
Abstract
Osteoporosis is a progressive metabolic disease characterized by decreased bone mineral density and increased fracture risk. Previous studies have shown that higher intake of vitamin K (VK) correlates with a reduced risk of osteoporosis. However, the effect of menaquinone-4 (MK-4), a specific form of VK, still remains obscure. Therefore, in this study, we investigated the effects of MK-4 on osteoclast differentiation by differentiating RAW 264.7 cells into osteoclasts with the help of receptor activator of nuclear factor-kappa B ligand (RANKL), assessed the mRNA expression of osteoclast-specific genes, and studied the effects of MK-4 in vivo in ovariectomized mice, a postmenopausal osteoporosis murine model. MK-4 inhibited osteoclast differentiation, decreased the mRNA expression of nuclear factor of activated T cells c1 (NFATc1), osteoclast-associated receptor (OSCAR), and cathepsin K (CTSK), and inhibited bone loss in ovariectomized mice. The findings strongly suggest that MK-4 is a therapeutic alternative for postmenopausal osteoporosis.
INTRODUCTION
Osteoporosis is a progressive metabolic disease in which decreased bone mass and bone microarchitecture lead to an increased susceptibility to fractures. 1 It typically occurs in postmenopausal women due to the decrease in estrogen hormone levels, secondary to reduced ovarian function, disrupting the dynamic balance between bone resorption by osteoclasts, and bone formation by osteoblasts. 2,3
During recent years, RAW 264.7 cells, belonging to the monocyte lineage /macrophage lineage, have been extensively used as a cell model for osteoclast differentiation to study bone diseases such as osteoporosis. Previous studies have shown that receptor activator nuclear factor-kappa B ligand (RANKL) plays a vital role in the activation, differentiation, and proliferation of osteoclasts, where RANKL stimulates tumor necrosis factor receptor-associated factor 6 (TRAF6). 4 TRAF6 further activates both mitogen-activated protein kinase (MAPK) and nuclear factor-kappa-B (NF-κB). 5
This leads to the activation of nuclear factor of activated T cells c1 (NFATc1) and c-Fos, transcription factors essential for the differentiation of osteoclasts. 6,7 These transcription factors play a key role in regulating the expression of several osteoclast-related genes and proteins such as tartrate-resistant acid phosphatase (TRAP), osteoclast-associated receptor (OSCAR), and cathepsin K (CTSK), suggesting the importance of RANKL in osteoporosis and other bone diseases. 8,9
Vitamin K (VK), a fat-soluble vitamin, plays a crucial role in blood clotting and bone metabolism. VK is naturally found in green leafy vegetables as phylloquinone (VK1), whereas menaquinones (VK2; MK-s, where s is the number of isoprenyl side-chain units), including short-chain MK-4, are produced by the conversion of VK1 to VK2, and other long chain MKs (MK-7 to MK-10) are synthesized by microorganisms, including intestinal bacteria. 10 During recent years, the importance of VK2 has been recognized for its role in heart and bone health, prevention of cancer, and regulation of calcium metabolism. 11,12
Recently, a study on ovariectomized (OVX) mice, a postmenopausal osteoporosis murine model, showed that VK2 (MK-4) decreased bone loss by promoting osteoblast activity and inhibiting osteoclast differentiation. 13 However, the mechanism underlying the MK-4 inhibitory effect on osteoclasts remains elusive. Therefore, in this study, we investigated the effects of MK-4 on RANKL-induced osteoclast differentiation in vitro using RAW 264.7 cells and on bone loss in vivo in OVX mice.
MATERIALS AND METHODS
Chemicals and reagents
Vitamin K2 (MK-4) and the TRAP staining kit were purchased from Sigma-Aldrich (St. Louis, MO, USA). The NFATc1 antibody was supplied by Santa Cruz Biotechnology, Inc., (Santa Cruz, CA, USA).
Cytotoxicity assay
RAW 264.7 cells were obtained from Korea Cell Line Bank (Seoul, Korea) and cultured in Dulbecco's modified Eagle's medium containing 10% fetal bovine serum and 1% penicillin–streptomycin (Gibco, Gaithersburg, NY, USA) at 37°C under 5% CO2. For the cytotoxicity assay, RAW 264.7 cells were cultured in a 48-well plate at 5 × 104 cells/well and treated with different concentrations of MK-4 (6.25, 12.5, 25, 50, 100, and 200 μg/mL) and incubated at 37°C for 24 and 48 h.
