
Abstract
Obesity is one of the most common diseases caused by an imbalance in the intake and expenditure of energy, and it is associated with various metabolic complications. This study aimed at investigating the anti-obesity effects and mechanisms of porcine collagen peptide (PCP) using 3T3-L1 preadipocytes and high-fat diet (HFD)-fed mice. The PCP treatment significantly inhibited the adipocyte differentiation and attenuated the mRNA expression of transcription factors (CCAAT/enhancer-binding protein alpha [C/EBPα] and peroxisome proliferator-activated receptor gamma [PPARγ]) and the lipogenic gene (fatty acid synthase [FAS]) expression in 3T3-L1 preadipocytes. In the in vivo study, HFD-fed mice were fed low- (1.5 g/kg body weight/day) and high- (4.5 g/kg body weight/day) PCP for 12 weeks and compared with the normal diet-fed group and HFD-fed control group. The PCP-fed groups showed significantly lower body weight gain, white fat weight gain, serum triglycerides, and adipocyte size compared with the HFD-fed group. The changes in body fat were associated with the upregulation of adiponectin and the downregulation of leptin, C/EBPα, PPARγ, and FAS. These results suggest that PCP has the potential to reduce obesity by suppressing adipogenesis and could be applied as a functional food material.
INTRODUCTION
Obesity is a multifactorial disease caused by excessive body fat mass accumulation. It is mainly associated with the number and size of adipocytes, pre-adipocytes, and differentiated adipocytes in adipose tissue and is known to significantly contribute to various metabolic diseases such as cardiovascular diseases, hyperlipidemia, and diabetes. 1,2 According to the World Health Organization (WHO), as of 2016, more than 1.9 billion adults older than the age of 18 were overweight and more than 600 million were obese. 3 Managing obesity is crucial for controlling and reducing the risk of various metabolic diseases. The best approach that can be adopted is to keep a healthy and efficient balance between the energy intake and expenditure. 4
Adipogenesis is the proceeding of differentiation of pre-adipocytes into mature adipocytes. 5,6 Prevention or treatment of obesity can be achieved through managing adipocyte differentiation, which can be modulated by suppressing the expression of certain key transcription factors, including peroxisome proliferator-activated receptor gamma (PPARγ), CCAAT/enhancer-binding protein alpha (C/EBPα), and sterol regulatory element-binding protein-1c (SREBP-1c). 7,8 The transcription factors are then engaged in the modulation of lipogenic genes, including fatty acid synthase (FAS). 9
Collagen peptides (CP) are manufactured by the enzymatic hydrolysis of natural collagen and are widely used as food supplements because of their beneficial biological effects, such as the treatment of osteoporosis and osteoarthritis, improvement of skin moisture, elasticity, and wrinkles. 10 –12 Recent studies have identified anti-obesity properties of CP hydrolyzed from various sources. The treatment of fish CP showed anti-adipogenic activity in 3T3-L1 cells and ameliorated obesity in high-fat diet (HFD)-fed mice. 13 In other animal studies, the administration of marine CP improved the lipid profiles in mice, and ingestion of chicken feet CP elicited a reduction in visceral fat content. 11,14,15
Porcine collagen peptide (PCP), a traditional source, is produced from porcine skin. It has a low molecular weight and contains functional peptides commonly used in dietary supplements. 12 In this study, we investigated the anti-obesity effect and mechanism of PCP in 3T3-L1 preadipocytes and HFD-fed mice by regulating the adipogenic process.
MATERIALS and METHODS
Supplementation with PCP
The PCP was provided by COSMAXNBT (Seongnam, Korea), which is BODYBALANCE® P from GELITA AG. The amino acid composition of PCP is shown in Supplementary Figure S1.
Chemicals
The 3T3-L1 preadipocytes were purchased from American Type Culture Collection (ATCC, Manassas, VA, USA). High-glucose Dulbecco's modified Eagle's medium (DMEM; Welgen, Gyeongsangbuk-do, Korea), bovine calf serum (BCS; Grand Island, NY, USA), and antibiotic antimycotic solution (AA; Corning Cellgro, Manassas, VA, USA) were used for cell growth. Fetal bovine serum (FBS; Gibco), Insulin, 3-isobutyl-1-methylxanthine (IBMX), and dexamethasone (DEXA) for inducing adipogenesis and Oil Red O solution were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Cell culture and adipocyte differentiation
The 3T3-L1 preadipocytes were cultured at 37°C and in a 5% carbon dioxide atmosphere using DMEM containing 10% BCS and 1% AA. For differentiation, the cells were aliquoted at 4 × 104 cells/well in six-well plates. When the cells became 100% confluent, they were further cultured for an additional 2 days. The cultured cells were treated with an 3-isobutyl-1-methylxanthine, dexamethasone, and insulin cocktail prepared by adding 5 μg/mL insulin, 0.5 mM IBMX, and 0.25 μM DEXA to DMEM supplemented with 10% FBS and 1% AA (day 0). After 3 days, it was changed to FBS media with only 5 μg/mL insulin added (day 3). After 2 days, it was changed to FBS media (day 5) and maintained for 3 days until sufficiently differentiated (day 8). The PCP was added on day 0 and day 3 at various concentrations.
Cell viability assay
The 3T3-L1 preadipocytes were seeded into a 96-well plate at 1 × 104 cells/well, and various concentrations of PCP were added. After 24 h, a solution of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Sigma-Aldrich) in phosphate buffer solution was added and incubated for 2 h. Then, after removing the medium, the insoluble formazan product was dissolved in dimethyl sulfoxide and measured at a dual wavelength (570 and 630 nm).
