exhibited characteristic proton signals of one oxygenated methine at δ 4.08 (H-6) and five sp 2 methines at δ 6.21 (H-13a), 5.48 (H-13b), 4.95 (H-14a), and 4.86 (H-14b) (Table 1). 13 C-NMR (125 MHz, CD3OD) spectra revealed the presence of 20 carbon signals, including two carbonyl carbons at δ 177.74 and 171.11, four olefinic double-bond carbons at δ 134.14–119.74, four triplet bond carbons at δ 81.56–64.73, one oxygenated methine carbon at δ 65.82, and other methylene and methyl carbons at δ 35.06–18.24.
Signals of C-4∼C-7 are exchangeable.
These were confirmed by MS and HSQC experiments (Table 1). The planar structure of AODA was determined by 1 H- 1 H COSY (bold lines) and HMBC (arrows) experiments (Fig. 1A). Conclusively, AODA shown in Figure 1A was finally identified by comparing data with MS and NMR results previously reported. 19
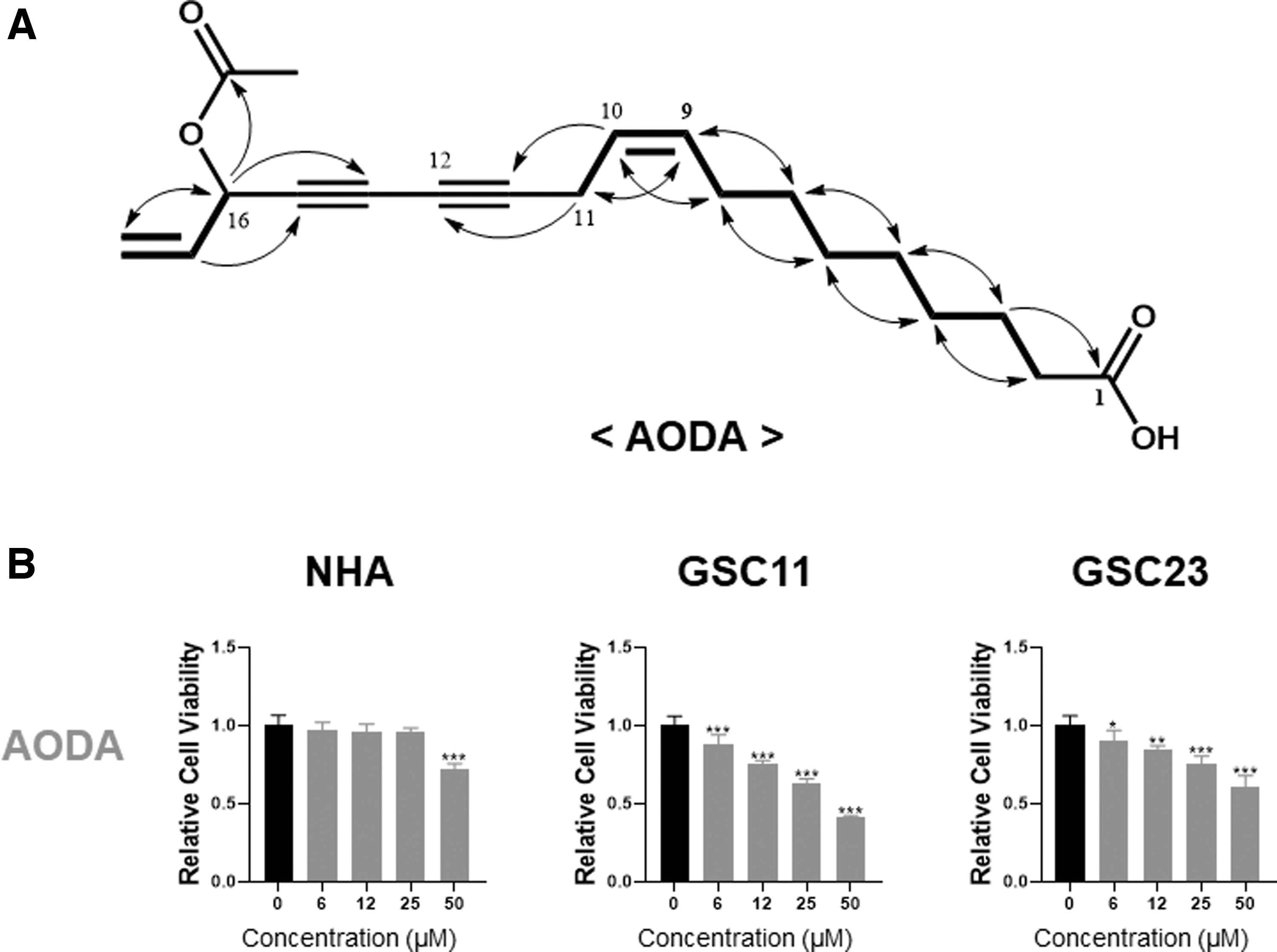
Cell viability assay after treatment with Dendropanax morbiferus analogs.
