
Abstract
Muscle atrophy (MA) is a case in which protein degeneration occurs excessively due to an imbalance between protein synthesis and breakdown, and is characterized by decreased muscle mass and weakened muscle strength. Despite mounting concern about MA, the number of patients with MA is increasing every year. The aim of the present study was to assess the impact of Gardeniae Fructus (GF) hot water extract on dexamethasone (DEX)-induced MA in mice. C57BL/6N mice were grouped (n = 8) as follows: Normal mice (Normal), MA mice were treated with distilled water (Control), MA mice were treated with GF 100 mg/kg (GF100), MA mice were treated with GF 200 mg/kg (GF200). For 10 days, DEX (25 mg/kg body weight, i.p.) injection was used to induce MA, and GF was administered. GF treatment restored the muscle weight decreased due to MA, and in particular, the weights of EDL+TA and Sol were significantly increased in the GF200 group. Also, it was confirmed that the swimming time was improved in the GF200 group. In addition, the expression of NADPH oxidase related to oxidative stress was significantly reduced, and protective (insulin-like growth factor I/phosphoinositide 3-kinase/protein kinase B pathway) and catabolic (AMP-activated kinase [AMPK]/sirtuin 1 [SIRT1]/proliferator-activated receptor-gamma coactivator-1α (PGC-1α)-forkhead box O (FOXO) pathway) pathways were significantly modulated. These results demonstrate that GF regulates muscle protein synthesis and catabolic pathways, and in particular, it is judged to improve MA by regulating the proteolytic AMPK/SIRT1/PGC-1α-FOXO pathway.
INTRODUCTION
Muscle is the most abundant tissue in the human body, and plays important physiological roles including breathing, locomotion, and glucose homeostasis. 1,2 Muscle is considered the important balance between protein synthesis and degradation and when protein degradation occurs excessively compared with protein synthesis due to the imbalance of protein homeostasis, muscle atrophy (MA) occurs. 2 –4 Also, the use of drugs such as glucocorticoids increases the breakdown of proteins, leading to MA. Furthermore, MA is affected by factors such as inflammation, malnutrition, aging, and a high-fat diet, and is characterized by symptoms such as muscle mass loss, muscle weakness, and disability. 4 MA is increasing in prevalence due to an aging population and an increase in life expectancy.
According to Shafiee et al.'s meta-analysis, MA was found to have a prevalence of up to 10% in the general population. 4,5 This indicates that MA is a common problem and causes substantial social and economic burden as the prevalence increases.
Protein homeostasis involves the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt)/mammalian target of rapamycin (mTOR) pathway, which is involved in muscle synthesis, and the forkhead box O (FOXO) pathway, which is involved in muscle degradation. 6 Insulin-like growth factor I (IGF-1) is known to regulate both the muscle synthesis and degradation pathways in skeletal muscle. In muscle synthesis, IGF-1 regulates the muscle anabolic activity by activating the PI3K/Akt/mTOR pathway for muscle protein biosynthesis. 7,8 PI3K/Akt/mTOR signaling plays an important role in skeletal muscle growth and differentiation. PI3K activates mTOR through phosphorylation and activation of Akt, leading to protein synthesis and muscle growth. 9 On the contrary, a decrease in Akt expression increases FOXO transcription, inducing MuRF-1 and MAFbx, which leads to the atrophy of skeletal muscle. 10 Therefore, the balance between the PI3K/Akt/mTOR and FOXO pathways in MA is considered important.
Gardeniae Fructus (GF) (Zhi-zi in Chinese) is the dried fruit of Gardenia jasminoides Ellis of the Rubiaceae family, and in traditional Korean medicine, GF is known as a yellow dye. It has also been reported to have pharmacological properties such as hepatoprotective, anti-inflammatory, antioxidant, and anticell death. 11 Lv et al. reported that gardenia ameliorates age-related diseases by protecting against inflammation, mitochondrial dysfunction, and oxidative stress (OS). 12 Geniposide, the main component of GF, is known to have various pharmacological effects such as antioxidant stress, anti-inflammation, and antiapoptosis. 13 Pan et al. reported that geniposide, the main component of gardenia, improved skeletal muscle fibrosis. 14 In addition, it has been reported that geniposide regulates glucose homeostasis in skeletal muscle. 15,16 As such, many studies have reported that geniposide has a positive effect on skeletal muscle.
The purpose of the present study was to evaluate the efficacy and mechanism of GF on muscle synthesis and degradation in MA. In the present study, MA was induced by injection of dexamethasone (DEX) in C57BL/6N mice, and the effect of GF on the IGF-1/PI3K/Akt and AMP-activated kinase (AMPK)/sirtuin 1 (SIRT1)/peroxisome proliferator-activated receptor-gamma coactivator-1α (PGC-1α)-FOXO pathways in MA was investigated.
MATERIALS AND METHODS
Materials
DEX (Cat. No. D2915) was obtained from Sigma Aldrich Co. (St Louis, MO, USA). NADPH oxidase (NOX)2 (SC-130543), p47phox (SC-17845), IGF-1 (SC-518040), PI3K (SC-1637), p-mTOR (SC-293133), mTOR (SC-517464), p-p70S6K (SC-8416), p70S6K (SC-8418), SIRT1 (SC-15404), PGC-1α (SC-13067), MuRF-1 (SC-398608), MAFbx (SC-166806), and β-actin (SC-47778) were obtained from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA), and p-PI3K (#4228), p-Akt (#9275), Akt (#9272), p-AMPKα (#2531), AMPKα (#2532), PGC-1α (SC-13067), p-FOXO3a (#9465), and FOXO3a (#12829) were obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA). Goat anti-mouse and goat anti-rabbit immunoglobulin G (IgG) horseradish peroxidase (HRP)-conjugated secondary antibodies were purchased from GeneTex, Inc. (Irvine, CA, USA). Isotroy was purchased from Troikaa Pharmaceuticals, Ltd. (Ahmadabad, India).