Oil Red O staining
The 3T3-L1 cells were incubated with PCP for day 5, and differentiated mature adipocytes were then washed in cold phosphate-buffered saline (PBS) and fixed by 10% formaldehyde. Cells stained with 0.1% Oil Red O solution were observed under a microscope (OLYMPUS CKX41, Japan). After observation, isopropyl alcohol was added to extract the stained solution from the lipid droplets, and the absorbance was measured at 510 nm.
Animals and diets
Male C57BL/6J mice (5-week-old, 19–20 g) were purchased from Central Lab Animal Inc., (Seoul, Korea). The mice were kept at a set temperature (22 ± 2°C) and humidity (55 ± 5%) with a 12 h light–dark cycle. The mice were divided into five groups (n = 10 per group) based on body weight as follows: (1) Normal group (NC) fed a normal chow (Central Lab Animal Inc.) with saline, (2) Control group (Con) fed a 60% HFD with saline, (3) Positive control group (Catechin) fed a 60% HFD with catechin (82 mg/kg of BW/day), (4) PCP Low group (PCP-L) fed a 60% HFD with PCP (1.5 g/kg of BW/day), (5) PCP High group (PCP-H) fed a 60% HFD with PCP (4.5 g/kg of BW/day).
After a 1-week acclimation period, the mice were provided a vehicle by oral gavage for 12 weeks. The mice were given free access to the diet and water, and the HFD provided for the study was purchased from Saeronbio (Uiwang, Korea) at Rodent Diet with 60 kcal% fat. Body weights were monitored twice a week. After 12 weeks, feed was withheld from the mice for 12 h; then, the mice were euthanized using the nitrous oxide chamber; and the blood and organs were collected. Centrifugation was performed to separate the serum, and each organ collected was washed with PBS and stored at −40°C. The study was approved by the University of Suwon Institutional Animal Care and Use Committee (USW-IACUC-2021-004).
Quantitative real-time polymerase chain reaction
Total RNA was extracted using RNA iso Plus (Takara Bio Inc., Kusatsu, Japan), and cDNA was synthesized for polymerase chain reaction (PCR) analysis. Real-time PCR was conducted using the LightCycler® 96 system (Roche, Basel, Switzerland). Primer sequences are listed in Table 1.
Oligonucleotide Primers Used for Quantitative Real-Time Polymerase Chain Reaction
Cebpα, CCAAT/enhancer-binding protein alpha; Cpt1, carnitine palmitoyltransferase 1; Fas, fatty acid synthase;. Gadph, glyceraldehyde-3-phosphate dehydrogenase; Pparγ, peroxisome proliferator-activated receptor-gamma.
Western blot
Adipose tissue homogenate was subjected to quantitation of protein using radio-immunoprecipitation assay buffer and loaded onto 6% and/or 10% sodium dodecyl sulfate gels. Transferred membranes were incubated with primary antibodies overnight. The membrane was printed on X-ray film. Antibodies used in this study were FAS, peroxisome proliferator-activated receptor gamma (PPARγ; Cell Signaling, MA, USA). β-actin was used as an internal control to protein expression.
Serum lipid and adipokines levels
The levels of serum triglyceride (TG), total cholesterol, and high-density lipid cholesterol were measured using the FUJI DRI-CHEM SLIDE kit and biochemical analyzer (FUJI photo film Co., LTD, Tokyo, Japan). Serum low-density lipid (LDL) cholesterol level was measured using Friedewald's method. Adipokines were quantified using a commercial kit from R&D Systems (Minneapolis, MN, USA).
Histological analysis
The adipose tissues were fixed in 10% formalin for paraffin blocks. Sections of the paraffin-blocked tissue were cut using a microtome at a thickness of 4 μm and stained with hematoxylin and eosin (H&E). The slides were examined under an optical microscope (Olympus BX53 microscope, Tokyo, Japan).
Statistical analysis
Data are presented as mean ± standard error or as mean ± standard deviation. The results were analyzed using SPSS Software Ver.22 (IBM Corporation, Armonk, NY, USA) and were validated by one-way analysis of variance followed by Duncan's multiple-range test or independent-sample t-tests. Differences between comparison groups with P < .05 were considered significant.
RESULTS
Cell cytotoxicity of PCP in 3T3-L1 cells
The MTT assay was performed to examine the cytotoxicity of PCP in 3T3-L1 cells at various concentrations (0, 125, 250, 500, 1000, and 2000 μg/mL) for 24 h. Figure 1A and B show the results before and after adipocyte differentiation. There was no significant difference in any treated group, indicating that PCP did not induce cytotoxicity up to 2000 μg/mL.
