
Abstract
Alcoholic liver disease (ALD) is a major chronic liver disease. Chronic alcohol consumption induces dysbiosis, disruption of gut barrier function, oxidative stress, inflammation, and changes in lipid metabolism, thereby leading to ALD. In this study, we investigated whether the commercial Morinda citrifolia extract Nonitri can ameliorate ALD symptoms through the gut–liver axis. We used mice chronically administered EtOH and found a marked increase in serum endotoxin levels and biomarkers of liver pathology. Moreover, the EtOH-treated group showed significantly altered gut microbial composition particularly that of Alistipes, Bacteroides, and Muribaculum and disrupted gut barrier function. However, Nonitri improved serum parameters, restored the microbial proportions, and regulated levels of zonula occludens1, occludin, and claudin1. Furthermore, Nonitri suppressed inflammation by inhibiting endotoxin-triggered toll-like receptor 4-signaling pathway and fat deposition by reducing lipogenesis through activating AMP-activated protein kinase in the liver. Furthermore, Pearson's correlation analysis showed that gut microbiota and ALD-related markers were correlated, and Nonitri regulated these bacteria. Taken together, our results indicate that the hepatoprotective effect of Nonitri reduces endotoxin levels by improving gut health, and inhibits fat deposition by regulating lipid metabolism.
INTRODUCTION
Chronic alcohol abuse is a major risk factor for tissue injuries. 1 Alcohol induces liver, gut, and brain damage, thereby causing various metabolic disorders. 2 Alcoholic liver disease (ALD) is one of the most common chronic liver diseases caused by excessive alcohol consumption over a long period of time. ALD progressively develops in stages of asymptomatic steatosis, steatohepatitis, fibrosis, cirrhosis, and hepatocellular carcinoma. 3 Alcohol not only induces inflammation but also inhibits AMP-activated protein kinase (AMPK) activation, which upregulates the expression of lipogenic genes, such as sterol regulatory element-binding protein 1 (SREBP1) and carbohydrate-responsive element-binding protein (ChREBP), and downregulates the expression of fatty acid oxidation-related genes such as peroxisome proliferator-activated receptor (PPAR)-α to cause ALD. 4,5
Several studies have reported that alcohol consumption-induced ALD directly inflicts damage to the liver; however, recent studies suggest that alcohol can induce ALD by altering the functions of the gut–liver axis. 6 Chronic consumption of alcohol changes the gut microbiota composition and increases the production of toxic substances such as lipopolysaccharide (LPS), causing inflammation in the gut and disrupting tight junctions to induce a leaky gut. 7 As a result, increased blood endotoxins enter the liver and induce ALD.
Morinda citrifolia of the family Rubiaceae is a fruit-bearing tree. In Southeast Asia, it is widely known as “noni.” 8 M. citrifolia contains active compounds such as flavonoids, tannins, anthraquinones, coumarins, and iridoids. 9 Moreover, this plant has been traditionally used for therapy because of its antibacterial, antiviral, antitumor, and anti-inflammatory properties. 10 Reportedly, M. citrifolia helps alleviate obesity and inhibits obesity-related metabolic disorders. 11 Guo et al reported that noni fruit and fermented noni juice reduce acute alcohol-induced liver injury. 12 Moreover, polysaccharides extracted from noni showed protective effects against inflammatory bowel disease. 13 However, the effect of M. citrifolia on gut health and alcoholic liver damage by regulating the gut microbiota has not yet been reported.
This study investigated the preventive effects of the commercialized M. citrifolia extract Nonitri® on mice with ALD and its effects on regulating intestinal microflora and modulating intestinal barrier integrity. Our results suggest that Nonitri mitigates ALD by maintaining the intestinal barrier and the eubiotic state of the gut environment, both of which play indispensable roles in the pathogenesis of ALD.
MATERIALS AND METHODS
Materials
Fluorescein isothiocyanate-dextran (FITC-dextran; 4 kDa), asperulosidic acid (AA), deacetylasperulosidic acid (DAA), scopoletin, acetonitrile (ACN), and formic acid (FA) were purchased from Sigma-Aldrich (St. Louis, MO, USA). Acetyl-CoA carboxylase (ACC), carnitine palmitoyltransferase 1 (CPT1), fatty acid synthase (FAS), tumor necrosis factor-α (TNF-α), interukin-6 (IL6), PPAR-α, SREBP1, and toll-like receptor 4 (TLR4) were obtained from Abcam (Cambridge, MA, USA), and AMPK, phospho-AMPK, ChREBP, stearoyl-CoA desaturase-1 (SCD1), and β-actin were purchased from Cell Signaling Technology (Beverly, MA, USA). Secondary anti-rabbit antibodies were purchased from Thermo Fisher Scientific (Waltham, MA, USA). Liquid chromatography–mass spectrometry (LC-MS)-grade water, ethanol (EtOH), and acetonitrile were obtained from JT Baker (Phillipsburg, NY, USA).
