
Abstract
This study was performed to investigate the effects of persimmon (Diospyros kaki) on high-fat diet (HFD)-induced hepatic lipotoxicity. The compounds of persimmon water extract (PWE) were identified as gallic acid, glucogallin, 1-O-Galloyl-(2-O-acetyl)-glu, and trihydroxy-octadecadienoic acid. The PWE was ingested by C57BL/6 mice with an HFD for 8 weeks. The PWE improved glucose tolerance and suppressed weight gain by inhibiting increases in the weight of liver and adipose tissues. The results of serum biomarker analysis showed that PWE suppressed biomarkers such as liver injury and dyslipidemia. In ex vivo tests, reduction of oxidative stress and improvement of mitochondrial dysfunction were confirmed in the liver of PWE groups. In a molecular study, it was confirmed that PWE decreased lipid accumulation, insulin resistance, inflammation, and apoptosis in the liver. Finally, in a metabolite analysis of liver tissue using ultra-high performance liquid chromatography with quadrupole time-of-flight mass spectrometry (UPLC-Q-TOF/MS), it was confirmed that PWE has an effect on lipid metabolism. In particular, PWE reduced phosphatidylcholines (PCs) and lysophosphatidylcholines (lysoPCs). Notably, it is presumed that the reduction of lysoPCs and PCs in the PWE group is related to the improvement of liver dysfunction due to lipotoxicity.
INTRODUCTION
The obese population has been quickly increasing over the past century, and obesity is considered the cause of various metabolic syndromes and complications such as cardiovascular disease risks, dyslipidemia, and hypertension. 1
Liver dysfunction causes significant damage to the body because the liver is a very important organ in the body. In various studies, it has been shown that nonalcoholic fatty liver disease (NAFLD) caused by obesity is very closely related liver disease and disease progression. 2 Hepatic lipotoxicity results in the progression of NAFLD and causes various problems involved in the generation of oxidative stress, apoptosis, and inflammation, as well as insulin resistance by impairing hepatic insulin signaling. Chronic injury by hepatic lipotoxicity promotes a fibrogenic response and eventually induces liver disease. 3
Although there are a variety of drugs for treating obesity and liver disease, adverse effects are becoming a problem. 4 Several lines of evidence show that polyphenols can be effective in the prevention of degenerative diseases. Accordingly, the prevention of degenerative diseases with food containing various polyphenols is being recommended as a new strategy. 5 Accordingly, alternatives for reducing obesity using natural substances such as phenolic compounds have been proposed, and several studies have confirmed that some phenolic compounds exert an antiobesity effect. 6
Persimmon contains several phytochemicals and is suggested as a diet-based regimen to cure various diseases. 7 It is reported that persimmon fruits have an effect in improving dyslipidemia; accordingly, the possibility of using persimmon fruit for the development of preventive and therapeutic agents for dyslipidemia has been suggested. 8
This study aims to investigate the effect of persimmon water extract (PWE) on improving liver dysfunction caused by high-fat diet (HFD)-induced hepatic lipotoxicity through the analysis of metabolites in liver tissue.
MATERIALS AND METHODS
PWE preparation
The persimmon used as a sample was cultivated in July 2020 in Yeongam, South Korea, and was supplied and certified by the Korea Forest Research Institute (Suwon, Korea). PWE was analyzed using a ultra-performance liquid chromatography (UPLC) system (Waters, Milford, MA) containing Acquity UPLC BEH C18 column (2.l × 100 mm, 1.7 μm particle size). The flow rate was 0.4 mL/min and gradient conditions were applied using 0.1% formic acid in distilled water (DW) (solvent A) and 0.1% formic acid in acetonitrile (solvent B), as follows: 1% B/99% A at 0–1 min and 1–100% B at 11.5 min. The electrospray ionization (ESI) conditions are as follows: collision energy at 20–40 V, 120°C drying gas (N2), and negative ion mode.
Animal experimental design
C57BL/6 mice (4 weeks, male) were obtained from Samtako (Osan, Korea). Animal experiments were done in accordance with the Institutional Animal Care and Use Committee of Gyeongsang National University (certificate: GNU-190530-M0028). The HFD consisted of 20 kcal%/g protein, 20 kcal%/g carbohydrates, and 60 kcal%/g fat and was provided to mice for 8 weeks. The mice of PWE groups were fed PWE at 50 and 100 mg/kg body weight (PWE 50 and PWE 100) with an HFD for 8 weeks. In the eighth week, an oral glucose tolerance test was conducted. Blood was collected from the abdominal aorta, and the extracted tissues were rinsed with phosphate-buffered saline (PBS) buffer and immediately stored at −70°C.
Biochemical parameter analysis in serum
The supernatant of centrifuged blood was used to measure serum biochemicals. The serum biochemicals (glutamic oxaloacetic transaminase [GOT]; glutamine pyruvic transaminase [GPT]; blood urea nitrogen [BUN]; creatine [CRE]; total cholesterol [TCHO]; triglyceride [TG], and lactate dehydrogenase [LDH]) were analyzed with a Fuji Dri-Chem 4000i (Fuji Film Co., Tokyo, Japan). The low-density lipoprotein cholesterol (LDLC) content results from the Friedewald formula, 9 and the high-density lipoprotein cholesterol/TCHO ratio indicates as HDL-c/total cholesterol ratio (HTR). To measure the ferric antioxidant power (FRAP) of serum, serum was reacted with FRAP reagent. After 30 min in the dark, absorbance was measured at 593 nm. 10
Liver lipid accumulation and biochemicals
The measurement of total lipid from the liver was conducted in accordance with Mopuri et al. 11
Liver tissue was homogenized with PBS for the measurement of malondialdehyde (MDA) and total superoxide dismutase (SOD) levels. The pellet obtained by centrifugation was mixed with 1 × cell extraction buffer and then it was incubated on ice for 30 min. After centrifugation at 10,000 g for 10 min at 4°C, the SOD content in the supernatant was measured by using an SOD kit. The supernatants obtained by centrifugation were used to measure MDA contents. One percent phosphoric acid and 0.67%TBA were added in the supernatants and then incubated for 1 h in a heated water bath (95°C). After another centrifugation, absorbance of the supernatants was measured at 532 nm.