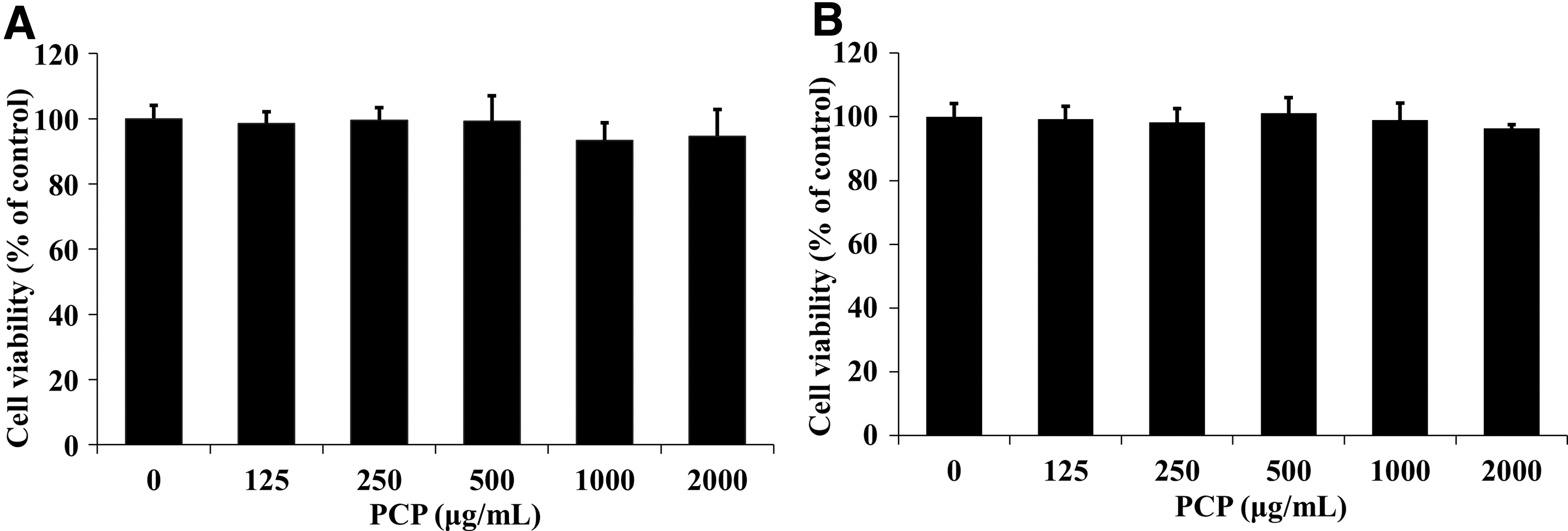
Effect of PCP on cell viability in 3T3-L1 adipocytes
Effect of PCP on lipid accumulation of 3T3-L1 cells
To examine the inhibitory effect of PCP treatment on adipogenic differentiation, Oil Red O staining was performed after differentiation of 3T3-L1 preadipocytes with treatment using various concentrations of PCP (0, 125, 250, 500, 1000, and 2000 μg/mL). As shown in Figure 2, each PCP group at different concentrations showed a significant reduction in lipid accumulation compared with the control group. These results show that PCP treatment significantly inhibited 3T3-L1 adipocyte differentiation at concentrations greater than 125 μg/mL.

Anti-adipogenic effects of PCP on 3T3-L1 adipocytes.
Effect of PCP on mRNA expressions related to adipocyte differentiation and fatty acid oxidation in 3T3-L1 cells
Figure 3 shows the mRNA expression of PCP in differentiated 3T3-L1 cells. The Cebpα levels were significantly decreased in a dose-dependent manner by PCP treatment (∼47% reduction) and Fas levels significantly decreased by ∼30% at 500 μg/mL PCP treatment compared with the control group (P < .05). Compared with the control group, Pparγ levels in cells treated with 500 μg/mL PCP were significantly decreased by 16% (P < .01). The Cpt1 level was significantly increased by 146% in the 500 μg/mL PCP-treated group compared with that in the control group (P < .05).
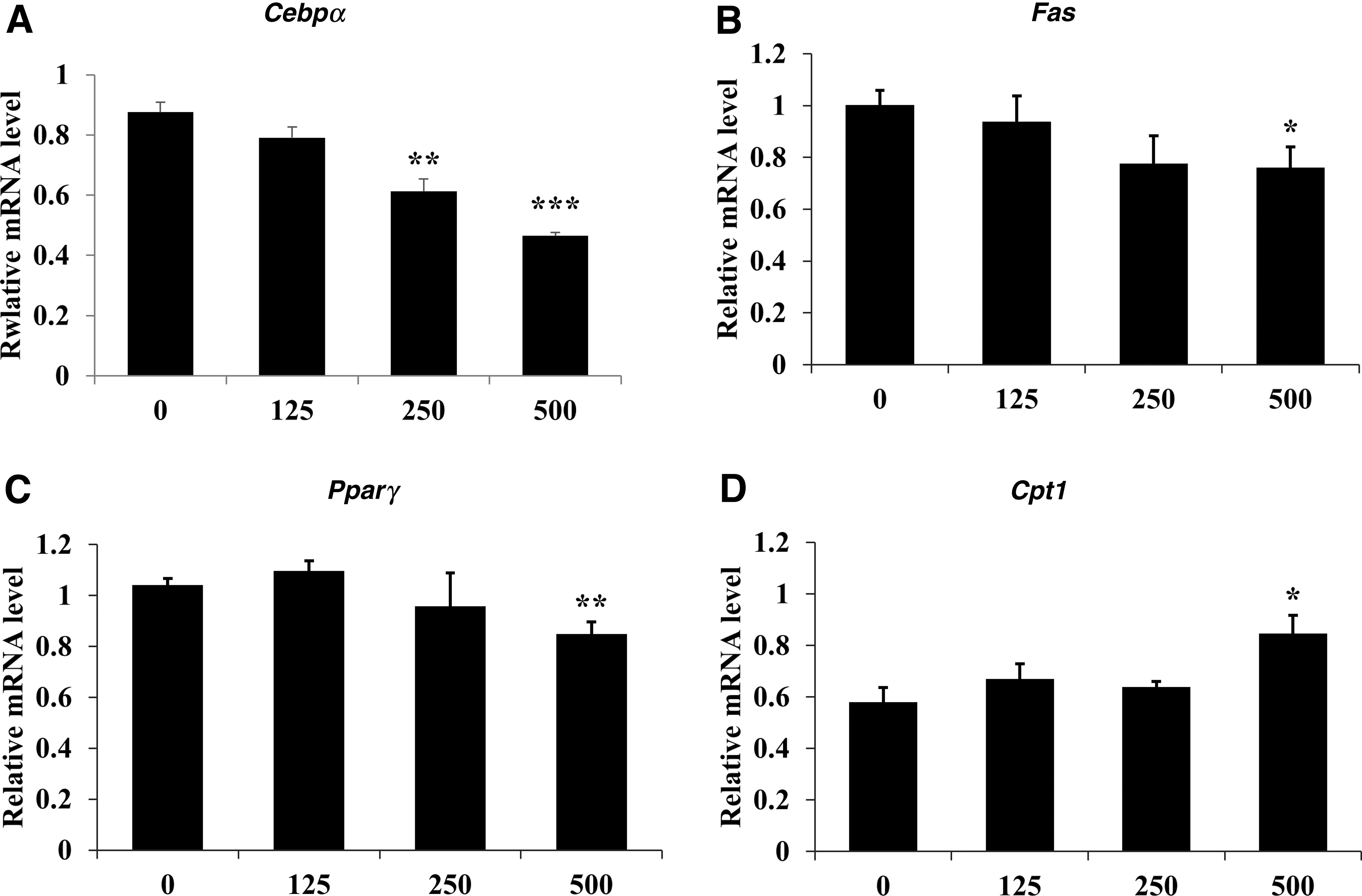
Effect of PCP on mRNA expression of
Effect of PCP on body weight gain in mice
To investigate the anti-obesity effect of PCP in HFD-fed mice, various concentrations of PCP were administered to each group of mice for 12 weeks. Figure 4 and Table 2 show the weight gains of each group after 12 weeks. The body weights of the experimental animals were measured every week. As shown in Figure 4, the body weights of the Con and obese model groups were significantly higher (135.42%) than that of the NC group (P < .01). Compared with the Con group, the PCP-H group showed significant weight loss from week 10 up to 12.73% until week 12.