Preparation of Nonitri
Nonitri was provided by COSMAX NS Inc., (Seongnam, Korea). Dried M. citrifolia fruits were pulverized for extraction. Powdered M. citrifolia fruit was extracted with 10% EtOH at 80°C for 4 h, filtered, concentrated, mixed with 50% w/w of dextrin, and then freeze dried.
High-performance liquid chromatography–electrospray ionization mass spectrometry
The high-performance liquid chromatography (HPLC) system was equipped with a Waters triple quadrupole mass spectrometer (Xevo TQ-MS, Waters Corp., Milford, MA, USA) with an electrospray ionization (ESI) source. The samples were analyzed on an ACQUITY UPLC BEH C18 VanGuard precolumn (1.8 μm, 2.1 × 5 mm) and an ACQUITY UPLC BEH C18 column (1.8 μm, 2.1 × 100 mm). The flow rate and column temperature were set at 0.2 mL/min and 40°C, respectively. Mobile phase A and B were 0.1% FA/water and 0.1% FA/ACN, respectively. A gradient condition of the two solvents was as follows: 0.0 − 1.0 min, 0% B; 1.0 − 16.0 min, linear increase from 0% B to 90% B; 16.0 − 17.0 min, 90% B; 17.0 − 17.5 min, linear decrease from 90% B to 0% B; 17.5 − 22.0 min, 0% B. For the ionization of the three constituents, capillary voltage, source temperature, desolvation temperature, and desolvation gas flow were set as 4 kV, 150°C, 500°C, and 1000 L/h, respectively.
Fragmentation parameters (Supplementary Table S1) of each constituent were automatically set using Waters' IntelliStart software. For the HPLC-ESI-MS experiments, 8 mg of Nonitri was carefully weighed and dissolved in 2 mL of 50% EtOH, and the solution was sonicated for 2 h. After sonication, the sample solution was filtered using a 0.22-μm PTFE filter and diluted with water to the appropriate concentration for the analysis.
Animals
Male C57BL/6J mice (8 weeks old) were purchased from Central Lab Animal Inc., (Seoul, Republic of Korea) and housed in a specific pathogen-free animal facility (12-h light/12-h dark cycle, 22 ± 2°C). Forty-five mice were randomly allocated into five groups and received diets with ad libitum food and energy intake. The animal experiments were approved by the Animal Welfare Committee of the Korea Food Research Institute (Approval No. KFRI-M-20016). All the mice were fed AIN-93 diet. Normal control (NC) mice were orally administered distilled water, and the alcohol-administered (EtOH) mice were orally gavaged with 40% EtOH (4.0 g/kg/day). The positive control (PC) group and Nonitri-administered groups (Nonitri-50 and Nonitri-100) were administered the same dose of alcohol as the EtOH group but concomitantly orally administered a daily dose of 50 mg/kg of Hovenia dulcis fruit extract as PC and 50 or 100 mg/kg of Nonitri, respectively. H. dulcis fruit extract and Nonitri were provided by Cosmax NS.
The concentration of EtOH was gradually increased during the first week to 40% and was maintained until the end of the experiment. Mice were given alcohol and H. dulcis fruit extract or Nonitri for 9 weeks. Body weight and food intake were measured weekly until the end of the experiment. At the end of week 9, all animals were euthanized and liver and colon were collected for further analysis.
Biochemical analysis
At the completion of the study, mice were made to fast for 12 h and euthanized using isoflurane inhalation. Blood was collected and serum was separated using centrifugation. Serum endotoxin levels were estimated using the Chromogenic Endotoxin Quant Kit (Thermo Fisher Scientific). Serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels were measured using kits from Cusabio (Wuhan, China), and serum triglyceride (TG), lactate dehydrogenase (LDH), and reduced glutathione (GSH) levels were determined using kits from Abcam. All assays were performed according to manufacturers' instructions.
Intestinal permeability analysis
FITC-dextran (500 mg/kg) was orally administered to 6 h fasted mice. Blood was drawn from the tail vein after 2 and 5 h of administration. The plasma was separated using centrifugation (3000 g for 10 min) and fluorescence was immediately measured using a microplate reader (Molecular Devices, Sunnyvale, CA, USA) at 485/535 nm (excitation/emission). Plasma levels of FITC-dextran were calculated using standard curves generated by the dilution of FITC-dextran in untreated plasma.