The method of mitochondria isolation referenced a study by Dragicevic et al. 12 The isolated mitochondria were mixed with 25 μM DCF-DA and potassium chloride-based respiration buffer and then incubated for 20 min to measure the reactive oxygen species (ROS) at fluorescence (excitation 485 nm/emission 528 nm) using a fluorometer (Infinite 200, Tecan Co., San Jose, CA, USA). To measure mitochondrial membrane potential (MMP, ΔΨm), isolated mitochondria with assay buffer were mixed with 1 μM JC-1, and after a 20-min incubation, fluorescence (excitation 530 nm/emission 590 nm) was measured. ATP was analyzed using an ATP bioluminescence assay kit (Promega, Madison, WI, USA) and a luminescence meter.
Western bolt analysis
The proteins were separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and electro-transferred to a PVDF membrane. The membrane was reacted in primary antibodies (1:1000) diluted in Tris-buffered saline, 0.1% Tween 20 (TBST) overnight at 4°C. After washing and being incubated with secondary antibody, it was treated with chemiluminescence reagent. The bands were detected by using the iB-right™ CL1000 Imaging System (Thermo Fisher, Waltham, MA, USA).
Liver metabolites analysis
Liver metabolites were analyzed in UPLC system containing Acquity UPLC BEH C18 column.
After equilibration with water containing 0.1% trifluoroacetic acid (TFA), the metabolites were separated in a gradient with acetonitrile containing 0.1% TFA at a flow rate of 0.35 mL/min for 14.5 min. The ESI conditions are as follows: positive ion mode, capillary cone voltages: 2.78 kV, sampling cone voltages: 26 V, collision energy: 10–30 eV, desolvation flow: 700 L/h at a temperature of 300°C, and source temperature: 110°C.
The metabolites were analyzed by multivariate statistical analysis using SIMCA-P+ version (Umetrics, Umeå, Sweden), and visualized using partial least-squares discriminant analysis (PLS-DA). The PLS-DA model was validated by goodness of fit (R2X and R2Y), predictability (Q2), and P-value, and cross-validated by a permutation test (n = 200).
Statistical analysis
Data are represented as mean ± standard deviation (SD). Statistical analysis was analyzed by one-way analysis of variance, followed by Duncan's multiple range test using the SAS program (Ver. 9.1 SAS Institute, Cary, NC, USA). The statistical difference (P < .05) is expressed as small letters on the bar in high order. Different small letters represent statistical difference (P < .05) of each group in the order of high to low values.
RESULTS
Identification of compounds in PWE
The compounds were identified through the database (
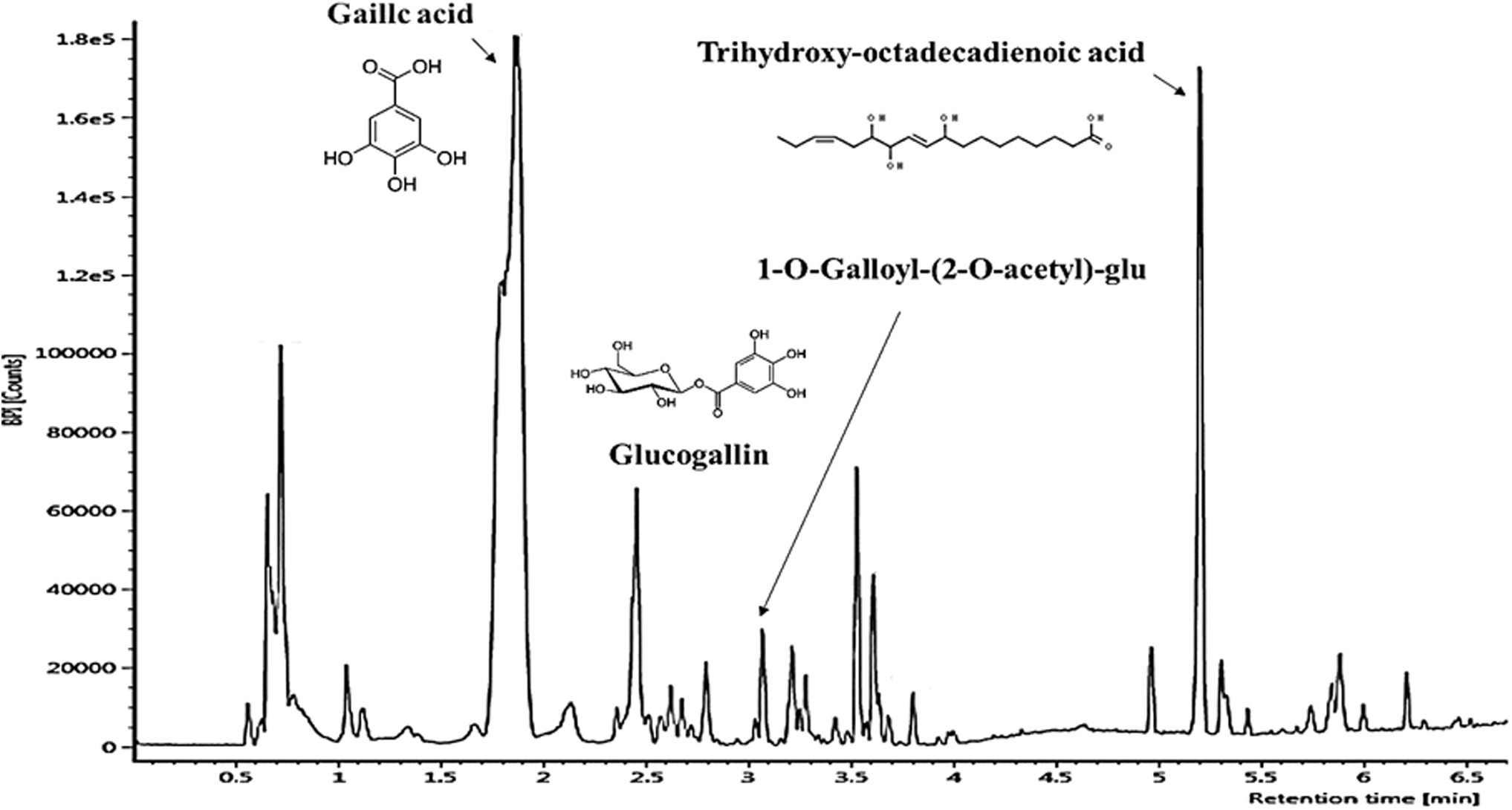
UPLC Q-TOF/MS chromatography of PWE in negative ion mode. PWE, persimmon water extract.