AODA specifically inhibits the viability of GSCs
Glioblastoma patient-derived GSC11, GSC23, and NHA were treated with different concentrations (0–50 μM) of AODA for 24 h. The effect of AODA on the viability of GSCs was determined by using the alamarBlue cell viability assay. As shown in Figure 1B, AODA reduced the viability of GSCs in a dose-dependent manner in vitro. However, it had almost no effect on the viability of NHA. At a concentration of about 50 μM, AODA inhibited the viability of both GSC11 and GSC23 (cell viability of 40–50%), whereas it only slightly decreased the viability of NHA (cell viability of 80%). These results suggest that AODA has a selective cytotoxic effect on GSCs, whereas it is less cytotoxic to NHA.
AODA promotes programmed cell death of GSCs
To estimate whether AODA could induce apoptosis, we performed Annexin V-FITC/PI staining using flow cytometry after GSCs were treated with AODA. After exposure to AODA at 10 and 25 μM for 24 h, only 2–8% of NHA cells were dead or undergoing apoptosis, whereas significantly higher proportions of GSC11 and GSC23 were undergoing early and late apoptosis (Figs. 2A and B). Western blot analysis showed that AODA induced an increase in cleaved Caspase 3, implying that AODA triggered apoptosis by activating protease caspase 3. Further detection revealed that the expression of an antiapoptotic protein Bcl-2 was downregulated by AODA (Fig. 2C). These results suggest that AODA might trigger apoptosis of GSCs by inhibiting Bcl-2.

Effect of AODA on cell death of GSCs.
AODA induces apoptotic cell morphology
To closely examine morphological changes of GSCs upon AODA treatment, optical diffraction tomography (ODT) was performed at an individual cell level. As shown in Figure 3A, 3D images of control GSCs had an elongated morphology in the laminin-coated plate. Conversely, AODA-treated GSCs exhibited well-known apoptotic phenotypes such as condensed intracellular components, cell shrinkage, and membrane bleb formations. Quantitative results shown in Figure 3B indicated that AODA significantly reduced cell volume, surface area, and dry mass, suggesting that AODA could cause programmed cell death of GSCs.
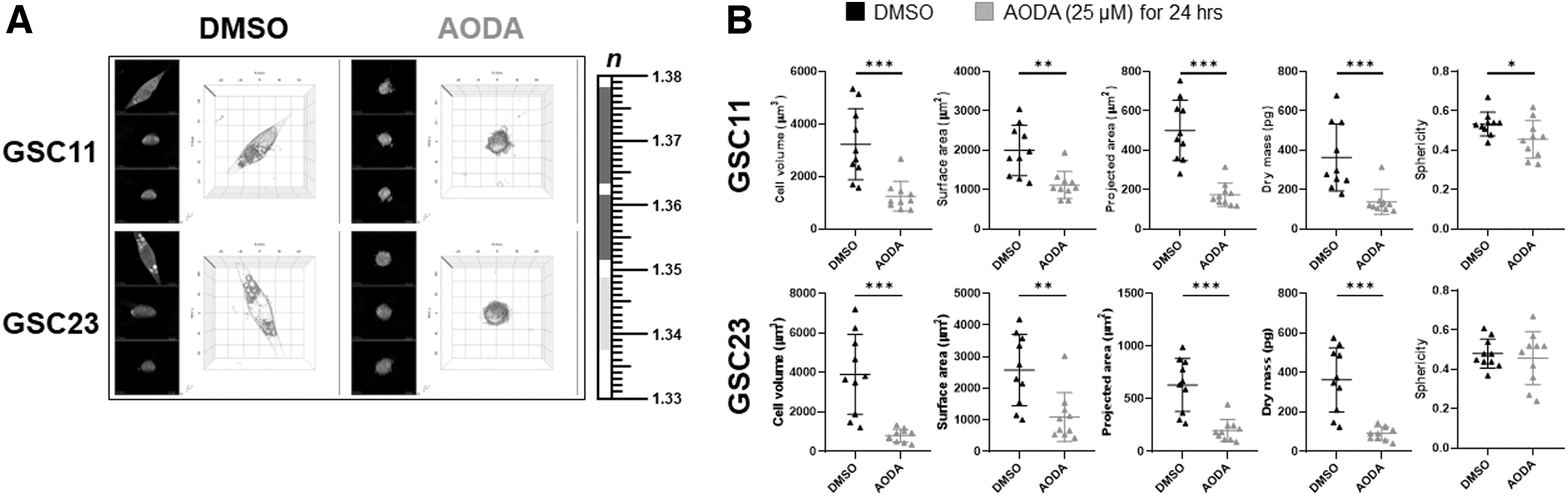
Live cell imaging using holotomography of GSCs treated with AODA (25 μM, 24 h) or DMSO.