Thereafter, 400 μL WST-1 (2-(4-iodophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium sodium salt) (EZ-Cytox, DOGEN, Daejeon, Korea) solution was added to each well, and the plates were incubated at 37°C for 2 h. The absorbance of the samples was measured at 450 nm wavelength using a microplate reader (Infinite 200; Tecan Trading AG, Switzerland).
TRAP staining and measurement of TRAP activity
RAW 264.7 cells were plated in 48-well plates at 2 × 104 cells/well and treated with 50 ng/mL RANKL per well (R&D Systems, Minneapolis, MN, USA) and different concentrations of MK-4 (50, 100, and 200 μg/mL) and incubated at 37°C for 5 days. Medium and cell factors were changed every 2 days. After 5 days, the cells were washed, fixed, and stained for TRAP using the TRAP staining kit following the manufacturer's instructions. TRAP-positive multinucleated cells with more than three nuclei were counted as osteoclasts by observing under an inverted light microscope (Olympus, Tokyo, Japan).
For measuring TRAP activity, the culture medium was collected and TRAP activity was measured using a TRAP assay kit (Sigma-Aldrich) following the manufacturer's instructions. The absorbance of the samples was detected at 450 nm wavelength using an ELISA microplate reader.
RT-qPCR analysis
The total RNA was extracted using TRIzol reagent, per the manufacturer's instructions, from RAW 264.7 cells. cDNA was synthesized from 1 μg of RNA using a reverse transcription kit (BioFACT™, Daejeon, Korea). qPCR was performed using a SYBR® Green Realtime PCR Master Mix (BioFACT). All experiments were performed according to the manufacturer's instructions. The amplification program was as follows: heating at 95°C for 10 min, and then 40 cycles at 95°C for 15 s and 60°C for 1 min.
The primers used in this experiment were as follows: mouse NFATc1: forward, 5′-CTC GAA AGA CAG CAC TGG AGC AT-3′ and reverse, 5′-CGG CTG CCT TCC GTC TCA TAG-3′; mouse OSCAR: forward, 5′-GGG GTA ACG GAT CAG CTC CCC AGA-3′ and reverse, 5′-CCA AGG AGC CAG AAC CTC GAA ACT-3′; mouse CTSK: forward, 5′-ACG GAG GCA TTG ACT CTG AAG ATG-3′ and reverse, 5′-GTT GTT CTT ATT CCG AGC CAA GAG-3′; mouse GAPDH: forward, 5′-TGC CAG CCT CGT CCC GTA GAC-3′ and reverse, 5′-CCT CAC CCC ATT TGA TGT TAG-3′. The relative mRNA expression was normalized to the level of glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
Osteoporosis model and treatment
Ovariectomized (OVX) mice, a mouse model for postmenopausal osteoporosis, were used for the in vivo study. All animal procedures were approved by the animal care and use committee of Korea Food Research Institute (KFRI-M-20047). Female C57BL/6 mice were acclimatized for a week and placed under standard conditions with free access to standard pellet feed and clean water before surgery. After 1 week, the ovariectomy and sham operation were performed; both ovaries were removed from mice in the OVX groups.
After 1 week, the mice were randomly divided into five groups (n = 8/group): (1) sham group (sham surgery without treatment), (2) ovariectomy control group orally fed only corn oil (OVX), (3) ovariectomy group treated with a low dose of MK-4 (OVX+MK4_L), (4) ovariectomy group treated with a medium dose of MK-4 (OVX+MK4_M), and (5) ovariectomy group treated with a high dose of MK-4 (OVX+MK4_H). MK-4 was administered orally with corn oil as the vehicle daily for 14 weeks, and mice in the low-, middle-, and high-dose groups received 0.9, 6.3, and 37.8 mg/kg MK-4 as per their body weight, respectively. The body weight of each mouse was monitored for the adjustment of the oral dose and the data were recorded weekly.