Gut microbial composition
At week 8, fresh fecal samples were collected and immediately stored at −80°C for analysis of gut microbial composition. Fecal DNA was extracted, and 16S rRNA gene amplification was performed on a MiSeq platform (Illumina, San Diego, CA, USA) at Macrogen (Seoul, Korea) according to the manufacturer's instructions. Microbial diversity and taxonomic composition analyses were performed using QIIME software. Correlation analysis was performed to evaluate the interactions between gut microbiota and parameters associated with lipid metabolism based on Pearson's correlation coefficient using R software (version 4.1.0).
Reverse transcription–quantitative polymerase chain reaction
Total RNA was extracted from the colon using the TaKaRa MiniBEST Universal RNA Extraction Kit (TaKaRa, Otsu, Japan) and reverse transcribed to cDNA using the iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA, USA). cDNA was amplified on a QuantStudio™3 Real-Time PCR System (Thermo Fisher Scientific) using the PowerUp™ SYBR™ Green Master Mix (Applied Biosystems, Foster City, CA, USA). The sequences of primers are as follows: claudin1: forward, 5′- GGGGACAACATCGTGACCG -3′ and reverse, 5′- AGGAGTCGAAGACTTTGCACT -3′; occludin: forward, 5′- TTGAAAGTCCACCTCCTTACAGA -3′ and reverse, 5′- CCGGATAAAAAGAGTACGCTGG -3′; zonula occludin-1 (ZO1): forward, 5′- GCCGCTAAGAGCACAGCAA -3′ and reverse, 5′- TCCCCACTCTGAAAATGAGGA -3′; and GAPDH: forward, 5′- AGGTCGGTGTGAACGGATTTG -3′ and reverse, 5′- TGTAGACCATGTAGTTGAGGTCA -3′. Target gene mRNA expression was normalized that to GAPDH and results were calculated using the 2−ΔΔCT method.
Western blotting
Total proteins of mouse livers from each group were extracted using PRO-PREP™ (iNtRON Biotechnology, Seongnam, Republic of Korea) containing phosphatase inhibitors and separated by 4–20% gradient sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel, followed by being transferred to polyvinylidene difluoride (PVDF) membrane. Then, the PVDF membrane was blocked in 5% skimmed milk and incubated with primary antibodies at 4°C overnight. The blots were then incubated with secondary antibodies, and bands were detected using the ChemiDoc XRS+ imaging system (Bio-Rad).
Histological analysis
Oil Red O (ORO) staining was performed to evaluate lipid accumulation in the liver. The tissues were embedded in Tissue-Tek OCT compound (Sakura Finetek, CA, USA) and cut into 10-μm sections using a cryostat (Leica CM1850, Wetzlar, Germany). The liver cryosections were stained with ORO. The slides were scanned using a PANNORAMIC 250 Flash III slide scanner (3DHistech, Ltd., Budapest, Hungary), and scanned images were acquired using the CaseViewer software (3DHistech, Ltd).
Statistical analysis
All experimental data are presented as the mean ± standard errors of the means. Statistical analysis was performed using one-way analysis of variance followed by Tukey's analysis using Prism 9.0 software (GraphPad, San Diego, CA, USA). Statistical significance was set at P < .05.
RESULTS
Phytochemical content of Nonitri
In the ESI-MS experiment, monosodiated ions of two iridoid compounds, AA and DAA, were selected as precursor ions, and monoprotonated scopoletin ions were selected as precursor ions (Supplementary Table S1). The iridoid precursor ions generated product ions as described in Supplementary Table S1 by loss of a monosaccharide (180 Da), and the scopoletin precursor ions generated product ions, as described in Supplementary Table S1, by the loss of C2H4O2 (60 Da). Further HPLC-ESI-MS experiments using standard and extract solutions showed that the retention times of AA, DAA, and scopoletin were 5.14, 3.88, and 7.01 min regardless of the presence of Nonitri (Fig. 1). The estimated concentrations of AA, DAA, and scopoletin in Nonitri obtained from standard addition curves were 1.91 ± 0.24, 14.07 ± 1.42, and 0.18 ± 0.01 μg/mg materials, respectively.

HPLC-ESI-MS/MS chromatograms of
Changes in body weight and biochemical parameters
Our investigations revealed that all EtOH-treated mice showed significantly lower body weights than the NC group without changes in food intake (Fig. 2A, B). Endotoxin levels were also markedly upregulated in the EtOH group (Fig. 2C). EtOH-treated mice showed significantly increased serum levels of hepatic enzymes, such as ALT, AST, and LDH, and the serum TG levels (Fig. 2D–G). Moreover, EtOH consumption decreased antioxidant enzyme GSH (Fig. 2H). Although Nonitri supplementation did not improve alcohol-induced reduction in body weights, it significantly reduced endotoxemia. In addition, the blood biomarkers of metabolic disorders were markedly reversed by Nonitri.