Preparation of the plant material
GF was a medicinal herb produced in Goheung, Jeollanam-do, in 2019, and was obtained from Bonchowon (Yeongcheon, Gyeongsanbuk-do). A voucher herbarium specimen was verified at the College of Korean Medicine in Daegu Haany University. The dried GF (100 g) was extracted by 10 × volume boiling water (100°C) for 2 h. The solvent was concentrated by evaporating in vacuo and then dried completely using a freeze dryer to obtain a powder (the yield rate; 24.6%).
GF analysis by ultra performance liquid chromatography chromatogram
The GF hot water extract (1 mg) was dissolved in 2 mL of 100% methanol. We injected 3 μL of the sample into a Waters Acquity UPLC system (Waters®, Milford, MA, USA) using a reversed-phase C18 column (Phenomenex HPLC part: Luna 3 μ C18(2) 100 Å, 150 mm × 4.6; Torrance, CA, USA). The mobile phase was composed of 5% acetonitrile in deionized water with 0.05% formic acid (A) and 100% acetonitrile with 0.05% formic acid (B). The gradient conditions were as follows: A:B = 90:10 (0 min)/90:10 (2 min)/80:20 (3 min)/80:20 (6 min)/0:100 (8 min)/0:100 (10 min)/90:10 (11 min)/90:10 (14 min). The flow rate was 0.7 mL/min with UV absorption monitoring at 254 nm.
Quantification of geniposide in the extract was performed by peak area measurement. A calibration curve for geniposide was made at the concentrations of 6.25, 12.5, 25, 50, 100, 200, and 400 μg/mL with an injection volume of 3 μL each and 5 replications. The amount of geniposide in the extract is analyzed to be 176.625 ± 0.809 mg/g (Fig. 1).

UPLC profile of geniposide in the GF at 254 nm.
Induction and treatment of MA
The animal experiment protocol referred to Shen et al.'s study, 16 and was performed with the approval of the Animal Care and Use Committee of Daegu Haany University (Approval No. DHU 2022-012). The 8-week-old male C57BL/6N mice were obtained from DBL (Eumseong, Korea). After 1 week of adaptation (environmental conditions: 12-h light/dark cycle, controlled temperature [22°C ± 2°C], and humidity [50% ± 5%]), 32 C57BL/6N mice were randomly divided into four groups (n = 8) as follows: (1) Normal, normal group; (2) Control, MA mice were treated with distilled water; (3) GF100, MA mice were treated with GF hot water extract (100 mg/kg body weight); and (4) GF200, MA mice were treated with GF hot water extract (200 mg/kg body weight).
The mice were injected with DEX (25 mg/kg body weight, i.p.) once a day for 10 days to induce MA. At the same time, GF hot water extract was orally administered for 10 days. On the 10th day, the mice were anesthetized by Isotroy inhalation anesthesia (induction, 4% isoflurane; maintenance, 2% isoflurane) for 5–7 min and sacrificed by inhalation anesthesia (isoflurane, Telangana, India). On the day of the end of the experiment, blood was collected from the abdominal vein after anesthesia. Serum was obtained by centrifugation (Hanil Scientific Inc., Gimpo, Korea) of blood for 10 min (at 4000 rpm, 4°C) and stored in a −80°C freezer for subsequent biochemical evaluation and ELISA.
Muscle tissues (Quad, quadriceps; Gast, gastrocnemius; EDL, extensor digitorum longus; TA, tibialis anterior; Sol, soleus) were collected and weighed with a precision balance (Series 360 ES Semi-micro, Precisa, Switzerland) to determine the muscle weight.
Histological examination
Muscle (Gast) tissues were fixed in 10% formalin, and then processed for embedding and sectioning at the Kyeongbook National University Hospital BMRI (Daegu, Korea). Muscle sections (3 μm thickness) were stained using hematoxylin and eosin (H&E), which were followed by the standard protocol. The images were captured using an Olympus BX51 microscope (Olympus Co., Ltd., Tokyo, Japan; 200 × and 400 × magnifications) and then analyzed using the I-Solution Lite software program (IMTechnology, Vancouver, BC, Canada). Quantification analysis of each positive signal for histological examination image was obtained from the randomly selected sections of at least 3 fields of each sample using the ImageJ 1.8.0 software (U.S. National Institutes of Health, Bethesda, MD, USA).
Forced swimming test
The mice in all the groups, except the normal group, were subjected to swimming practice 5 min/day at days 1, 4, 7, and 10 from the start of the experiment. For swimming practice, mice were placed in an acrylic plastic pool (120 × 70 × 50 cm) filled with fresh water (23°C ± 1°C) to a depth of 25 cm. The mice in all groups except the normal group underwent a forced swimming test before sacrifice. Exhaustion was defined as loss of coordinated movement and failure to rise to the surface within 5 s, and the swim time was recorded.
Measurement of glutamic oxaloacetic transaminase and glutamic pyruvic transaminase levels
The assays of glutamic oxaloacetic transaminase (GOT) and glutamic pyruvic transaminase (GPT) in serum were carried out using a GOT and GPT assay kit (Asan Pharmaceutical Co., Ltd., Seoul, Korea).