Serum biochemical marker
Blood was collected, and serum was separated through centrifugation and stored at −80°C. For determining bone alkaline phosphatase (BALP) activity, the serum was prepared using the BALP kit (DKKorea, Inc., Seoul, Korea) according to the manufacturer's instructions, and the activity was measured using an ELISA microplate reader.
Assessment using bone micro-CT
The femur was measured using micro-CT (GE eXplore CT 120; GE Healthcare, Milwaukee, United Kingdom). The scanning parameters were set at 18 mm and exposure was performed at 80 kV, 32 mA, and 200 ms/frame. Images were analyzed using the MicroView® version 2.5 software (GE Healthcare), and two-dimensional and three-dimensional images were obtained. Bone morphometric parameters such as bone mineral density (BMD), bone surface/bone volume (BS/BV), bone surface area/total volume (BS/TV), bone volume/total volume (BV/TV), trabecular thickness (
Statistical analysis
All results are expressed as means ± standard errors (SEM). Significant differences were determined using a one-way ANOVA followed by post hoc Tukey's test. Values with P < .05 were considered statistically significant.
RESULTS
Effect of MK-4 on cell cytotoxicity and RANKL-induced TRAP activity
RAW 264.7 cells were treated with different concentrations of MK-4 (6.25, 12.5, 25, 50, 100, and 200 μg/mL) in vitro to analyze the cytotoxic effect of MK-4 from 24 to 48 h. No antiproliferative effects on RAW264.7 were observed with the treatment of MK (except 200 μg/ml) (Fig. 1).
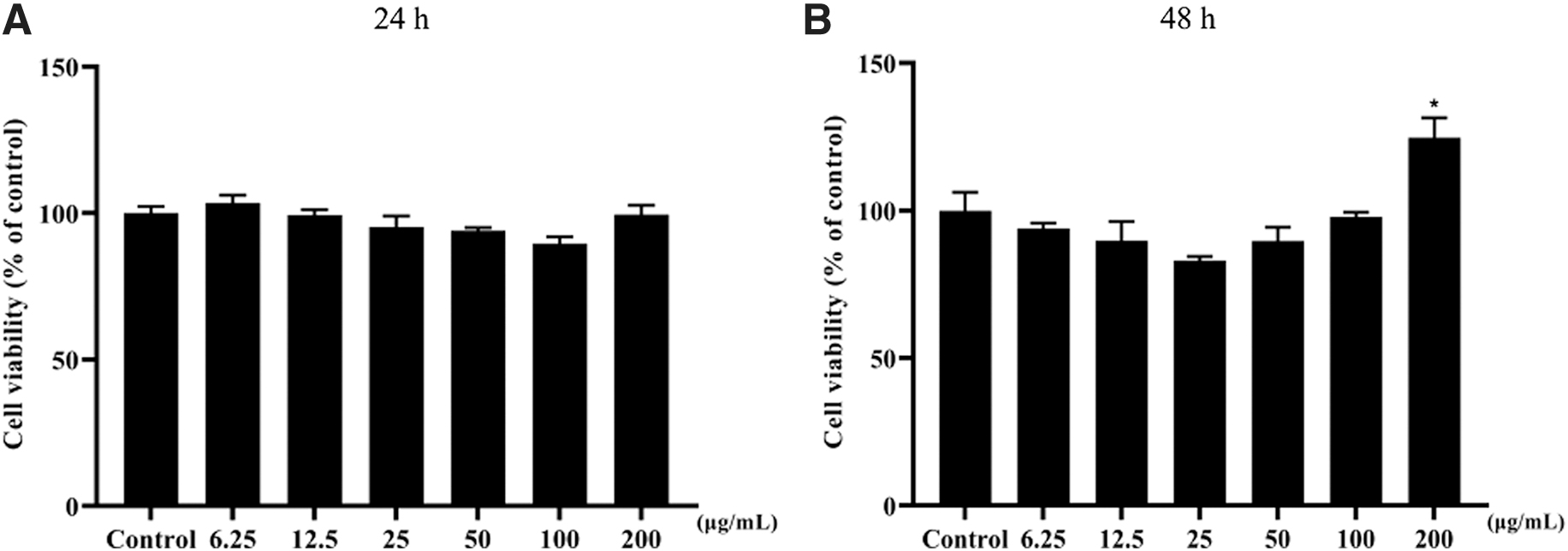
Cytotoxicity effects of RAW264.7 cells treated with MK-4 for 24 h
The effect of MK-4 on RANKL-induced osteoclast differentiation was determined using the TRAP staining assay. RAW 264.7 cells treated with RANKL alone showed a higher number of TRAP-positive cells than the normal control group (Fig. 2A). However, a significant dose-dependent decrease in the number of TRAP-positive cells in MK-4-treated RAW 264.7 cells was observed. Moreover, significant decrease in TRAP activity was observed in the MK-4-treated cell group in a dose-dependent manner compared with that in the RANKL-treated group (Fig. 2B).