Body weight changes and glucose tolerance
Statistical differences in body weight between the HFD group and the PWE group were confirmed from the fifth week, and at the final eighth week, the PWE 100 group had lower body weight than the PWE 50 group (Fig. 2A).
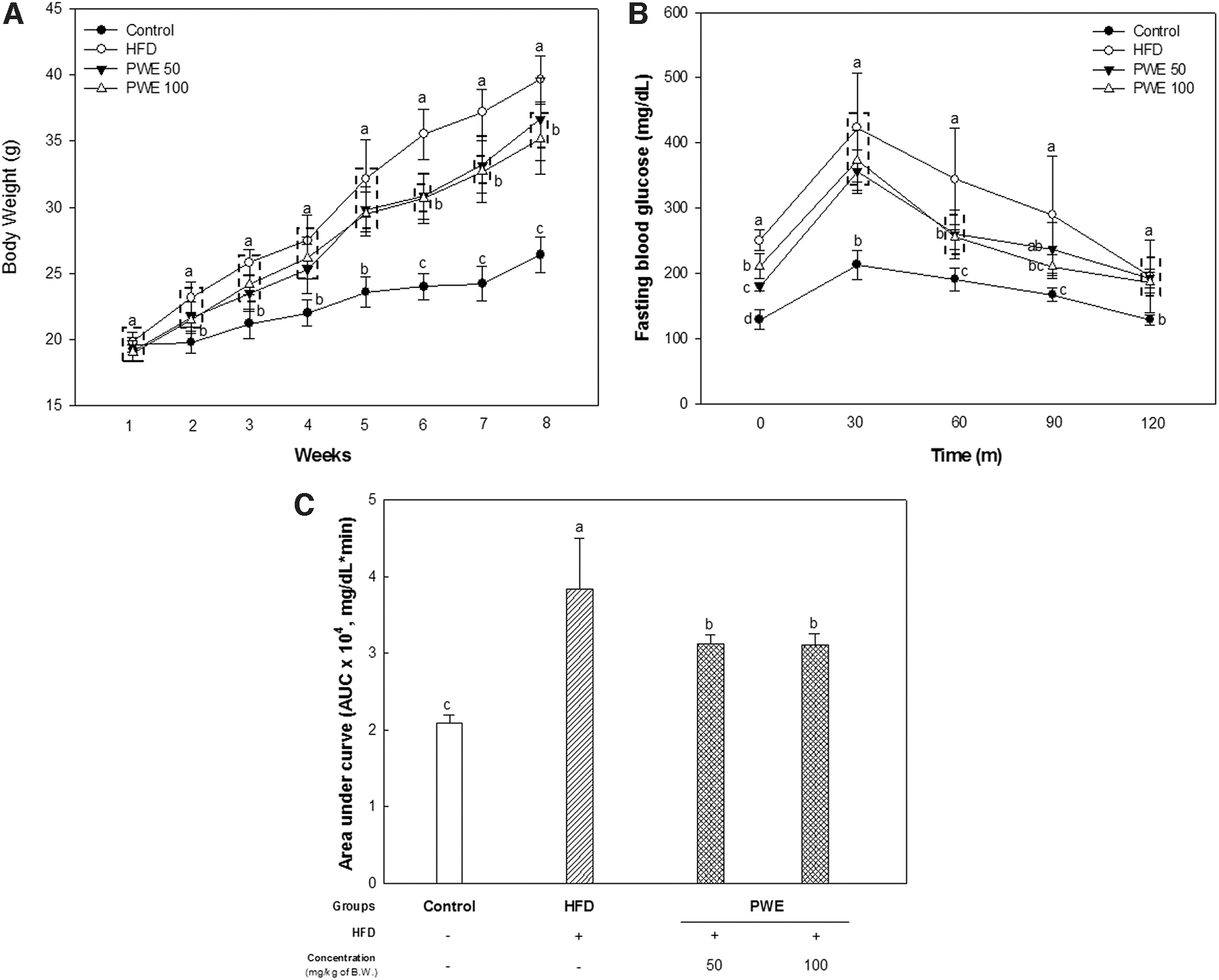
Effect of PWE on body weight changes and glucose tolerance in mice with a high-fat diet.