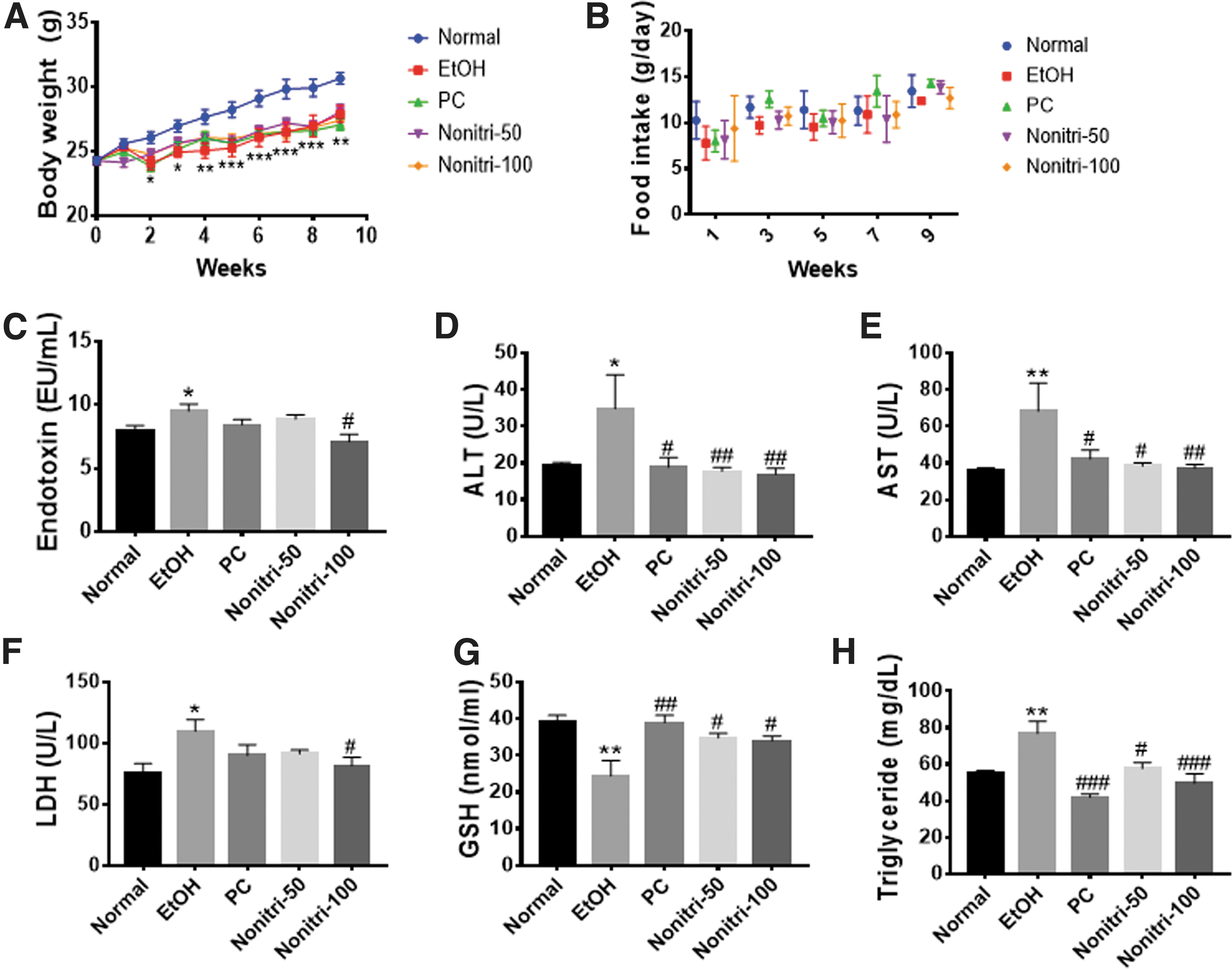
Effects of Nonitri treatment of mice with chronic alcohol consumption on body weight and blood markers.