AODA suppresses stem cell marker and tumor sphere-forming ability in GSCs
To determine whether AODA could affect GSC stemness properties, self-renewal ability of GSCs and stem cell marker CD133 mRNA expression was examined. As shown in Figure 4A, AODA dramatically reduced the self-renewal capacity of GSCs as evaluated by neurosphere production using an in vitro limiting dilution assay. In GSCs treated with AODA, sphere size was dramatically reduced more compared with control cells (Fig. 4B). Real-time PCR analysis revealed that AODA reduced stem cell marker CD133 mRNA expression (Fig. 4C). These results indicate that AODA can inhibit stem-like features of GSCs.
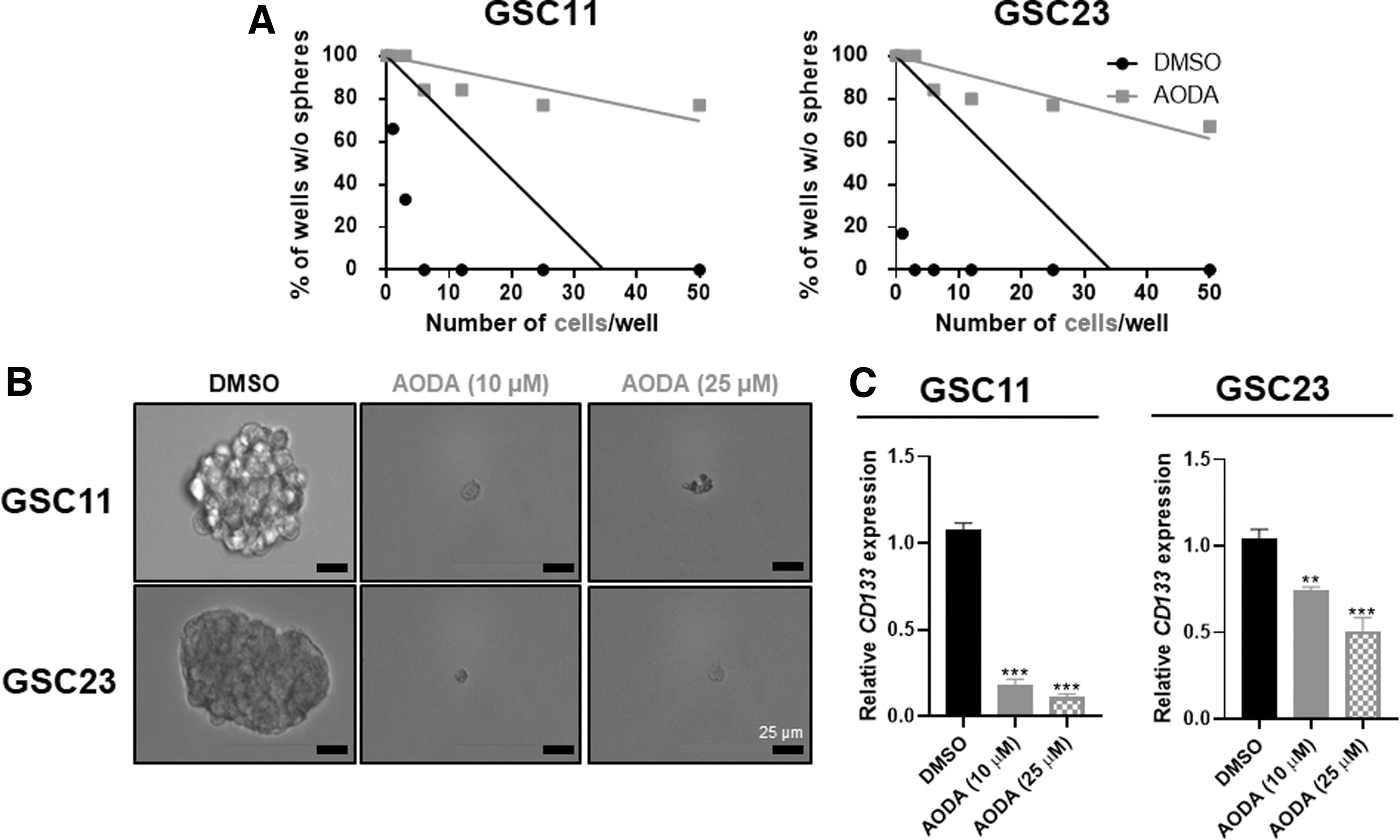
AODA inhibits GSCs stem-like features.