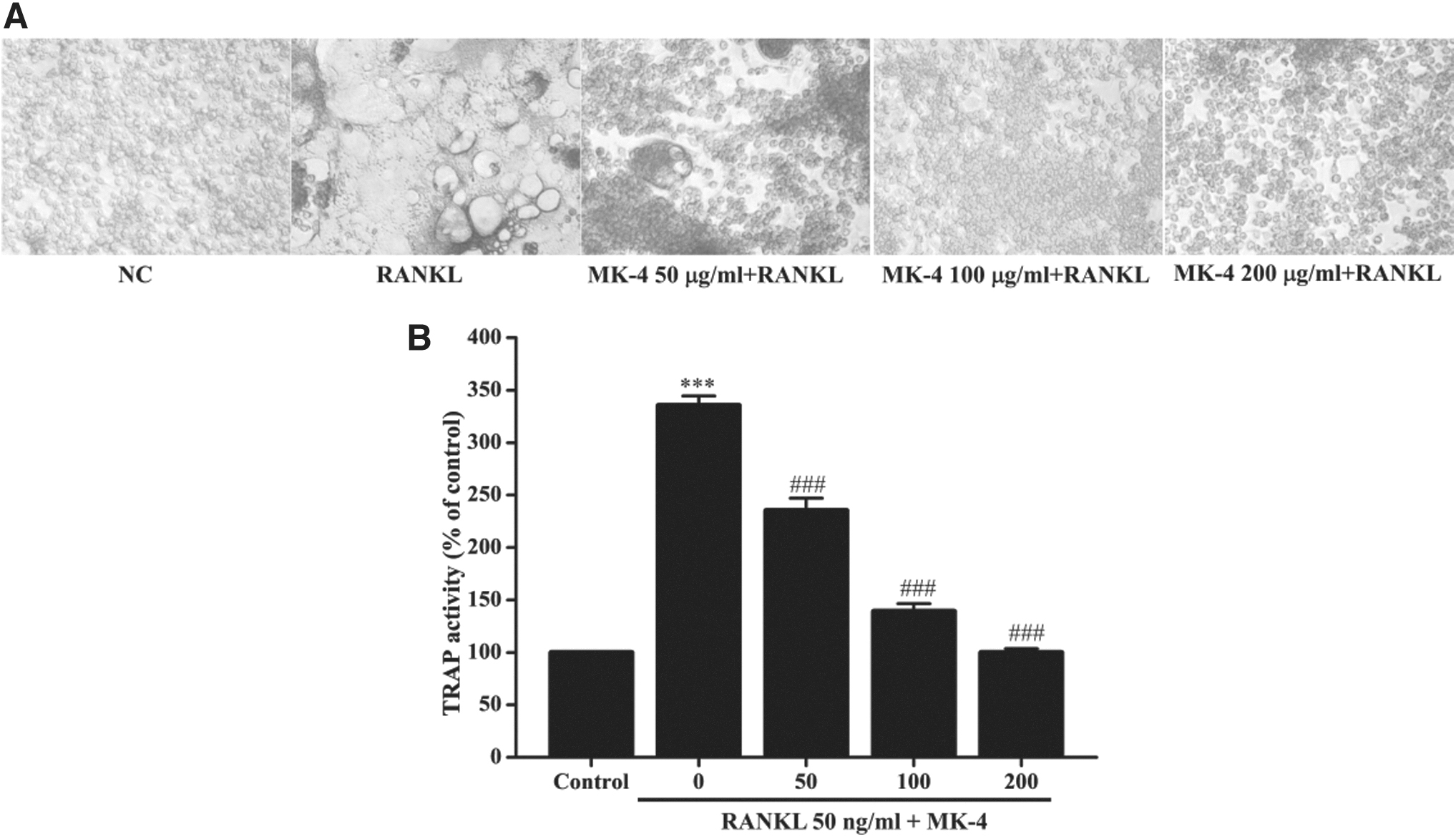
Effects of MK-4 on osteoclast differentiation.