Effects of Nonitri on intestinal microbial composition and intestinal barrier function
At the genus level, EtOH-treated mice showed significantly different gut microbial compositions. Although no significant difference was found, the EtOH group had a higher abundance of Bacteroides and Alistipes and a lower abundance of Muribaculum than the NC group (Fig. 3A, B). Moreover, our observations confirmed that gut permeability increased in the EtOH-treated group (Fig. 3C). EtOH-treated mice did not show decreased occludin gene expression but exhibited significantly reduced gene expressions of ZO1 and claudin1 (Fig. 3D). However, Nonitri regulated the proportions of Bacteroides, Alistipes, and Muribaculum and improved gut permeability by upregulating ZO1, occludin, and claudin1 gene expression.

Nonitri changes the microbial composition and ameliorates gut barrier function in mice after chronic alcohol consumption.
Effects of Nonitri on inflammation in the liver
The effects of Nonitri on hepatic inflammation were determined by western blotting of proteins obtained from the liver of the mice from different groups. EtOH-treated mice showed significantly increased TLR4, IL6, and TNF-α protein expression (Fig. 4). However, Nonitri supplementation reduced TLR4 and inflammatory cytokine expressions.
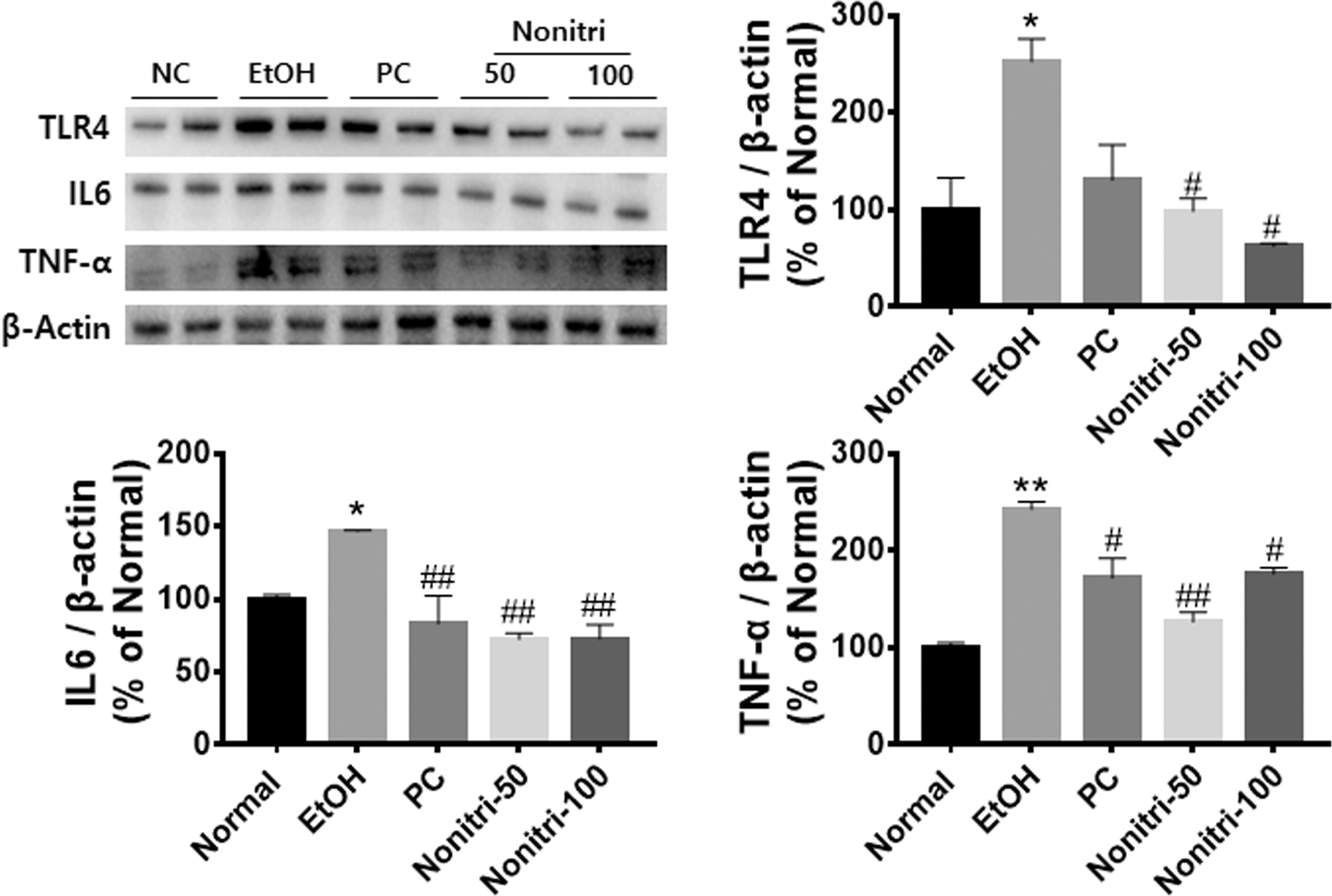
Nonitri ameliorates alcohol-induced inflammation in the liver. Representative western blots of TLR4, IL-6, and TNF-α are shown. β-Actin was used as an internal control. Blots are normalized to β-actin and values are presented as the mean percentages of NC ± SEM. *P < .05, and **P < .01 versus NC; # P < .05, and ## P < .01 versus EtOH. IL, interleukin; TLR4, toll-like receptor 4 TNF-α, tumor necrosis factor-α.