AODA inhibits phosphorylation of ERK and AKT in GSCs
Activation of AKT and ERK signaling pathways in the majority of GBMs represents an important node in the signaling of glioma growth, promotion of invasion, proliferation, and survival. 20,21 Therefore, we examined effects of AODA on these signaling pathways by treating GSCs with AODA. Incubation with AODA caused suppression of AKT and ERK phosphorylation as shown in Figure 5. These results demonstrate that AODA can inhibit ERK1/2 and AKT phosphorylation in AODA-treated GSCs compared with the control.
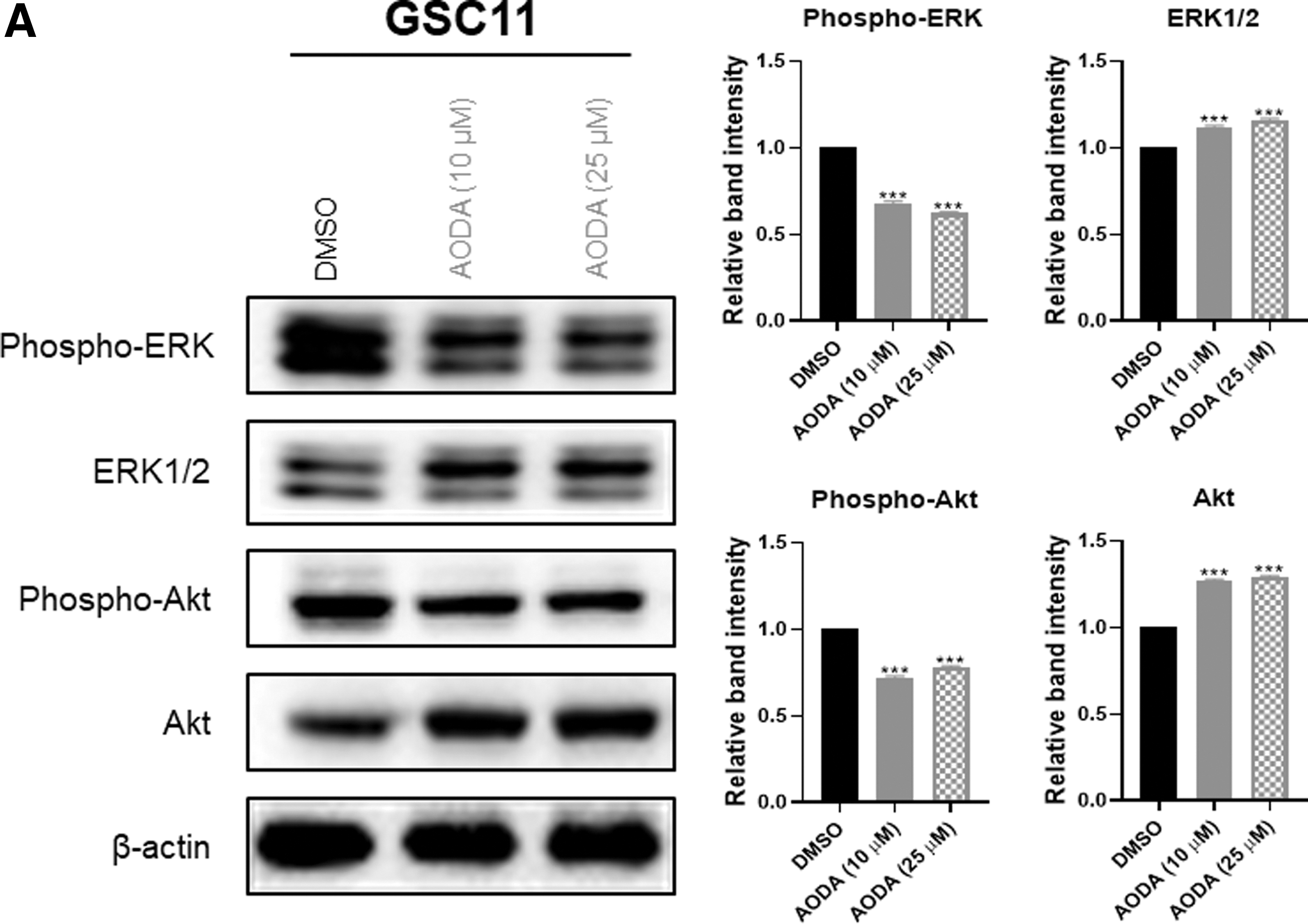
Effect of AODA on the GBM-associated signaling pathways in GSCs. Western blot analysis of GSC lines treated with AODA (10 and 25 μM, 24 h) or DMSO. Numbers represent quantified values of blots. ***P < .001. Results were obtained from three independent experiments and represented the mean ± SEM. Significant quantitative differences among groups were determined by one-way ANOVA, followed by Tukey's multiple comparison test.
DISCUSSION
Glioblastoma, one of the most lethal cancers, is extremely resistant to conventional chemotherapy and radiotherapy, with almost inevitable relapses often arising following surgery. 1,2 Tumor recurrence is caused by the presence of glioma stem cells, a small subpopulation of cells with stem-like features. 4,5 Mechanistically, GSCs are widely known to have DNA damage recovery pathways that could repair damaged DNA and lead to their survival after conventional therapy. 1,2 Therefore, research on therapies targeting GSCs is essential for developing new therapeutic strategies against malignant brain cancer.
In this study, we found that AODA isolated from leaves of D. morbifera inhibited the growth of GSC11 and GSC23 cells, but not NHA. In addition, AODA induced apoptotic cell death and morphologic features of apoptosis evidenced by ODT. Moreover, AODA inhibited the cell sphere-forming capacity of GSCs and suppressed stem-like features of GSCs. Taken together, these results suggest that AODA might be effective in targeting GSCs when it is combined with conventional therapy or used alone.