It was also confirmed that PWE had an effect on improving glucose tolerance. Fasting blood glucose and the glucose area under the curve used for calculating the glycemic index were decreased in the PWE (Fig. 2B, C).
The weight of adipose tissue and liver tissue and liver lipid accumulation
HFD intake significantly increased the weight of various adipose tissues as well as liver tissue. On the other hand, the weight of the PWE group's liver tissue and various adipose tissues decreased. Furthermore, liver lipid accumulation also lowered in the PWE groups than the HFD group (Table 1). Thus, the weight change of these tissues seems to be closely related to body weight change.
Effects of Persimmon Water Extract on Tissue Weight in Mice with a High-Fat Diet
The bar graphs indicate the mean ± SD (n = 7), and the statistical difference (P < .05) is expressed as the small letters on the bar in high order.
HFD, high-fat diet; PWE, persimmon water extract; SD, standard deviation.
Biomarker analysis in the serum
The GOT and GPT are commonly considered hepatic injury indicators. In addition, elevation of transaminase such as GOP and GTP is also considered a surrogate and reliable marker of NAFLD. 14 The GOT and GPT levels increased in the HFD group. However, the GOT and GPT levels in the PWE groups were lower than the HFD group. The BUN and CRE are considered renal dysfunction marker, and these are confirmed to be in the normal range (reference in Fuji Dri-Chem 4000i; CRE: 0.00–0.20 mg/dL, BUN: 12.6–16.1 mg/dL) in both PWE and HFD groups.
The levels of TCHO, TG, and LDLC in the HFD group significantly increased. On the other hand, although it did not significantly affect the LDLC level and HTR ratio, the levels of TCHO and TG were reduced in the PWE group.
The serums FRAP and LDH were also significantly different between the PWE group and the HFD group. LDH levels were increased and FRAP was decreased in the HFD group, but these changes were suppressed in the PWE group.
Measurement of liver tissue damage
Obesity and metabolic syndrome cause a disturbed balance between oxidant and antioxidant, and thereby increase oxidative stress and reduce antioxidant levels. 15 Accordingly, in the HFD group, levels of MDA, which is a lipid peroxide and marker for oxidative stress, were significantly increased. In addition, levels of SOD, a key antioxidant in the body, decreased in the HFD group. On the other hand, it was found that not only did the MDA level decrease but also the SOD content increased in the liver tissue of the PWE groups (Fig. 3A, B).
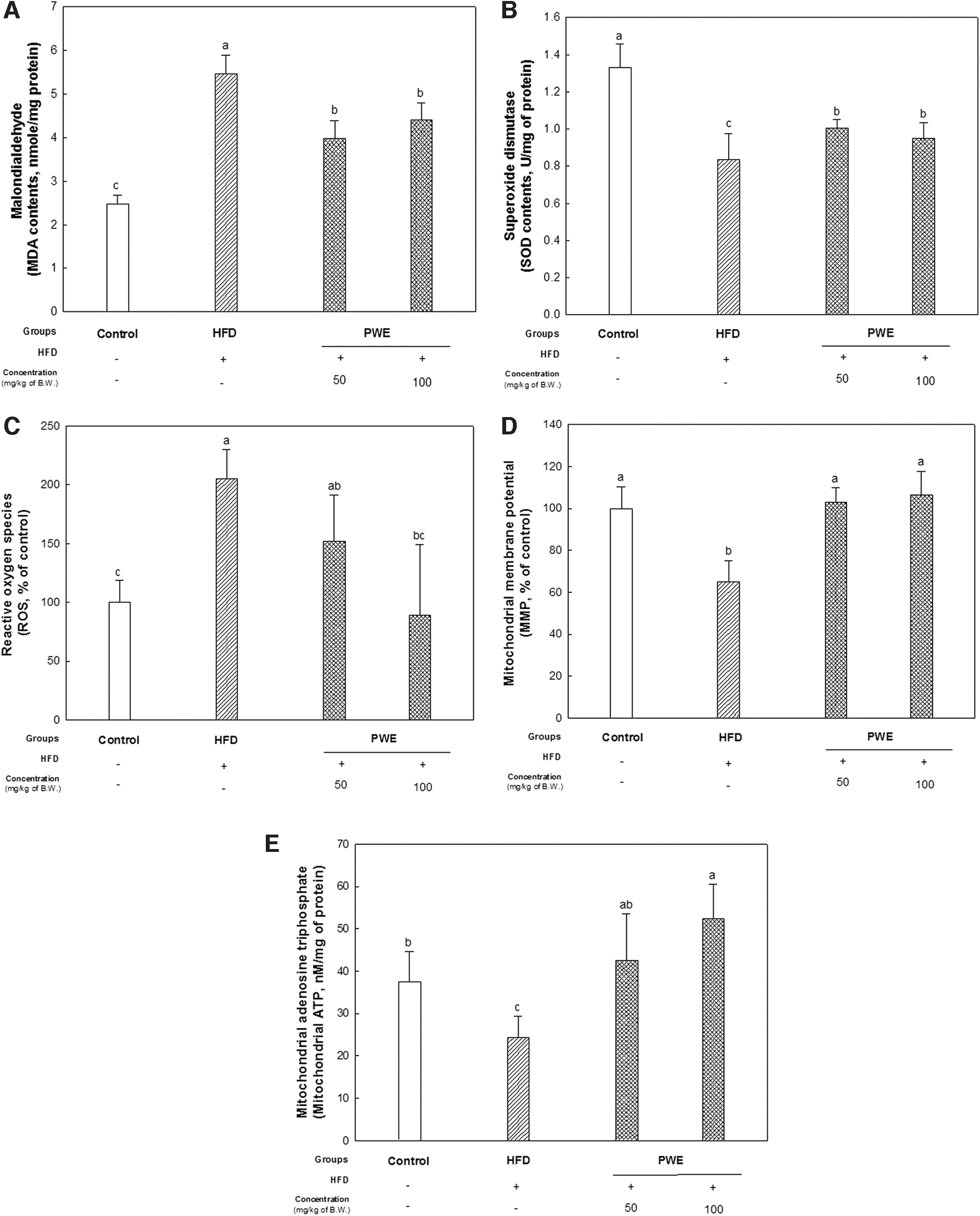
Effect of PWE on liver damage in mice with a high-fat diet.