Western blotting
Muscle (Gast) tissues were lysed with cold RIPA buffer (Thermo Fisher Scientific, Waltham, MA, USA) containing freshly added Halt Protease & Phosphatase Inhibitor Cocktail and ethylene-diamine-tetraacetic acid solution on ice for 20 min. Whole-cell lysates were centrifuged (Hanil Scientific Inc., Gimpo, Korea) for 20 min (at 13,000 rpm, 4°C) and the supernatants were transferred into new tubes. Lysates were stored at −80°C before the analysis. For the estimation of NOX2/p47phox/IGF-1/p-PI3K/PI3K/p-Akt/Akt/p-mTOR/mTOR/p-S6K/S6K/SIRT1/p-AMPK/AMPK/PGC-1/p-FOXO3a/FOXO3a/MuRF-1/MAFbx/β-actin (1:1000), 12 μg of protein was electrophoresed through 8–14% of sodium dodecyl sulfate–polyacrylamide gel (SDS-PAGE). Separated proteins were transferred to a nitrocellulose membrane, blocked with 5% (w/v) skim milk solution for 1 h, and then incubated with primary antibodies (overnight at 4°C), respectively. After the blots were washed, they were incubated with the anti-mouse and anti-rabbit IgG HRP-conjugated secondary antibodies (1:3000) for 2 h at room temperature.
Each antigen–antibody complex was visualized using ECL Western blotting detection reagents and detected by chemiluminescence with Sensi-Q 2000 Chemidoc (Lugen Sci Co., Ltd., Gyeonggi-do, Korea). Band densities were measured using the ATTO Densitograph Software (ATTO Corporation, Tokyo, Japan). The protein levels of the groups are expressed relative to those of the normal rat (represented as 1). 17
Statistical analysis
In vivo values are expressed as means ± standard deviations. Statistical comparisons were analyzed by one-way ANOVA tests followed by the least significant difference (LSD) test using SPSS (version 26.0, IBM, Armonk, NY, USA). Values of P < .05 were considered significant.
RESULTS
Body weight and muscle changes to DEX-induced MA
To verify the effect of GF in ameliorating MA, a DEX-induced MA mouse model was generated. First, as a result of observing changes in body weight, the Normal group increased by 1.37 ± 0.57 g during the experiment period, but the body weight decreased by 0.14 ± 0.78 g in the Control group. On the contrary, in the GF administration group, there was an increase by 0.09 ± 1.25 g and 0.14 ± 0.27 g, respectively (Table 1). Also, as a result of confirming the effect of GF on the muscle weight of mice, the DEX-induced control group showed significantly lower muscle weight than the Normal group (Quad, Gast, Sol, P < .001; EDL+TA, P < .01), and it was higher in the GF-treated group (Fig. 2A).
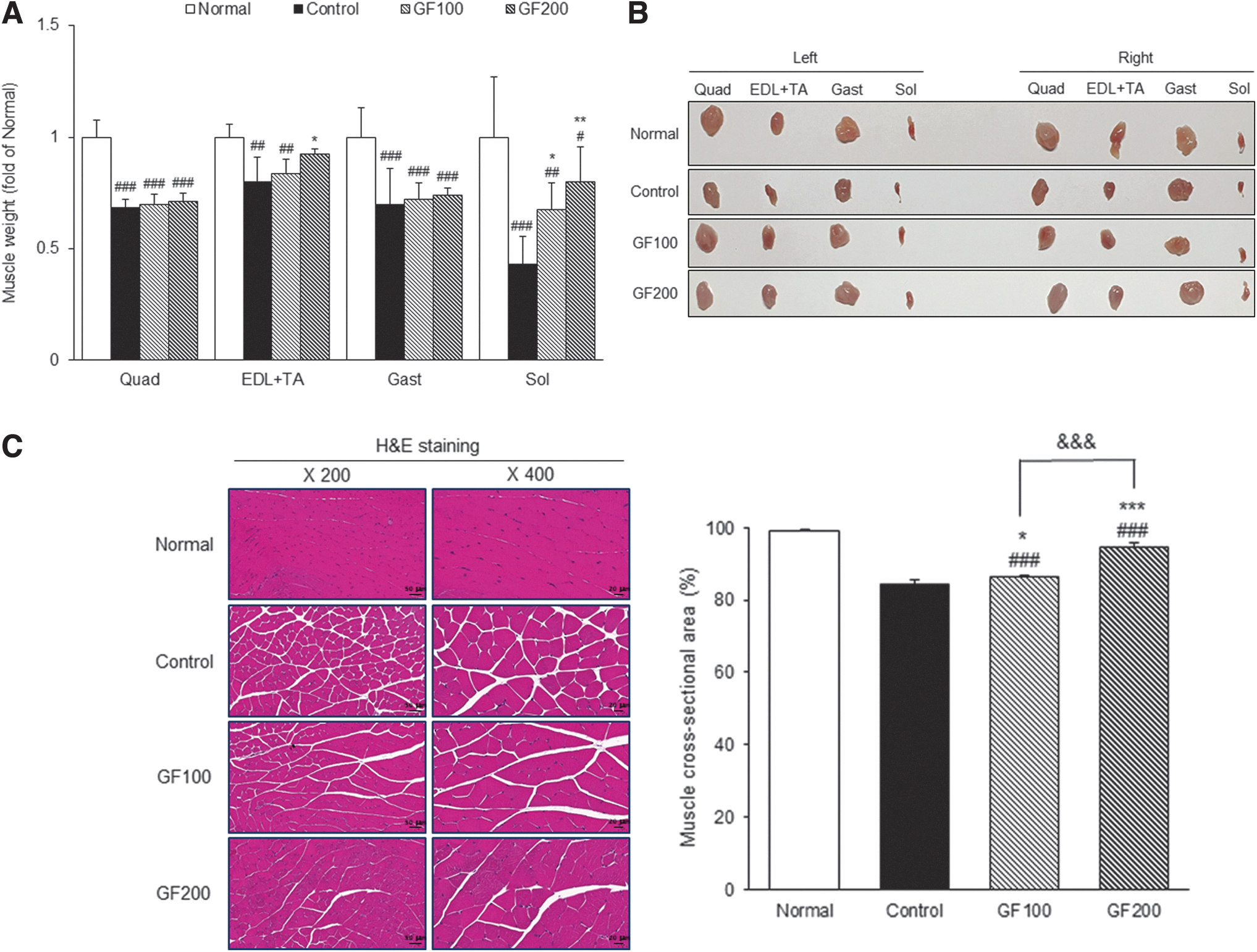
Effects of GF on DEX-induced MA.