Effect of MK-4 on RANKL-induced expression of osteoclast-related genes
The effect of MK-4 on the mRNA expression of osteoclast-related genes was measured using RT-qPCR. Increased expression of NFATc1, CTSK, and OSCAR was observed in cells treated with RANKL alone (Fig. 3A–C). Treatment with MK-4 significantly decreased their mRNA expression in a dose-dependent manner. These results demonstrate that MK-4 efficiently inhibits in vitro osteoclastogenesis by suppressing the expression of key genes.
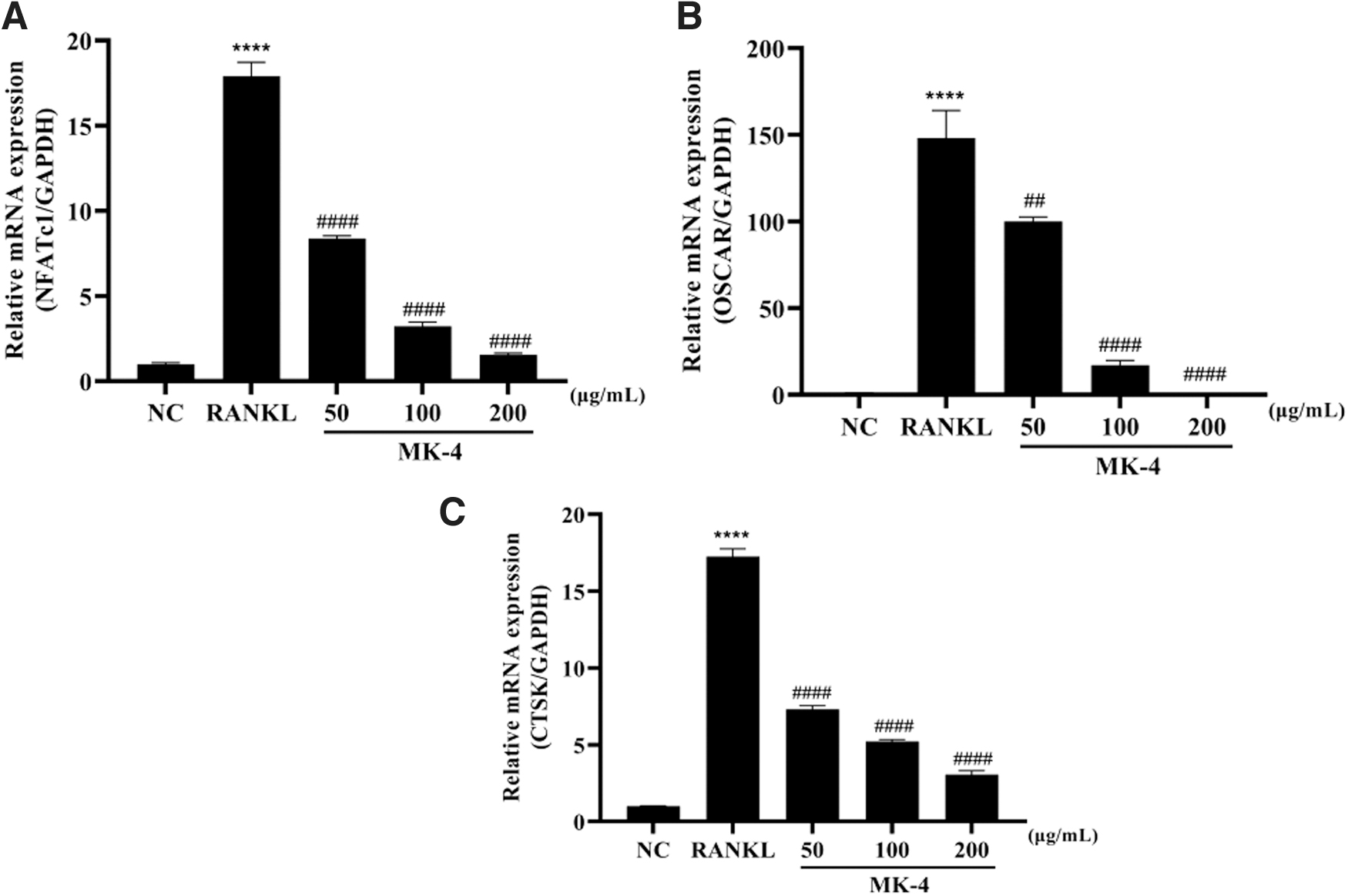
Effects of MK-4 on the expression of osteoclast-related genes in RANKL-induced RAW 264.7 cells. The mRNA expression of
Effect of MK-4 on body and uterine weights and BALP in ovariectomized mice
The effect of MK-4 on OVX-induced osteoporosis was evaluated by orally administering low, medium, and high concentrations of MK-4 to the OVX-induced mice for 14 weeks. The effects of MK-4 on body and uterine weights and BALP levels of mice are summarized in Table 1. The body weights of the OVX group and the MK-4-treated OVX group mice were significantly higher than those of the sham group. However, the body weights of mice in the OVX+MK-4-L group were significantly lower than those of mice in the other treatment groups.
Effect of Menaquinone-4 on Body and Uterus Weights and Bone Alkaline Phosphatase Levels of OVX Mice
P < .05.
P < .001 versus sham group.
P < .05.
P < .01.
P < .001 versus OVX group.
BALP, bone alkaline phosphatase.
The uterine weights of the OVX group mice were lower than those of the sham group. However, the higher uterine weights of mice in the MK-4-L and MK-4-M groups were higher than those of mice in the OVX group. The treatment had no effect on the uterine weights of mice in the MK-4-H group. The serum level of BALP was significantly higher in mice of the OVX group, but decreased to nearly normal levels in the MK-4-treated groups (Table 1).