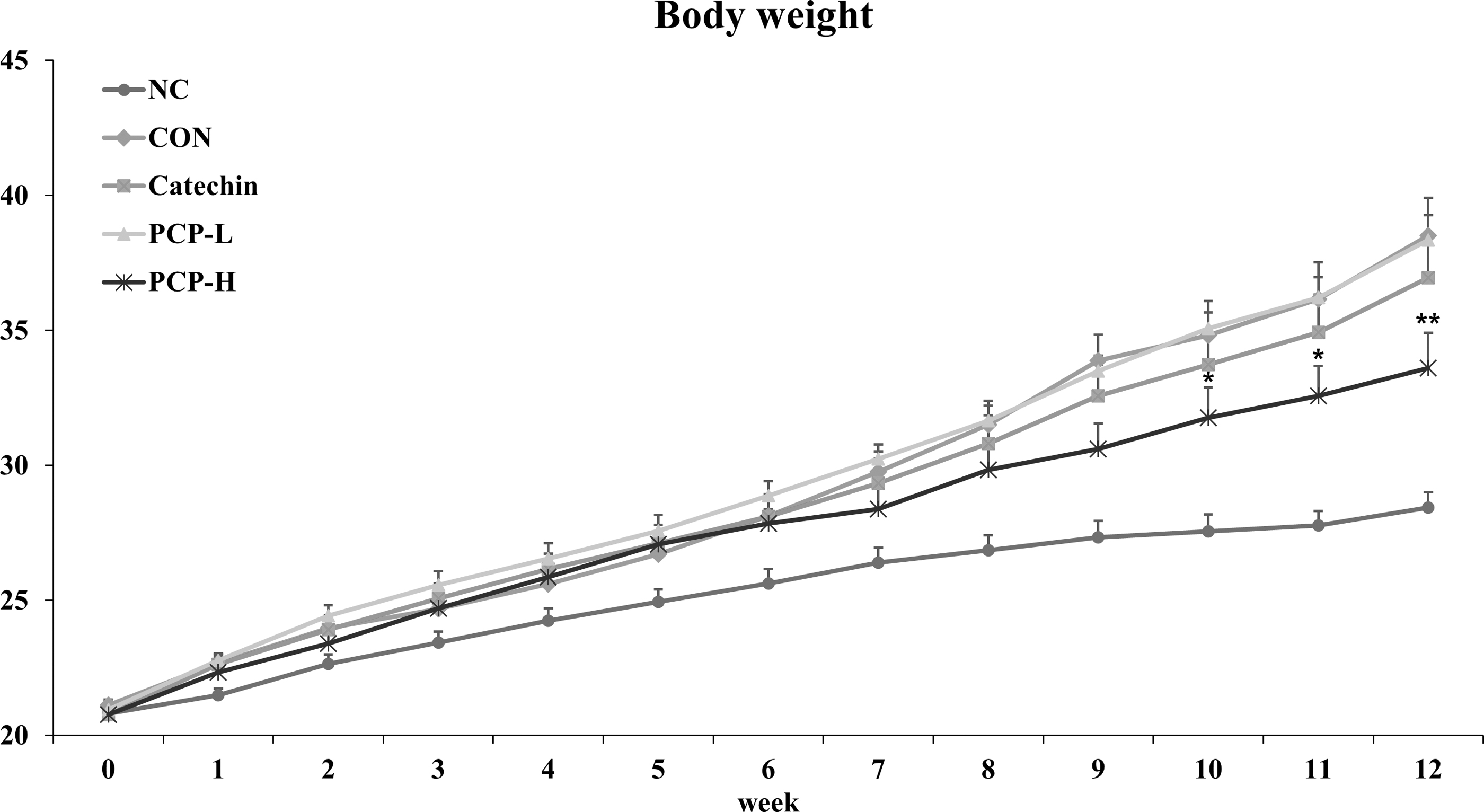
Effects of PCP on body weight in HFD-fed mice for 12 weeks. Data are presented as mean ± SE. Significant differences versus Con were denoted by *P < .05, ** P < .01. Catechin, HFD plus catechin (82 mg/kg of BW/day); Con, HFD plus saline; HFD, high-fat diet; NC, normal diet plus saline; PCP-H, HFD plus PCP (4.5 g/kg of BW/day); PCP-L, HFD plus PCP (1.5 g/kg of BW/day).