Effects of Nonitri on lipid metabolism in the liver
We found that in alcohol-fed mice, the expression of phosphorylated AMPK was downregulated (Fig. 5A). Moreover, these mice showed significantly increased expressions of lipogenesis-regulating proteins, SREBP1 and ChREBP. Overexpression of these two proteins caused upregulation of downstream lipogenic enzymes, such as SCD1, ACC, and FAS. In contrast, reduced AMPK phosphorylation led to decreased expression of β-oxidation-related proteins, such as PPAR-α and CPT1 in alcohol-fed mice. Nonitri supplementation further facilitated the EtOH-induced reduction of AMPK phosphorylation. In addition, Nonitri downregulated lipogenic protein expressions and upregulated lipid oxidation protein expressions. ORO staining showed that EtOH consumption significantly (P < .05) increased lipid accumulation, whereas Nonitri-induced changes in lipid metabolism inhibited alcohol-induced liver fat deposition (Fig. 5B).

Nonitri ameliorates alcohol-induced changes of lipid metabolism in liver.
Correlation of gut microbiota and metabolic parameters
To determine the interactions between gut microbiota and serum and liver parameters in ALD, we performed a heat map analysis, showing color-coded Pearson correlations between the most abundant gut bacterial taxa at the genus level and ALD-related parameters (Fig. 6). It is worth noting that Bifidobacterium, Muribaculum, and Lactobacillus were negatively correlated with the pathophysiological parameters of ALD but were positively associated with the parameters of lipid oxidation. Furthermore, Kineothrix, Bacteroides, Mucispirillum, Alistipes, Turicibacter, and Enterocloster showed positive correlations with the pathophysiological parameters of ALD.
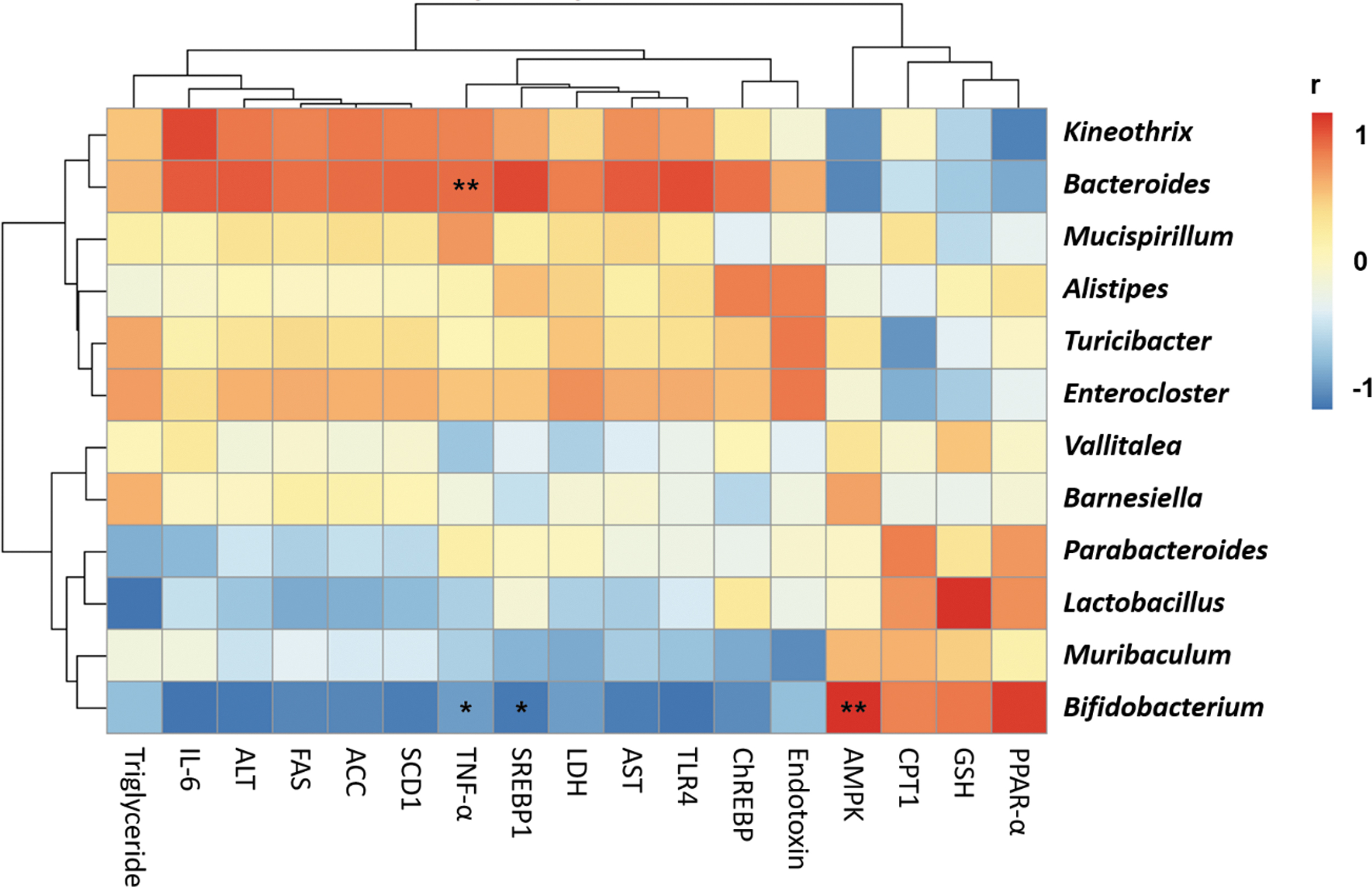
Alcohol-induced gut microbiota changes are closely associated with the features of liver pathology. Correlation analyses between gut microbiota and biochemical parameters in alcohol-induced liver disease affected by Nonitri. The color scale represents the Pearson R value, with red and blue specified positive and negative correlation, respectively (*P < .05; **P < .01).
DISCUSSION
Alcohol-induced chronic liver damage directly affects the liver 14 and/or indirectly affects the liver through the gut–liver axis. 2 M. citrifolia contains several iridoid compounds, such as AA and DAA, and the coumarin compound scopoletin. 15 These compounds exhibit a wide range of biological functions, including anti-inflammatory, antioxidant, and anticancer effects. 16 Therefore, we expected these compounds to be the active compounds of the commercial product Nonitri. In this study, we determined whether Nonitri could improve alcoholic liver damage by improving gut health. Endotoxin from the gut is an essential factor in the pathogenesis of ALD 17 and its increase causes a significant increase in serum markers of liver injury, such as ALT, AST, and LDH. 18 Moreover, changes in the hepatic enzymes are strongly associated with metabolic disorders and correlate with serum TG levels. 19