Effects of MK-4 on bone structure in OVX mice
The BMD of the femoral heads of mice in the OVX group was lower than those of the mice in the sham group (Fig. 4A). Furthermore, MK-4-L significantly increased the bone density in the femoral head in OVX mice, although no such significant increase was observed in the MK4-M and MK-4-H groups. Significant decreases in BV/TV, Tb.Th, and Tb.N values were observed in the OVX group compared with those in the sham group. In particular, substantial increases in BV/TV, Tb.Th, and Tb.N values were observed in the MK-4-L group compared with those of the OVX group. In addition, Tb.Sp was significantly higher in mice of the OVX group than in mice of the sham group. However, Tb.Sp was significantly lower in mice of the MK-4-L group than in mice of the OVX group (Fig. 4B–F).

Effects of MK-4 in mice with ovariectomy (OVX)-induced bone loss.
DISCUSSION
Bisphosphonate and selective estrogen receptor modulators are frequently used for the treatment of osteoporosis; however, they are associated with adverse effects. 14,15 Consequently, a need for safer agents such as calcium, vitamin D3, vitamin K, and bioactive compounds with less undesirable adverse effects to prevent postmenopausal osteoporosis is imperative. 16,17 Previous studies have revealed that VK2 may play a crucial role in bone metabolism and bone-related diseases. 13,18
RANKL, a pro-osteoclastogenic cytokine, plays a crucial role in osteoclast differentiation of RAW 264.7 cells. TRAP staining is a standard method used to identify osteoclast expression and activation. 8 In this study, MK-4 treatment reduced the TRAP-positive cells and inhibited TRAP activity.