D. morbifera extracts can inhibit phospho-ERK1/2 signaling in human hepatocellular carcinoma cells, resulting in apoptosis and cell senescence. 16 Phospho-ERK1/2 signaling is an important signaling pathway for cell proliferation, differentiation, apoptosis, and drug resistance. 20,22 In this study, phosphorylation of ERK in GSC was suppressed by AODA. Phosphorylation of ERK is associated with temozolomide (TMZ) resistance mechanism. 23 Although we examined effects of AODA on GSCs directly in vitro, AODA might also contribute to eliminating GSCs using TMZ by sensitizing cells and preventing TMZ resistance.
AODA also inhibited the phosphorylation of AKT in GSC cells. AKT signaling is downstream of the serine/threonine kinase in RTK/PTEN/PI3K pathway. It has been reported that AKT is activated and phospho-AKT level is elevated in most GBM tumors and cell lines. 21 This activated and phosphorylated AKT signaling can increase the proliferation of glioma cells and facilitate apoptosis evasion and tumor invasion. Considering these points, AODA is thought to have the effect of slowing the malignancy of GBM.
Footnotes
AUTHORS' CONTRIBUTIONS
Study design: H.-J.K., Y.S.P., J.-Y.C., and S.-H.K.
Data collection: H.-J.K. and S.-S.S.
Statistical analysis: H.-J.K. and J.K.
Data interpretation: H.-J.K., H.Y.J., J.-Y.C., and S.-H.K.
Article preparation: H.-J.K., S.L., J.-Y.C., and S.-H.K.
Literature search: H.-J.K. and J.-H.M.
Funds collection: S.-H.K., J.-Y.C., and S.-S.S.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This research was supported by a grant (Nos. 2019R1I1A3A01059211 and 2022R1A5A8033794) of the National Research Foundation (NRF) funded by the Ministry of Science and ICT (MSIT), Republic of Korea. It was also supported by a grant (No. 2020183C10-2022-AA02) of the R&D program for Forest Science Technology funded by Korea Forest Service, Republic of Korea.