In this study, we found that EtOH consumption significantly increased endotoxin, serum hepatic enzymes, and TG levels, suggesting that alcohol consumption changed gut microbiota and induced inflammation and fat deposition in the liver. Alcohol consumption causes changes in the intestinal microbiota and increases intestinal permeability, thereby releasing endotoxins such as LPS. 20
Recent research has identified alcohol-induced gut dysbiosis, due to imbalances in the gut microbiota, as one of the primary causes of ALD. Microbial dysbiosis associated with increases in Proteobacteria and decrease in the abundance of Bacteroidetes in patients with ALD lead to increased permeability of the gut epithelium and decreases in mucin production. 21 It was also observed that Lachnospiraceae and Ruminococcaceae were decreased in patients with alcohol-related cirrhosis. 22 In addition, alcohol intake in mice was associated with a decrease in the abundance of Firmicutes such as Lactobacillus and Lactococcus. 23 Gut barrier disruption leads to the penetration of endotoxin, a component of the Gram-negative bacterial cell wall, into the blood stream in alcoholic patients and animal models. 24,25 It has been demonstrated that increased blood endotoxin concentrations correlate with increased gut permeability. 26 Therefore, targeting the microbiome could be an attractive strategy for treating ALD.
In this study, we found that alcohol consumption induced changes in gut microbial composition by altering the levels of Alistipes, Bacteroides, and Muribaculum. The increases in the abundance of Alistipes and Bacteroides is correlated with changes in blood ALT and TG, 27,28 which induce ALD. 29 Bacteroides is an endotoxin-producing bacterium that may increase intestinal permeability. 30 Muribaculum is a beneficial gut bacterium and a dominant genus in the mouse gut. 31 Moreover, alcohol consumption substantially reduces the abundance of these bacteria. 32 Therefore, we believe that the changes in these bacteria lead to the disruption of gut barrier function by increasing the levels of toxic substances in the gut. ZO1, claudin1, and occludin, which are tight junction proteins, are closely associated with gut permeability. Downregulated expression of the genes encoding these proteins results in a leaky gut, thereby causing endotoxemia and metabolic disorders in various tissues of the body. 33
Therefore, decreased expression of ZO1 and claudin1 in alcohol-fed mice causes an increase in gut permeability. In this study, 100 mg/kg of Nonitri improved the proportion of Alistipes, Bacteroides, and Muribaculum, thereby reducing the generation of toxic substances in the gut. In addition, Nonitri enhanced the gut barrier function by increasing the expression of ZO1, occludin, and claudin1. These results suggest that Nonitri improves gut barrier function by modulating the gut microbiota.