Changes in Body Weight Gain of Mice Fed Porcine Collagen Peptide for 12 Weeks
Data are presented as mean ± SE. Different alphabets show significant differences among groups (P < .05).
BW, body weight; Catechin, HFD plus catechin (82 mg/kg of BW/day); Con, HFD plus saline; HFD, high-fat diet; NC, normal diet plus saline; PCP, porcine collagen peptide; PCP-H: HFD plus PCP (4.5 g/kg of BW/day); PCP-L, HFD plus PCP (1.5 g/kg of BW/day); SE, standard error.
Effect of PCP on changes in liver, adipose tissue weight, and adipose cell size
The adipose tissue (subcutaneous, epididymal, retroperitoneal, and mesenteric) and liver weights were measured (Table 3). Liver weight showed no significant difference among the experimental groups. Adipose tissue weight in the Con group was notably higher than that in the NC group. Compared with the weights of each adipose tissue in the Con group, those in the PCP-H group were significantly reduced as follows: subcutaneous fat 36.36%, epididymal fat 31.99%, retroperitoneal white adipose tissue (WAT) 31.66%, and mesenteric fat 32.50% (P < .05).
Effects of Porcine Collagen Peptide on Liver and Adipose Tissue Weight in Mice Fed High-Fat Diet for 12 Weeks
Data are presented as mean ± SE. Different letters indicate significant differences among groups (P < .05).
WAT, white adipose tissue.
H&E staining was conducted to examine cell size in both epididymal adipose tissue (Fig. 5A) and retroperitoneal adipose tissue (Fig. 5C). As shown in Figure 5, both adipocyte sizes in the Con group were greater than those in the NC group. However, compared with the Con group, cell size of the PCP-H group was significantly smaller in both adipose tissues (Fig. 5B, D).
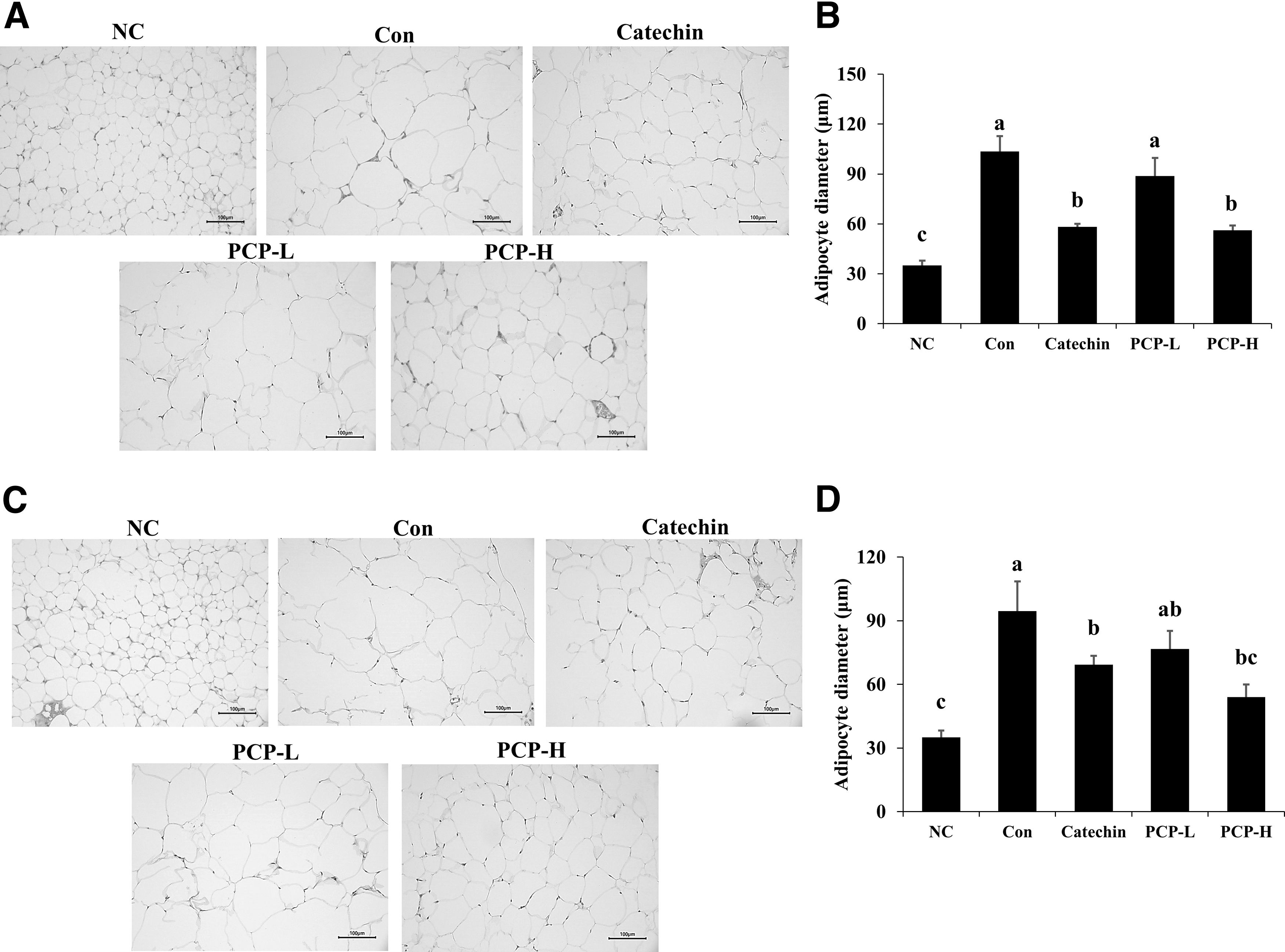
H&E staining of epididymal fat tissue and retroperitoneal fat tissue from ND- or HFD-fed mice treated with PCP for 12 weeks. Histological sections of
Effect of PCP on serum lipid levels
Serum lipid levels in the HFD-fed group were significantly higher than those in the NC group (Table 4). The serum levels of TG in the PCP-L and PCP-H groups decreased by 26.00% and 28.73%, respectively, compared with the Con group (P < .05). The serum LDL level in the PCP-L group was significantly lower than that seen in the Con group.