A previous study has shown that NFATc1-deficient mice developed osteoporosis due to osteoclast differentiation being blocked, and c-Fos knockout mice developed osteoporosis due to attenuated osteoclast function, which suggests that c-Fos and NFATc1 are key transcription factors for osteoclast differentiation. 8,19 –21 In addition, NFATc1 increased the expression of RANKL-induced osteoclast-specific genes such as TRAP, MMP-9, CTSK, ATP6v0d2, and OSCAR. 8,22 –24 CTSK is a cysteine protease expressed in osteoclasts and is important for the degradation of the bone organic matrix during bone resorption. 25,26
Therefore, CTSK is an effective target in the treatment of osteoporosis. The protein OSCAR is expressed during preosteoclast differentiation to mature osteoclast differentiation and may contribute to the etiology and severity of osteoporosis and rheumatoid arthritis. 27,28 In this study, MK-4 significantly decreased the expression of NFATc1 in RANKL-induced RAW 264.7 cells and suppressed the expression of selected osteoclast-related genes (NFACTc1, CTSK, and OSCAR). These results suggest that MK-4 may possess antiosteoporotic activity by regulating the expression of bone resorption markers.
In a previous study, rats with OVX-induced osteoporosis shared symptoms of osteoporosis with humans, such as the increase in body weight and reduction in uterine weight and bone mass as a postmenopausal osteoporosis, which is similar to our observations in this study, where the OVX group mice showed increased body weights and lower uterine weights. 29 –31 We used younger mice (7 weeks old) and treated them with lower doses than previous studies, which used relatively older rats (12–16 weeks old) and treated them with higher doses. Our study shows a specific effect of a low concentration of MK-4 in the species of mice used in this study, at a specific age, and for a certain administration period.
For this reason, the uterine weight of the mice did not seem to have changed in the high MK-4 concentration group. ALP, a marker of bone formation (and specifically BALP), is increased in the serum of postmenopausal women. 32 Although daily adequate intake levels have been determined for vitamin K1, there are no established recommended daily allowances for vitamin K2. 33 Some studies suggested that a dosage as low as 1.5 mg/day may be beneficial in maintaining healthy bones in healthy postmenopausal women. 34,35 In addition, some animal studies have confirmed that MK-4 protects against bone loss induced by ovariectomy in rats at a dosage of 3 mg/kg. 36,37
Based on the finding of these studies, we administrated of MK-4 orally in the 0.9, 6.3, or 37.8 mg/kg body weight, respectively. The novelty of this study is that we used younger mice (7 weeks old) and treated them with lower doses than previous studies, which used relatively old rats (12–16 weeks old) and treated them with higher doses. A previous study reported that positive effects of MK-4 administration were observed in a group of younger postmenopausal of women (55–65 years) compared with those in a group of older women (65–75 years). 38 We demonstrated that the oral administration of MK-4 suppressed the body weight gain and production of BALP induced by OVX in our mouse model.
Micro-CT has been used to study the structural properties of bones in three dimensions. 39 Bone density and bone microstructure are important measures in the evaluation of calcium content and bone quality in the OVX mouse model. 40 In this study, the reduction in BMD, BV/TV, Tb.Th, and Tb.N in OVX mice was improved with the administration of low-dose MK-4. Tb.Sp was significantly higher in mice in the OVX group than in mice in the sham group, whereas it was significantly lower in mice in the MK-4-L group. These results suggest that low-dose MK-4 significantly inhibited bone loss in the OVX-induced osteoporosis mouse model.
This study has some of the following limitations: In the in vitro study, we focused on the role of MK-4 in osteoclast differentiation and the corresponding NFATc1 signaling; however, we need to further evaluate the deeper mechanism regarding MAPKs and NF-κB. In the in vivo study, we did not use various dose levels to better evaluate the improvement of BMD and bone microarchitecture. Further studies are required to determine the appropriate dose levels.
In conclusion, the results of this study suggest that MK-4 effectively inhibits RANKL-mediated osteoclast formation through the suppression of NFATc1 expression, and subsequent suppression of osteoclast-related genes (NFATc1, CTSK, and OSCAR). In addition, MK-4 suppresses the reduction in bone thickness and number and considerably increases BMD and trabecular area in the OVX model. These results indicate that MK-4 may serve as a novel supplement for the prevention and treatment of metabolic bone diseases such as osteoporosis.
AUTHORs' CONTRIBUTIONS
A.L., A.H., and S.S. contributed to methodology; M.S. analyzed data; A.H. and S.S. were involved in editing; S.H. carried out formal analysis and review; S.L. was in charge of conceptualization and writing.
Footnotes
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This study was supported by a grant from the Korea Food Research Institute (E0210102) and the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, and Forestry (Grant Nos. GA121047 and GA122055).