Body Weight and Percent of Muscle Mass
Normal, normal group; Control, muscle atrophy-induced mice were treated with distilled water; GF100, muscle atrophy-induced mice were treated with Gardeniae Fructus hot water extract 100 mg/kg body weight; GF200, muscle atrophy-induced mice were treated with Gardeniae Fructus hot water extract 200 mg/kg body. Data are mean ± SD (n = 8). Significance: # P < .05, ## P < .01, ### P < .001 versus the Normal group, * P < .05 versus the Control group.
SD, standard deviation.
Moreover, as a result of comparing muscle mass as a percentage of body weight, the Control group was significantly lower than the Normal group (18.91% ± 3.45% vs. 9.81% ± 3.04%), and significantly higher in the GF200 group compared with the Control group (Table 1). When each muscle was collected and visually examined, it was confirmed that MA was induced by the DEX-treatment, and that MA was alleviated through the GF treatment, and the size of the muscle was larger than that of the control group (Fig. 2B). To confirm the effect of GF on muscle damage, the Gast sections were evaluated by H&E staining. The cross-sectional area of Gast muscle was lower in the DEX-treated groups compared with the Normal group (98.99% ± 0.41% vs. 84.51% ± 1.08%), whereas the GF-treated group was significantly higher by as much as the Normal group level (Fig. 2C).
Improvement of swimming time by GF on DEX-induced MA
To investigate the effect of GF on muscle function, the exercise performances of mice were measured, and the results are shown in Figure 3. The Control group had a significantly lower swimming time than the Normal group (63.54 ± 26.59 sec vs. 37.11 ± 4.68 sec), whereas the GF treatment group showed a higher swimming time than the Control group. In particular, the GF200 group was significantly higher than the Control group (37.11 ± 4.68 sec vs. 60.82 ± 26.71 sec, P < .05).
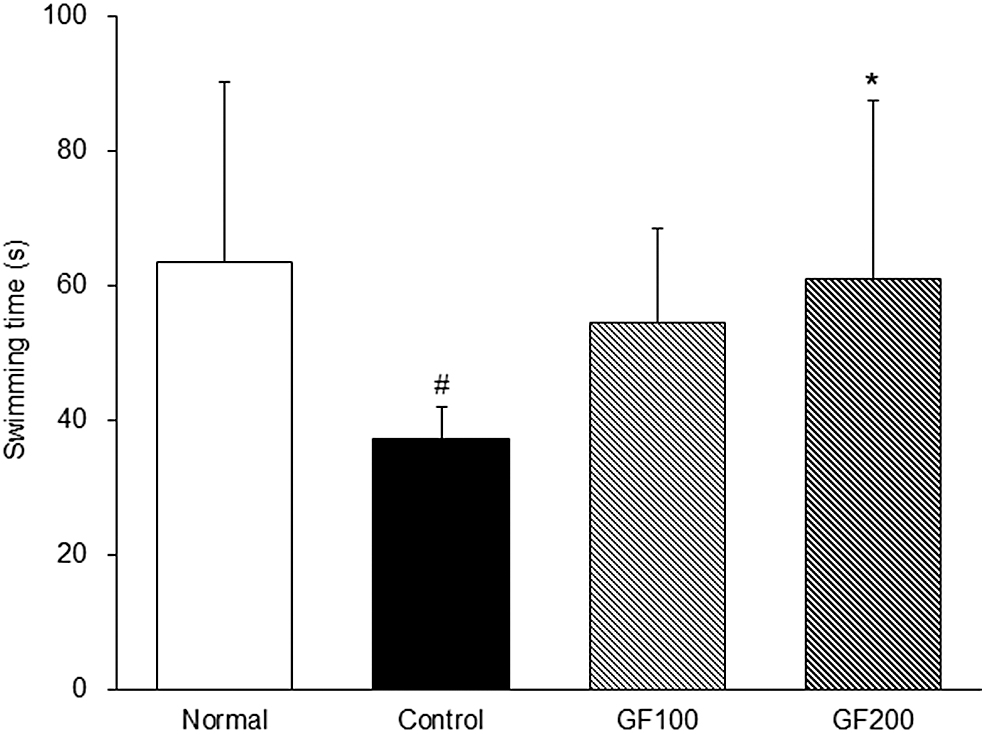
Improvement of swimming time by GF on DEX-induced MA. Normal, normal group; Control, MA-induced mice were treated with distilled water; GF100, MA-induced mice were treated with GF hot water extract 100 mg/kg body weight; GF200, MA-induced mice were treated with GF hot water extract 200 mg/kg body. Data are mean ± SD (n = 8). Significance: # P < .05 versus the Normal group, *P < .05 versus the Control group.