In the liver, TLR4 is activated by toxic substrates from the gut, 34 resulting in increased expression of inflammatory cytokines, such as TNF-α and IL6. 35 These cytokines can induce hepatosteatosis and liver injury through the modulation of SREBP1. 36 –38 In this study, TLR4, TNF-α, and IL6 levels were significantly increased following EtOH treatment. However, Nonitri significantly reduced the expression of TLR4 and also decreased the expression of the inflammatory cytokines, suggesting that the hepatic inflammatory response was reduced by altering the gut–liver axis, and this reduction of inflammatory cytokines is thought to have some effect on lipid metabolism.
AMPK is a cellular energy homeostasis enzyme that regulates fatty acid oxidation and lipogenesis. 39 Alcohol consumption inhibits the phosphorylation of AMPK, which plays a vital role in developing hepatic steatosis in alcohol-induced liver damage. 40 ChREBP and SREBP are downstream proteins of AMPK that enhance fatty acid synthesis 5 ; these proteins directly regulate the expression of genes encoding lipogenic enzymes, such as ACC, FAS, and SCD1, thereby inducing steatosis. 41,42 It is also known that AMPK regulates the expression of PPAR-α, which plays a central role in fatty acid oxidation. 43 PPAR-α is known to regulate the expression of the rate-limiting enzyme of β-oxidation, CPT1, 44 and to downregulate the expression of these two genes in ALD-affected rats. 45 In this study, we found that phosphorylated AMPK was reduced in the EtOH-treated group, which resulted in increased expression of lipogenic proteins and decreased expression of β-oxidation-related proteins.
However, Nonitri treatment reversed the expression of these proteins, thereby reducing lipid accumulation. Therefore, it is suggested that Nonitri inhibits hepatic fat deposition by decreasing lipogenesis and increasing fatty acid β-oxidation by improving the EtOH-induced inhibition of AMPK phosphorylation.
PC has been used as a functional food material to ameliorate alcoholic liver damage and hangovers. Although PC did not improve gut health, it improved liver damage caused by alcohol. 46 Our study well elaborates the molecular mechanism and potential of Nonitri to inhibit alcohol-induced liver damage. It also demonstrated a reduction of endotoxin levels by improving gut health and inhibiting fat deposition by modulating lipid metabolism by modulating the two major regulatory pathways. Moreover, the effects of Nonitri were also confirmed by correlation analysis between metabolic parameters and gut microbiota. As expected, Alistipes and Bacteroides were positively correlated with ALD parameters, and Muribaculum negatively correlated with these indices. These correlations between gut microbiota and ALD-related markers support previous studies that alcohol intake induces ALD through the gut–liver axis. 47
Although several researchers have reported that M. citrifolia causes hepatotoxicity, 48 –50 many studies report that high doses of M. citrifolia do not cause toxicity. 51 –54 Therefore, the use of Nonitri, a commercialized M. citrifolia extract, is considered promising for the development of functional food materials.
In this study, Nonitri regulated ALD-related gut microbial composition, such as that of Alistipes, Bacteroides, and Muribaculum, and restored gut barrier function. Moreover, Nonitri reduced the increased expression of inflammatory cytokines and regulated the expression of lipid metabolism-related proteins in the liver. Taken together, our results suggest that Nonitri holds potential as a functional food for improving alcoholic liver damage through the improvement of the gut–liver axis.
Footnotes
AUTHORs' CONTRIBUTIONS
H.-Y.P. designed the research; M.H.D. and M.P. analyzed data; M.H.D., E.L., H.H.L.L., M.-J.O., and M.K. performed the experiments; M.H.D. and H.-Y.P. wrote the article.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This work was supported by the Technology Development Program (Grant No. S2954137) funded by the Ministry of SMEs and Startups (MSS, Republic of Korea). This research was also supported by the Main Research Program (Grant No. E0210602-02) of the Korea Food Research Institute (KFRI) funded by the Ministry of Science and ICT.
SUPPLEMENTARY MATERIAL
Supplementary Table S1