Effects of Porcine Collagen Peptide on Serum Lipid Profile in Mice Fed High-Fat Diet for 12 Weeks
Data are presented as mean ± SE. Different letters indicate significant differences among groups (P < .05).
HDL, high-density lipids; LDL, low-density lipids; TCHO, total cholesterol; TG, triglyceride.
Effect of PCP on adipogenesis and lipogenesis
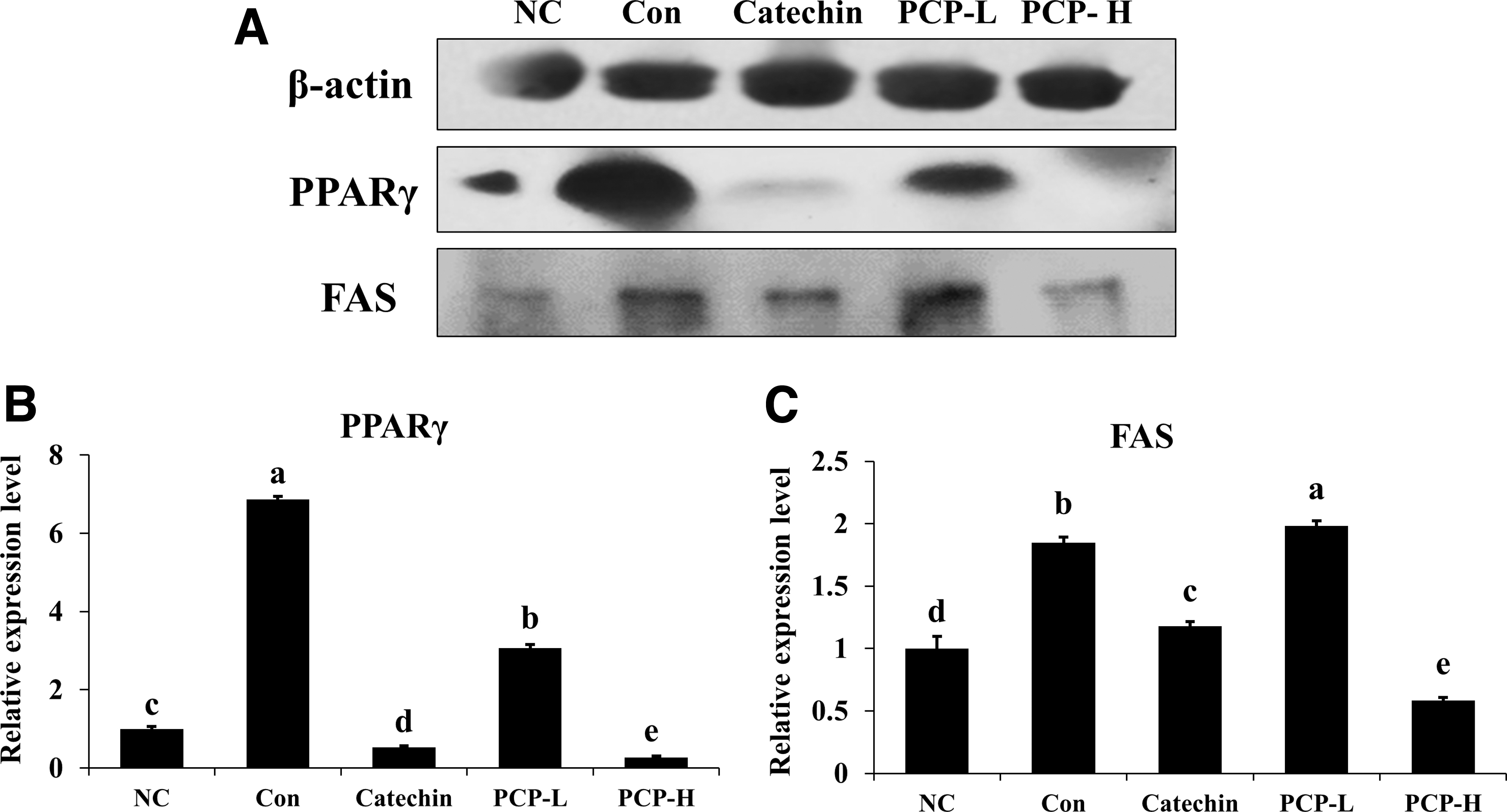
To demonstrate the effect of PCP administration on the mRNA expression related to adipogenesis in white adipose tissue of experimental animals, Pparγ, Cebpα, Fas, and Cpt1 mRNA expression levels were detected and compared (Fig. 6). The mRNA expression of Pparγ, Cebpα, and Fas in the PCP-treated groups was significantly lower than those in the Con group. In the PCP-H group, mRNA expression of Pparγ, Cebpα, and Fas was decreased by 53.03%, 69.62%, and 57.40%, respectively, compared with the Con group (P < .05). However, there were no significant differences in the mRNA expression of Cpt1. PPARγ and FAS protein expression levels in white adipose tissue were compared among each group. The protein levels of PPARγ and FAS in the PCP-H group were significantly suppressed compared with those in the Con group (Fig. 7).