Measurement of GOT and GPT levels
To investigate the effect of GF on liver function, the GOT and GPT were measured, and the results are shown in Figure 4. The Control group showed significantly higher GOT and GPT values than the Normal group (GOT; 9.81 ± 4.61 IU/L vs. 35.88 ± 2.78 IU/L, GPT; 3.14 ± 1.17 IU/L vs. 18.88 ± 1.32 IU/L), whereas the GF-treated group showed lower values than the Control group. In particular, the GOT and GPT levels after GF200 treatment were lower than GF100 treatment.
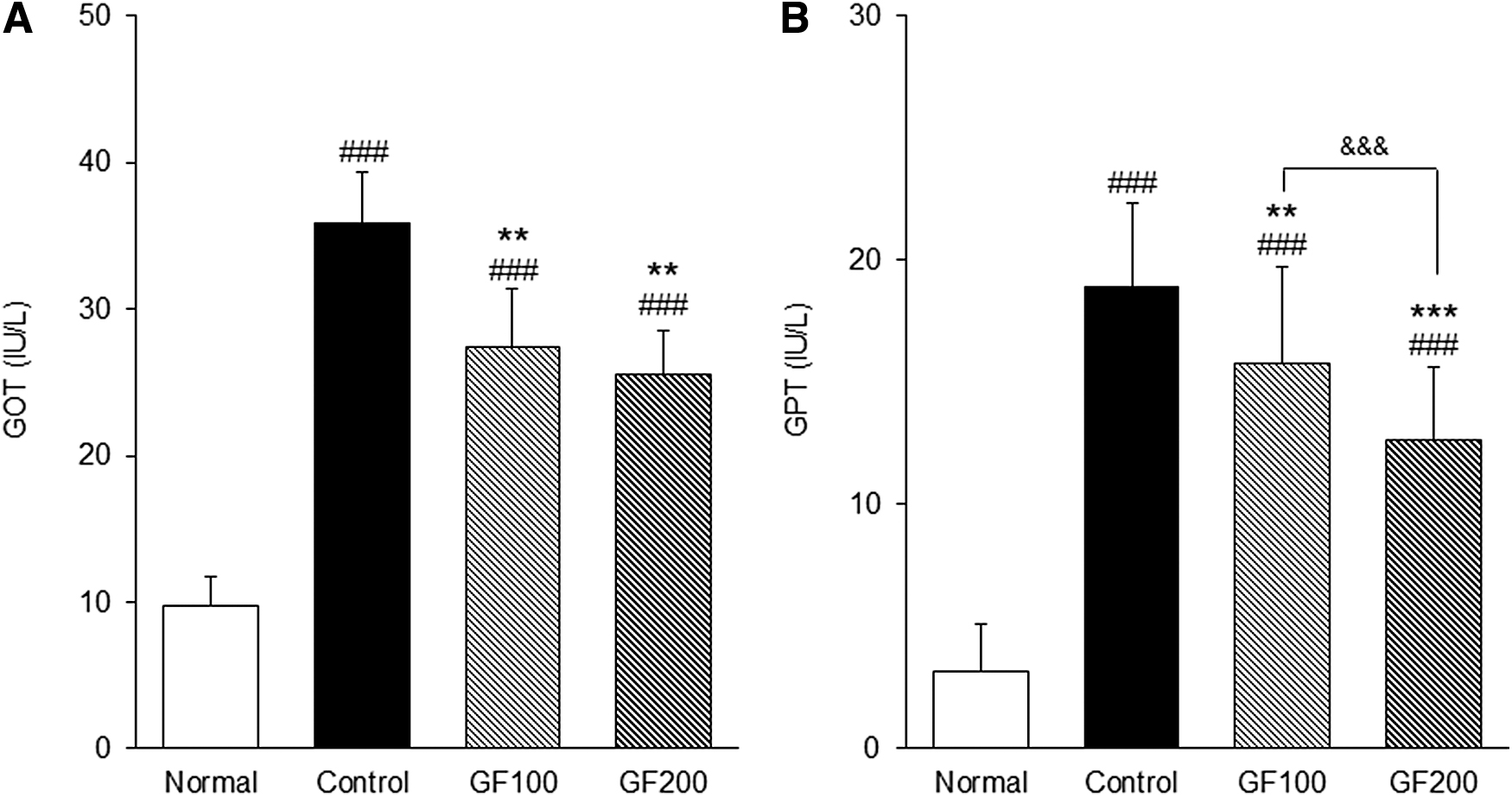
Effect of GF on levels of liver function.
Expression of NOX in DEX-induced MA
The change of NOX, OS-generating related factors, was examined using Western blotting. The expression of NOX in the DEX-induced MA group was significantly higher than the Normal group (NOX2; 1.21-fold, p47phox; 1.17-fold). Conversely, the level of NAPDH oxidase was significantly lower in the GF-treated group than in the Control group (P < .01 or P < .001). In this study, the GF-treated group showed similar expression levels to the Normal group (Fig. 5).