The reduction of ROS production and elevation of MMP and ATP levels were found in the PWE groups compared with HFD group. In particular, it was observed that the mitochondrial function of the PWE 100 group recovered in a similar tendency to the control group (Fig. 3C–E).
Protein expression changes in liver tissue
The upregulation of factors related to lipid accumulation by decreasing p-AMPK and increasing SREBP-1, PPARγ, and fatty acid synthase (FAS) in the HFD group was confirmed. Protein changes related to insulin resistance such as a decrease in AKT phosphorylation and increase in p-IRS-1 (Ser307), and insulin degradation enzyme (IDE) were also upregulated in the HFD group (Figs. 4 and 5). In addition, inflammation (TNF-α and p-JNK) and apoptosis (bcl-2-like protein 4 [BAX], released cytochrome c, and procaspase 3) proteins also increased in the HFD group (Fig. 6). However, these protein changes in the HFD group tended to improve in the PWE 100 group.
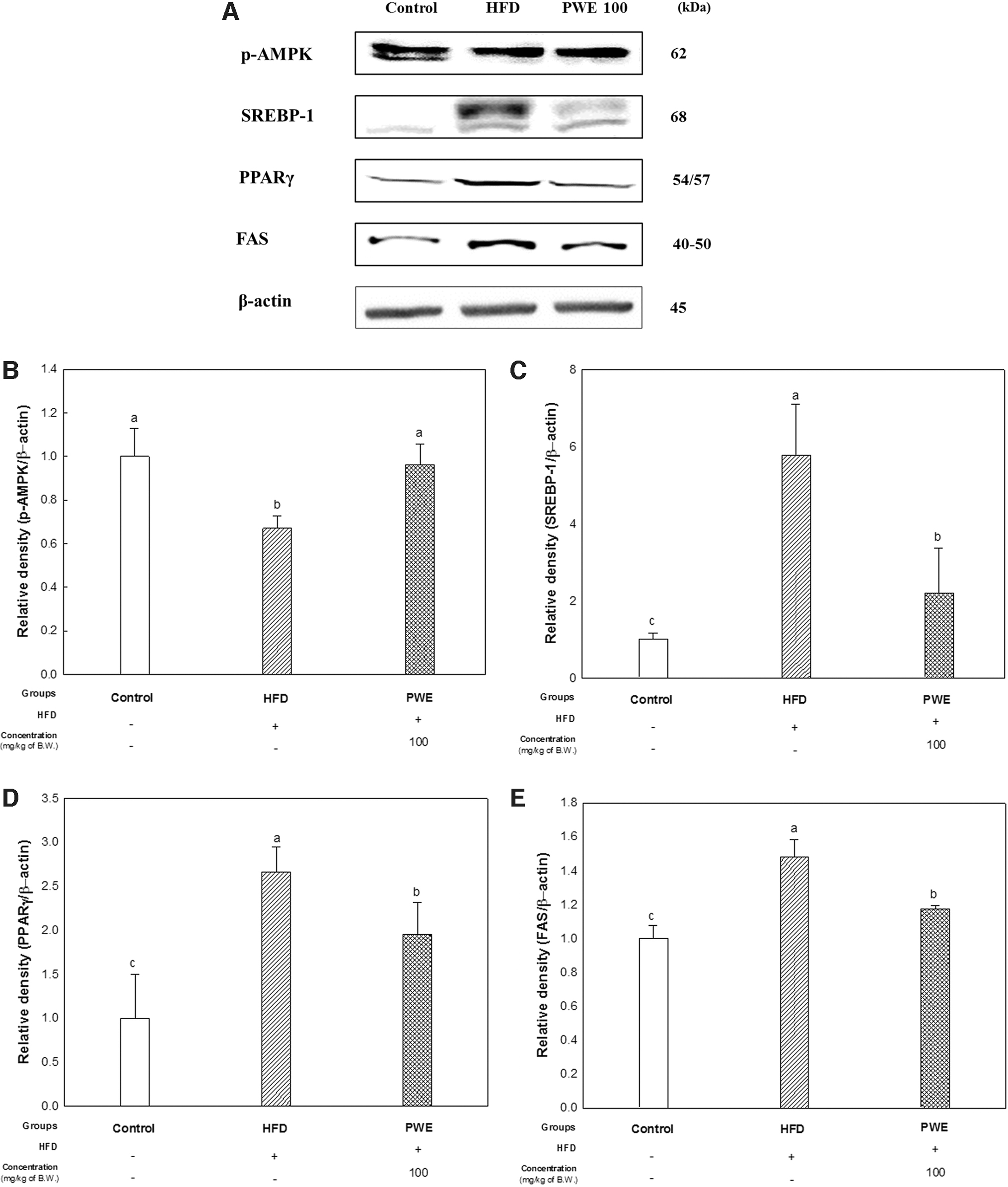
Effect of PWE on protein expression related to lipid accumulation in mice liver with a high-fat diet.