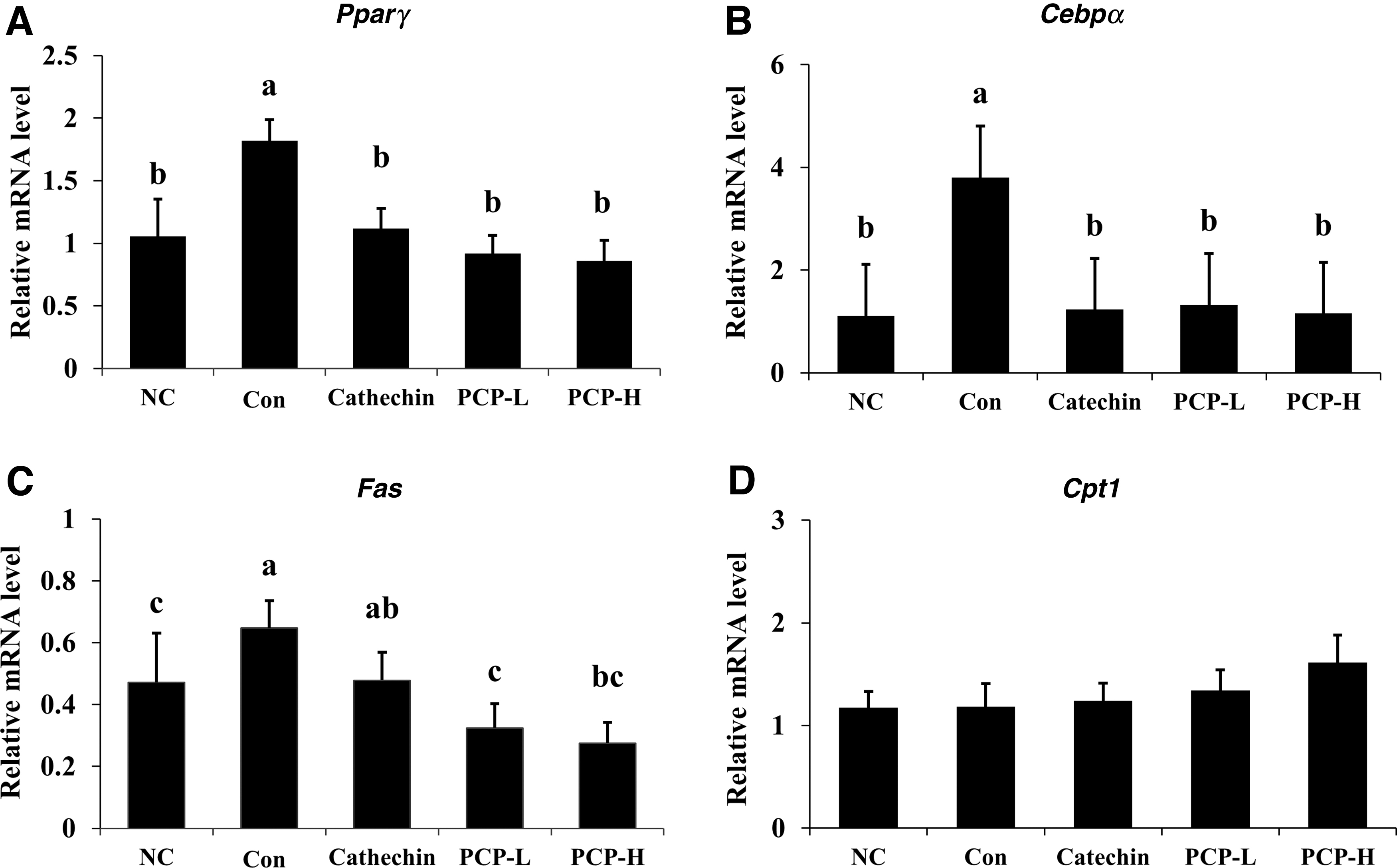
Effect of PCP on mRNA level of Pparγ, Cebpα, Fas, and Cpt1 of subcutaneous WAT. The mRNA expression of
Effect of PCP on leptin and adiponectin levels
A comparison of serum leptin levels in the Con group with those in the NC group revealed that the serum leptin level was significantly increased by HFD (Fig. 8A). The serum leptin level after PCP treatment was reduced in a concentration-dependent manner, with an approximate reduction of 37.26% in the PCP-H group. In contrast, the serum adiponectin levels of the PCP-H group were significantly increased by ∼20.09% compared with the Con group (P < .05, Fig. 8B).