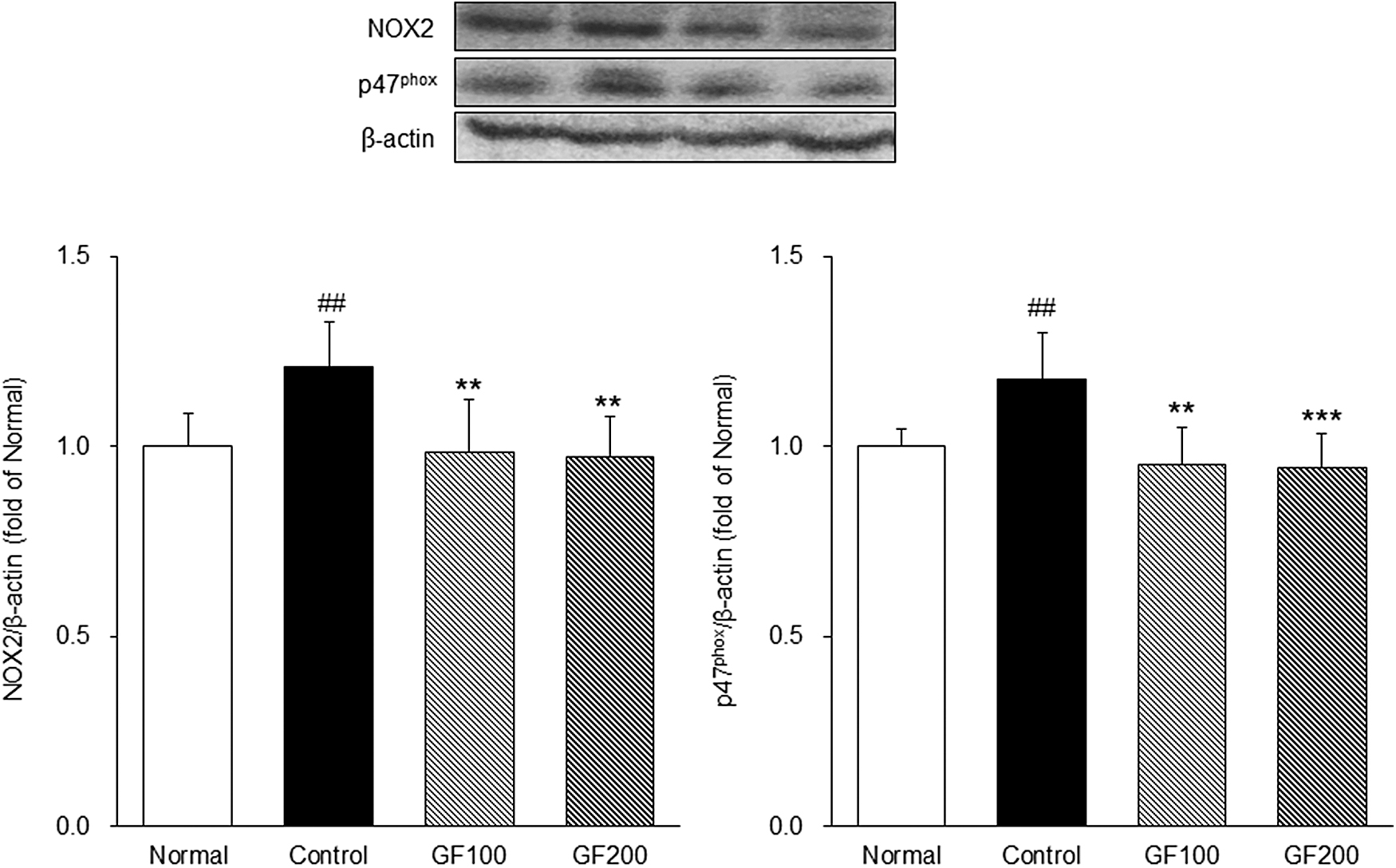
Effects of GF on NOXs. The expressions of NOXs were measured by Western blotting. Normal, normal group; Control, MA-induced mice were treated with distilled water; GF100, MA-induced mice were treated with GF hot water extract 100 mg/kg body weight; GF200, MA-induced mice were treated with GF hot water extract 200 mg/kg body. Data are mean ± SD (n = 8). Significance: ## P < .01 versus the Normal group, **P < .01, ***P < .001 versus the Control group.
Expression of the muscle protein synthesis pathway in DEX-induced MA
IGF-1 is involved in muscle growth as a potent anabolic factor and is known to activate the PI3K/Akt/mTOR pathway. 18 Also in skeletal muscle, the mTOR signaling pathway is recognized as a major pathway regulating protein synthesis, activating and inhibiting S6K to regulate protein synthesis. 19,20 As shown in Figure 6, the expression of factors including IGF-1, p-PI3K, p-Akt, and p-mTOR of the DEX-induced control group was significantly lower than the Normal group, and p-S6K was also significantly lower (IGF-1; 0.72-fold, p-PI3K; 0.64-fold, p-Akt; 0.62-fold, p-mTOR; 0.72-fold, p-S6K; 0.76-fold). Conversely, the expression of these proteins was significantly higher in the GF-treated group than in the Control group (P < .05, P < .01, and P < .001). Therefore, GF improved the anabolic function in DEX-treated mice through the PI3K/Akt/mTOR pathway by regulating IGF-1.