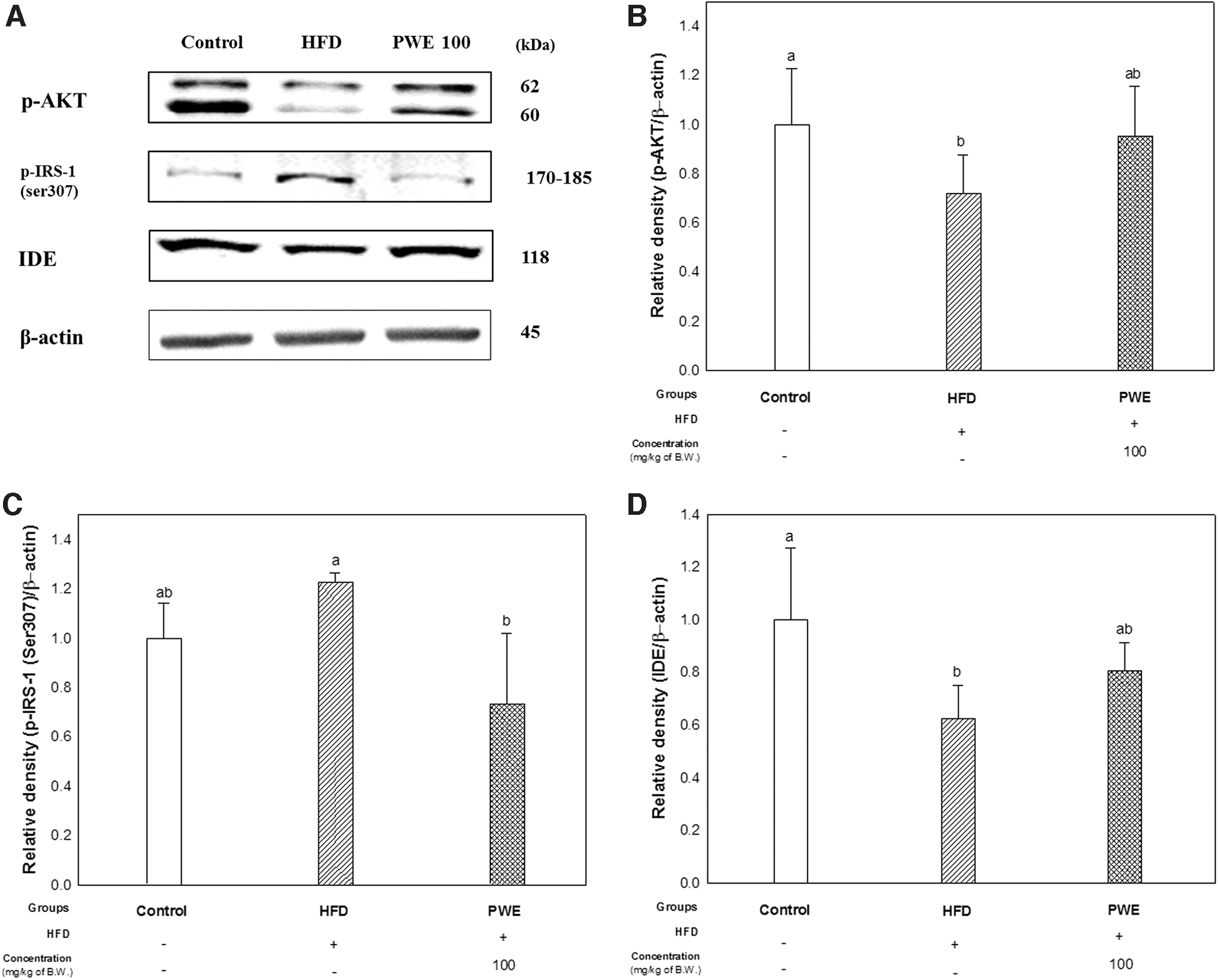
Effect of PWE on protein expression related to insulin resistance in mice liver with a high-fat diet.

Effect of PWE on protein expression related to inflammation and apoptosis in mice liver with a high-fat diet.
Metabolite analysis in liver tissue
As a result of PLS-DA using all metabolites, it was found that there was a significant difference in metabolites between all groups. As a result of verifying goodness of fit (R2X and R2Y), predictability (Q2), and P-value of PLS-DA, it was confirmed that R2X = 0.345, R2Y = 0.550, Q2 = 0.448, and P value = 6.00e-07 (Fig. 7A). Also, to verify the effectiveness of the PLS-DA model, cross-validation was performed with a permutation test (Fig. 7B). These results mean that there are statistical differences in metabolites between groups.