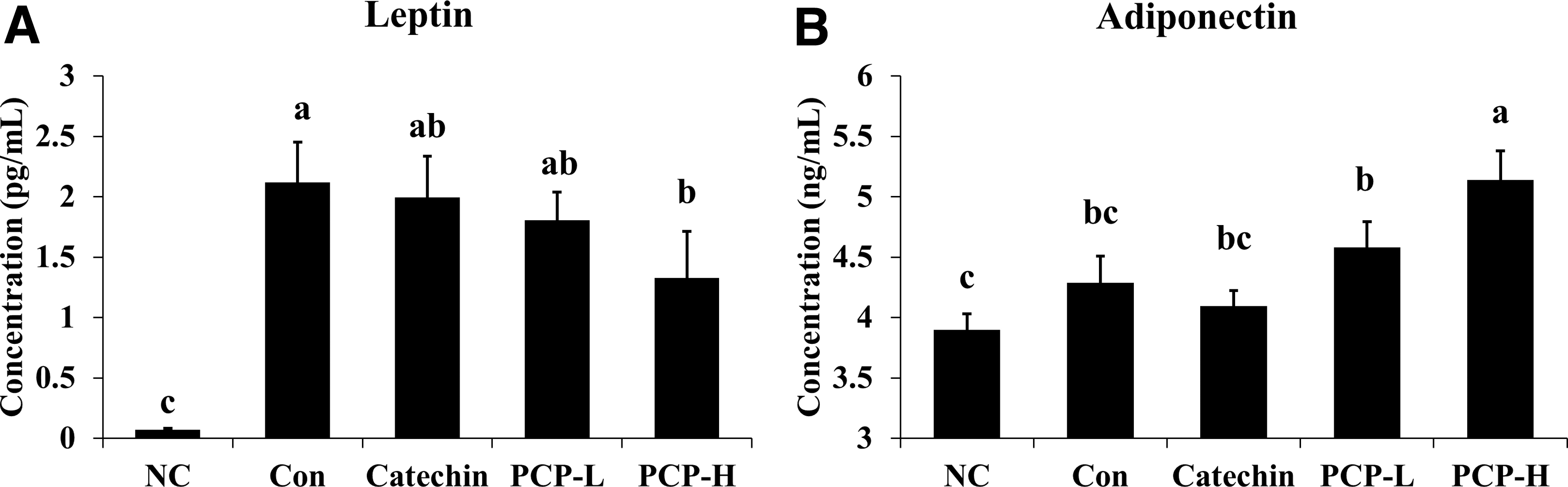
Changes in serum
DISCUSSION
Obesity, a major metabolic disease, occurs due to an abnormal increase in the presence of adipocytes in the body, resulting in the excessive accumulation of body fat. Various anti-obesity agents have been developed for treating obesity, but only a few agents have been approved based on their efficacy and appropriate safety profiles. 16 Collagen is commonly used in a number of industries, including food, pharmaceuticals, cosmetics, and leather. 17 –19 The PCP is a hydrolyzed form of collagen, which is enzymatically degraded and exhibits high solubility. 20
In this study, PCP did not show any significant levels of cytotoxicity at concentrations up to 2000 μg/mL in 3T3-L1 preadipocytes. Based on these concentrations, adipocyte differentiation with PCP treatment of preadipocyte 3T3-L1 cells was considerably suppressed in a dose-dependent manner, and transcription factors such as C/EBPα and PPARγ were significantly downregulated by PCP treatment.
In the animal study of HFD-induced obesity, the PCP-H group showed 26% of body weight loss over a period of 9–12 weeks, and significant weight loss of mesenteric, epididymal, subcutaneous, and retroperitoneal white adipose tissues. Serum TG levels in both the PCP-L and -H groups were significantly lower than those in the control group. In the PCP-L and -H groups, the expression levels of adipogenic genes, including Cebpα, Pparγ, and Fas, were greatly decreased as compared with those in the Con group.
Adipogenesis is defined as the transformation of fibroblasts from preadipocytes to adipocytes. 21 It is controlled by the activation of key transcription factors, PPARγ and C/EBPα, and it is found to be highly expressed during adipogenesis 22 ; lipid metabolizing enzymes such as FAS are regulated by these factors during adipogenesis. 21 In this study, mRNA expression and protein levels of PPARγ and C/EBPα in WAT were significantly reduced by PCP treatment both in vitro and in vivo. The mRNA and protein expression of FAS was also decreased by PCP treatment, and this result was consistent with PPARγ and C/EBPα expression. CPT-1, a fatty acid oxidation-related factor, 23 was significantly higher in the 500 μg/mL treatment in vitro, but there was no statistical difference among the groups in vivo.
If the concentration of the PCP treatment groups was higher in vivo, the expression of the mRNA level of Cpt1 would be expected to be significantly higher than that of the Con group. Recent studies have reported the anti-obesity effects of CP based on the suppression of adipogenesis and lipid metabolism. Fish CP have been shown to inhibit adipogenic differentiation in 3T3-L1 pre-adipocytes and obesity in HFD-fed mice by reducing the transcription factors C/EBPα and PPARγ. 13 Skate CP have shown anti-obesity effects by regulating lipid metabolism, such as decreasing FAS expression and increasing CPT-1 in in vivo studies. 14 Chicken CP has also induced a reduction in the visceral fat content in HFD-fed mice. 11
Obesity is the main risk factor for type 2 diabetes, which is associated with insulin resistance. The imbalance of adipokines, such as adiponectin and leptin, has been associated with metabolic syndromes, including type 2 diabetes. 24 Leptin, a reliable adipokine marker, is overexpressed in obesity and increases insulin levels, resulting in free fatty acid accumulation in the liver. 9,25,26 Adiponectin is known to be very low in obese people and negatively correlated with body fat accumulation. 25
In the present study, the leptin level in the PCP-H group was significantly lower, and the adiponectin level was higher than that in the Con group. A previous study of marine collagen peptides (MCP) in patients with type 2 diabetes showed that MCP treatment decreased serum TG and TC levels and increased adiponectin. 27 Studies on the blood-glucose-lowering effect of sturgeon CP suggest that CPs could be associated with glucose transportation or glucose absorption. 28,29
Altogether, PCP may be considered successful in suppressing adipogenesis in 3T3-L1 preadipocytes and for reducing body weight gain, white adipose tissue weight and size, and serum TG in HFD-fed mice. The anti-obesity effects are associated with the regulation of transcription factors such as PPARγ, C/EBPα, and adipokine levels. These results demonstrate that PCP has a latent ability to be developed as a useful functional food material or therapeutic agent for managing obesity.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
FUNDING INFORMATION
This study was supported by grants from the COSMAX NS Incorporation and FINE BS Company, Limited, korea.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1