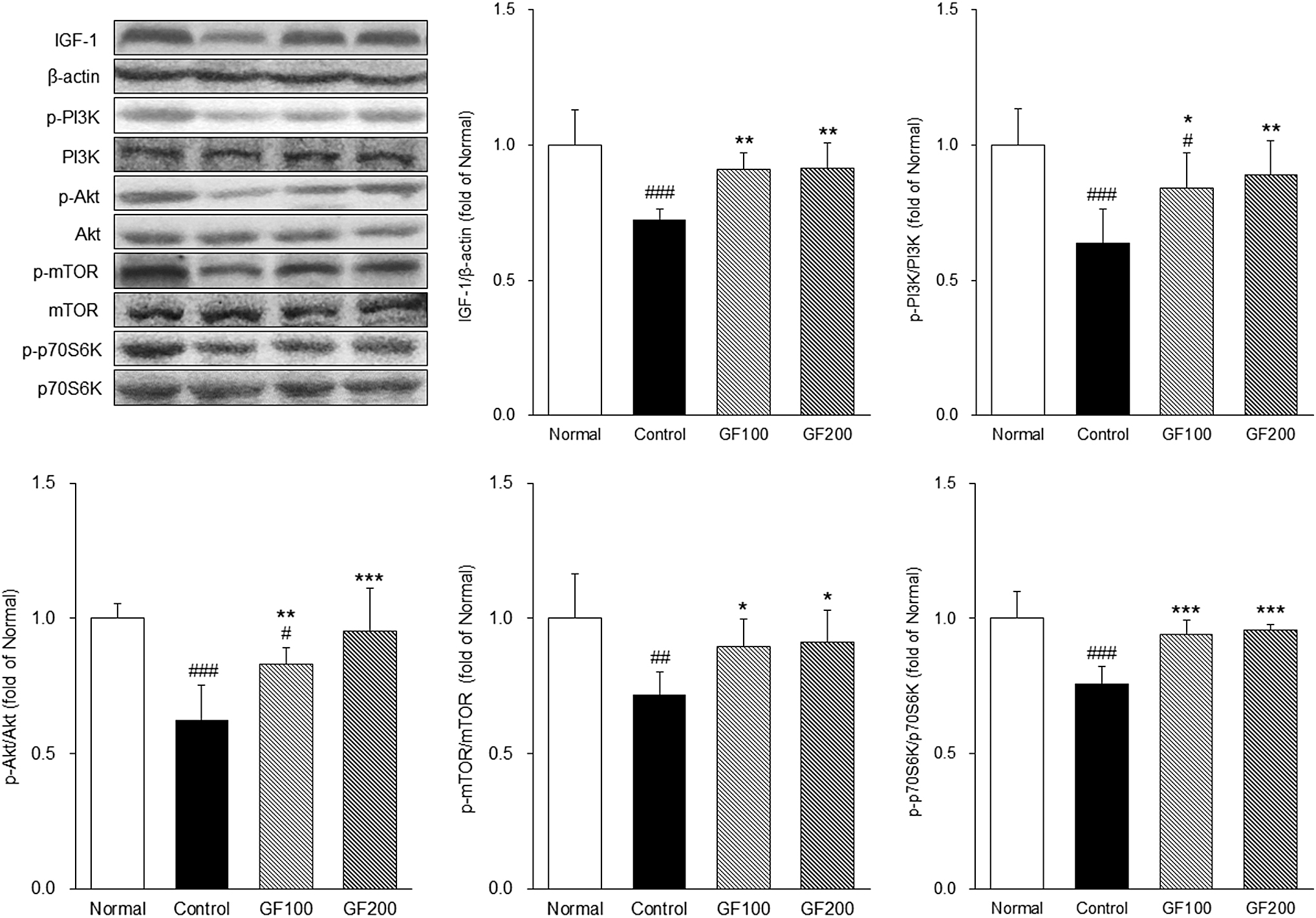
Effects of GF on the muscle protein synthesis pathway. The expressions the muscle anabolic-related proteins were measured by Western blotting. Normal, normal group; Control, MA-induced mice were treated with distilled water; GF100, MA-induced mice were treated with GF hot water extract 100 mg/kg body weight; GF200, MA-induced mice were treated with GF hot water extract 200 mg/kg body. Data are mean ± SD (n = 8). Significance: # P < .05, ## P < .01, ### P < .001 versus the Normal group, *P < .05, **P < .01, ***P < .001 versus the Control group.
Expression of the muscle protein catabolic pathway in DEX-induced MA
SIRT1 is associated with activation of AMPK and increases the transcriptional activity of FOXO3a. In this study, activation of AMPK was found to block muscle growth. 21 We investigated the molecular mechanism of GF on MA by evaluating the levels of proteins involved in the muscle degradation pathway. As shown in Figure 7, factors including SIRT1, PGC-1α, and p-FOXO3a in the DEX-induced control group were significantly lower than the Normal group (SIRT1; 0.82-fold, PGC-1α; 0.68-fold, p-FOXO3a; 0.69-fold), and factors including p-AMPK, FOXO3a, MuRF-1, and MAFbx were significantly higher than the Normal group (p-AMPK; 1.38-fold, FOXO3a; 1.23-fold, MuRF-1; 1.21-fold, MAFbx; 1.28-fold).

Effects of GF on the muscle protein catabolic pathway. The expressions the muscle catabolic-related proteins were measured by Western blotting. Normal, normal group; Control, MA-induced mice were treated with distilled water; GF100, MA-induced mice were treated with GF hot water extract 100 mg/kg body weight; GF200, MA-induced mice were treated with GF hot water extract 200 mg/kg body. Data are mean ± SD (n = 8). Significance: # P < .05, ## P < .01, ### P < .001 versus the Normal group, *P < .05, **P < .01, ***P < .001 versus the Control group, && P < .01, &&& P < .001 versus the GF group.
In this study, the expression of SIRT1, AMPK, PGC-1α, and FOXO3a (P < .05, P < .01, and P < .001) and the expression of MuRF-1 and MAFbx regulated by FOXO3a transcription (P < .05 and P < .001) were also significantly higher than the Control group. In particular, in the GF200 group, the following factors were found to be similar to those of the Normal group.
DISCUSSION
MA is a disease that occurs when protein exceeds synthesis, and it is a common complication of various diseases such as aging, chronic diseases, and cancer. 1 We conducted an experiment to find an effective herbal medicine for preventing MA. In the present study, we confirmed the preventive effect of GF on muscle dysfunction using a DEX-induced MA mouse model.