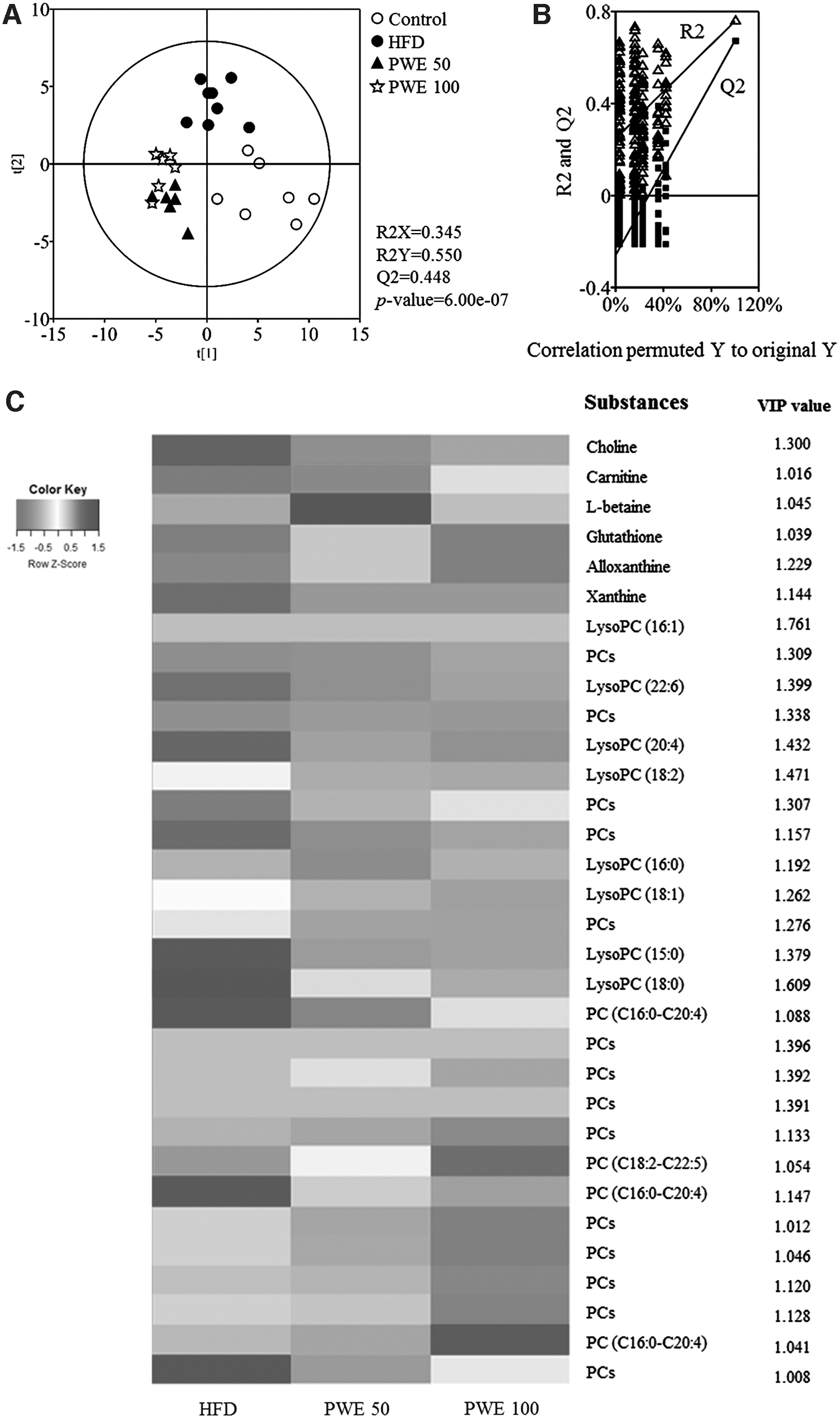
Measurement of metabolite changes in mouse liver tissue on
The heat map was made of metabolite concentration, and the selected metabolites have the value of variable importance in projection (VIP) above 1. Most of them are lipid metabolites such as phosphatidylcholines (PCs), and lysophosphatidylcholines (lysoPCs) and lipid metabolism intermediates. 16 Notably, there is a difference between the PWE and HFD groups, and most lipid metabolite-related substances were reduced (Fig. 7C). In addition, the VIP value of lysoPCs is larger than other metabolites; in other words, lysoPCs are an important factor in metabolite differences between groups.
DISCUSSION
The accumulation of excess visceral fat and NAFLD development are major characteristics of obesity, and serum GOP and GTP are reliable surrogate markers of NAFLD. 1,14 However, the increase of GOP and GTP was suppressed in the PWE group (Table 1). In addition, the phosphorylation of AMPK, a major factor related to fat accumulation in liver tissue, at the Thr172 residue increased in the PWE group (Fig. 4). AMPK is a critical player in regulating energy balance and carbohydrate and fat metabolisms, and it has been confirmed that major chronic diseases such as obesity, cancer, and diabetes cause the dysregulation of AMPK. 17
AMPK is activated when the intracellular AMP:ATP ratio increases and Thr172 residue α-subunit of AMPK is phosphorylated. The activated AMPK then stimulates fatty acid oxidation. 18 AMPK inhibits the cleavage and transcriptional activation of SREBPs, which are involved in the uptake and biosynthesis of fatty acids, and cholesterol biosynthesis by phosphorylation. In the liver, SREBPs are involved in the pathogenesis of NAFLD, nonalcoholic steatohepatitis (NASH), hepatitis, and hepatic cancer. 19 Accordingly, the expression of SREBP-1 as well as PPARγ, which stimulates lipid uptake and adipogenesis, and the expression of lipogenic enzymes such as FAS were decreased in the PWE group (Fig. 4).
The reduction of lipid accumulation is also estimated to be related to the decrease in lipid metabolism-related substances in the PWE group. Of particular note is that the VIP value of lysoPCs is high, and lysoPCs decreased in the PWE groups (Fig. 7C). According to research by Kim et al., HFD increases lipid profiles such as PCs, lysoPCs, and fatty acids. In particular, they suggested that lysoPCs can be considered potential biomarkers for the chronic consumption of HFD in nonobese individuals. 16 LysoPCs are a minor component on cell membranes, and are formed by the hydrolysis of PC, and it is observed that lysoPC levels are increased in the liver or plasma of NASH patients. 20
In addition, lysoPCs were reported to be related to lipotoxicity. Kakisaka et al. reported that lysoPCs are a key instigator of lipotoxicity. 21 Lipotoxicity is a central factor in the progression of fatty liver disease such as the inflammatory response, hepatocellular death, and activation of a fibrogenic response. 3 It is speculated that the reduction of liver lipid accumulation, including lysoPCs, suppressed hepatic lipotoxicity, and as a result reduced liver damage caused by HFD in the PWE group (Figs. 3 and 7). The antioxidant activity of PWE is also presumed to be related to the reduction of liver damage. Gallic acid, which was confirmed to be a main compound of PWE in UPLC Q-TOF/MS analysis (Fig. 1), has characteristics such as strong free radical scavenging activities and antioxidant activity.