First, ultra performance liquid chromatography (UPLC) chromatogram was performed to confirm the geniposide content of GF used in this experiment. Many studies have reported that geniposide has a positive effect on skeletal muscle. 14,15 The UPLC analysis revealed that the GF used in this study contained 176.625 ± 0.809 mg/g of geniposide. These results suggested that GF may be effective for MA improvement through physiologically active ingredients. Also, we confirmed the levels of GOT and GPT in the serum. Many studies have reported that GOT and GPT levels are high in patients with MA, and that the increase in GOT and GPT levels is associated with an increase in the prevalence of MA. 22 In this study, it was confirmed that the GOT and GPT levels were lower in the GF-treated group than in the MA-induced control group.
In recent studies, OS has been shown to be a major cause of MA. OS is known to be a key regulator of cellular signaling pathways and is associated with muscle protein degradation and myonuclear apoptosis. 23 –25 OS is well known as the root cause of several diseases, and an increase in OS in the human body adversely affects mitochondrial dysfunction, DNA, cell proliferation, and progression of age-related disorders. 26 –28 Since overactivation of NOX is known to be a major source of ROS, various studies targeting NOX are being conducted. 28,29 NOX is composed of transmembrane proteins and cytosolic proteins. NOX activation is initiated by phosphorylation of the cytoplasmic protein p47phox, leading to the activation of transmembrane proteins such as NOX2 and several cytosolic proteins. 30
In this study, it was confirmed that the expression of NAPDH oxidase was higher in the Control group induced by DEX than in the Normal group, and the expression of NOX was significantly higher in the GF-treated group than in the Control group. These results suggest that GF affects OS and can have a positive effect on muscular atrophy by effectively reducing OS.
It has been reported that skeletal muscle hypertrophy is associated with an increase in IGF-1, and the addition of IGF-1 to muscle cells promotes myotube hypertrophy. 31 In addition, IGF-1 induces activation of the PI3K/Akt pathway, and PI3K/Akt activated by IGF-1 not only plays an important role in muscle synthesis but also prevents muscle protein atrophy. 32 Activation of the PI3K/Akt pathway phosphorylates and activates mTOR kinase, and it is known that activated mTOR kinase increases polysynaptic synthesis through phosphorylation and activation of p70S6 kinase (p70S6K). 33 According to the study of Lee et al., increasing p-Akt/Akt and p-p70S6K/p70S6K attenuated skeletal MA. 34 In addition, Cui et al. reported that IGF-1/PI3K/Akt signaling not only promotes muscle hypertrophy but also inhibits atrophy, and can alleviate MA through IGF-1/PI3K/Akt signaling. 35 These findings indicate that the IGF-1/PI3K/Akt pathway inhibits MA by promoting skeletal muscle hypertrophy by aiding muscle synthesis.
As in the previously published results, in this study, the expressions of IGF-1, p-PI3K, p-Akt, p-mTOR, and p-p70S6K were significantly higher in the GF-treated group than in the Control group. These results indicate that GF activates PI3K/Akt signaling by regulating IGF-1 in the sarcopenia model to relieve MA.
To effectively inhibit MA, it is considered important to regulate not only protein synthesis but also mitochondrial biosynthesis pathways and protein degradation essential for cell activity. AMPK is a key factor that limits muscle size and hypertrophy, and is known to promote MA through autophagy and proteolysis. 21 Also, the mitochondrial dysfunction due to MA results in insufficient energy supply and activation of AMPK, which promotes the expression of MuRF-1 and MAFbx/Atrogin-1, in turn accelerating muscle breakdown. 36,37 The negative effects of AMPK on muscle growth have been studied for a long time. The elevation of AMPK promotes the activation of FOXO3a along with inhibition of the IGF-1/PI3K/Akt pathway.
FOXO3a is a transcription factor involved in protein degradation, and activation of FOXO3a activates the expression of MuRF-1 and MAFbx/Atrogin-1. MuRF-1 and MAFbx/Atrogin-1 activated by AMPK and FOXO3a were shown to increase the conditions associated with various diseases related to MA, such as inflammation, aging, and chronic diseases. 7 In the study of Sandri et al., it was found that FOXO induces MAFbx/Atrogin-1 to induce skeletal MA, and various other studies have revealed that the FOXO pathway is related to MA. 10,38,39 As a result of this study, as in previous studies, the AMPK/SIRT1/PGC-1α and FOXO pathways were regulated in the GF group, and in particular, these pathways were well regulated in the GF200 group. In addition, the expression of MuRF-1 and MAFbx/Atrogin-1 was reduced by regulating this pathway. Because muscle proteins are constantly undergoing replacement, both the catabolic and anabolic pathways must be well regulated.
These results suggest that GF suppresses MA through regulation of the muscle protein catabolic pathway.
Consequently, in the present study, GF treatment significantly reduced the expression of OS-related factors. Also, the IGF-1/PI3K/Akt and AMPK/SIRT1/PGC-1α-FOXO pathways were significantly regulated. In conclusion, GF regulates muscle protein synthesis and catabolic pathways, and in particular, it is judged to improve MA by regulating the proteolytic pathway, the AMPK/SIRT1/PGC-1α-FOXO pathway. These results help to understand the mechanism of GF protection of muscle mass.
Footnotes
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This work was supported by the National Research Foundation of Korea (NRF) (No. 2018R1A5A2025272) and the Ministry of Agriculture, Food and Rural Affairs (MAFRA) (No. 121014031HDD20).