Accordingly, it is suggested that gallic acid can protect damage caused by oxidative stress. 22 Serum LDH and FRAP indicate oxidative stress and total antioxidant capacity. 23 All these biomarkers changed by HFD were suppressed in the PWE group (Table 2). It was indicated that PWE containing gallic acid and its derivatives could reduce the increase in oxidative stress and decrease in antioxidant activity caused by HFD.
Effects of Persimmon Water Extract on Serum Biomarkers in Mice Fed a High-Fat Diet
The bar graphs indicate the mean ± SD (n = 7), and the statistical difference (P < .05) is expressed as the small letters on the bar in high order.
BUN, blood urea nitrogen; CRE, creatine; FRAP, ferric antioxidant power; GOT, glutamic oxaloacetic transaminase; GPT, glutamine pyruvic transaminase; HTR, HDL-c/total cholesterol ratio; LDH, lactate dehydrogenase; LDLC, low-density lipid cholesterol; TCHO, total cholesterol; TG, triglyceride.
Hepatocyte lipoapoptosis is also a key histological feature of NAFLD. 20 TNF-α as proinflammatory molecules regulates immune responses, inflammation, and apoptosis. TNF-α stimulates the activation of JNK by phosphorylation, and JNK pathway can promote apoptosis, which is also associated with an increase in lysoPCs. 24 According to research by Kakisaka et al., the lysoPCs phosphorylate and activate JNK, which then results in proapoptotic protein Bax activation, mitochondrial dysfunction, and caspase activation. 21 Accordingly, elevation of TNF-α- and JNK-induced apoptosis as well as mitochondrial dysfunction in the liver of the HFD group was confirmed. However, all these changes were improved in the PWE group. That is, PWE can suppress the occurrence of mitochondrial dysfunction as well as (lipo)apoptosis by inhibiting the increase in lysoPCs in liver tissue due to an HFD (Fig. 6).
Lipotoxicity also induces insulin resistance by impairing hepatic insulin signaling. 3 Accordingly, the improvement trend of the PWE group is also presumed to be related to the decrease in liver lipid toxicity (Fig. 2B, C). When phosphorylation of AKT was confirmed to evaluate insulin resistance in liver tissue, it was found that the phosphorylation of AKT increased in the PWE group (Fig. 5B). Akt is important in the insulin-induced metabolism of glucose and lipids, and the phosphorylation of Akt, which is the activated form, is an essential step in insulin signaling. 25 Insulin receptor substrate (IRS)-1 is necessary for many insulin responses, and the p-IRS-1 at serine residue is increased in insulin-resistant states.
Accordingly, p-IRS-1 (Ser307) is suggested as a molecular indicator of insulin resistance. 26 In addition, insulin resistance results in the reduction of insulin turnover by lowering IDE, eventually inducing a hyperinsulinemic condition. 27 The results of reduced p-IRS-1 at serine residues and increased expression of IDE in the PWE group mean that PWE improved insulin sensitivity (Fig. 5C, D). Also, an interesting fact is that this improvement in insulin sensitivity may also be related to the reduction of lysoPCs (Fig. 7). LysoPC imposes islet cell injury in insulin target tissue;, accordingly, it is a consequence insulin resistance. 28 In addition, it is reported that HFD-induced overproduction of PCs is also related to insulin resistance. 29 In the end, our results suggest that not only lysoPCs but also a decrease in PCs is associated with improved insulin resistance in the PWE group (Fig. 7C).
Gallic acid has already been demonstrated to be effective in improving NAFLD in various studies. 30 –32 According to research by Hsu and Yen, gallic acid is effective for suppressing HFD-induced hepatosteatosis and dyslipidemia, as well as oxidative stress. 30 In addition, it was shown that gallic acid improves impaired glucose and lipid homeostasis in HFD-induced NAFLD mice. In particular, this research demonstrated that the hepatoprotective effect of gallic acid is related to the involvement of gallic acid in various metabolic pathways such as lipid, glucose, and amino acid metabolism. 31
In a recent study, it was confirmed that gallic acid can reduce liver steatosis by inhibiting the expression of acetyl-CoA carboxylase and FAS, and the researchers suggested an important clinical application of gallic acid in the prevention and treatment of liver diseases. 32 Also noteworthy here is that gallic acid is involved in lipid metabolism. Further research is needed, but it is presumed to be related to gallic acid and the reduction of lysoPCs.
In conclusion, this study suggested that PWE can improve liver dysfunction from hepatic lipotoxicity by regulating lipid metabolism. Accordingly, we offer the possibility that PWE could be a useful treatment for suppressing the progression of NAFLD and liver dysfunction caused by hepatic lipotoxicity. However, there is one aspect to be considered. The direct correlation of gallic acid, identified as the main substance of PWE in this study, with these physiological activities needs to be verified through more diverse studies.
Footnotes
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This work was financially supported by National Institute of Forest Science, Republic of Korea (certificate: 2020031160D-00). J.Y.K., S.K.P., J.M.K., M.J.K., J.H.M., H.L.L., and H.R.J. were supported by the BK21 Program, Ministry of Education, Republic of Korea.