
Abstract
Osteoporosis is characterized by low bone mass and elevated structural deterioration of the bone tissue, resulting in bone weakness with an increased risk of fracture. Considering biological activities of various phytochemicals extracted from apples, we herein demonstrated the potential antiosteoporotic effects of apple-derived nanovesicles (apple NVs) using osteoblastic MC3T3-E1 cells. Apple NVs significantly stimulated the growth of MC3T3-E1 cells. The cellular alkaline phosphatase (ALP) activity was significantly upregulated in the 5 μg/mL apple NVs-treated group. In addition, the concentrarion of mineralized nodules was significantly increased in the apple NVs-treated groups. Furthermore, apple NVs increased the expression of the genes and proteins associated with osteoblast growth and differentiation, such as Runx2, ALP, OPN, and BMP2/4, which further activated ERK- and JNK-related mitogen-activated protein kinase signaling. These results demonstrate that apple NVs have a potential to prevent osteoporosis by promoting osteoblastogenesis in osteoblastic MC3T3-E1 cells through regulating the BMP2/Smad1 pathways.
INTRODUCTION
Osteoporosis is a progressive skeletal disorder with decreased bone mass and increased structural deterioration of bone tissue, leading to bone weakness with an increased risk of fracture. 1 Most drugs currently used to prevent and treat osteoporosis exert therapeutic effects by inhibiting bone resorption, which has limited theraepeutic potential in the complete prevention of bone loss. 2 –4 Numerous studies have investigated how to prevent and treat osteoporosis, 5 –8 which highlight several herbal medicines and natural plant compounds that facilitate bone tissue regeneration. 9,10 Sacco and Horcajada demonstrated a positive correlation between the dietary intake of phytonutrients and bone health, which emphasized the potential benefits of these phytonutrients to prevent osteoporosis. 11
Several phytochemicals extracted from apples and apple by-products have been reported to exert various biological activities and health-promoting effects. 12 Apples contain various polyphenolic compounds, such as catechin, epicatechin, phloridzin, phloretin, procyanidin, chlorogenic acid, hydroxycinnamates, rutin, and quercetin. 13 Several studies have shown that polyphenols are effective in the prevention of cancer, arteriosclerosis, hypertension, diabetes, inflammation, and aging by inhibiting the free radical-induced oxidation of the lipid membranes. 14,15 In addition, anti-inflammatory effects of apple polyphenolic compounds have been widely explored in vitro and in vivo. 16,17
Nanovesicles (NVs) produced and secreted by plants are generally classified into different types, according to their dimensions. 18 –20 NVs transport cargo molecules, such as proteins, lipids, mRNA, or miRNA in the fruit juice and plants. 18 –20 Recently, several reports suggested that the exosome-like nanoparticles are also produced in various plants, fruits, and mushrooms. 21 –25 Plant-derived NVs exhibit various beneficial effects, such as anti-inflammatory, 22,23 antioxidant, 24 and antitumor properties. 25,26 However, the effects of plant-derived NVs on bone health and disease are not fully elucidated. Therefore, we herein investigate functions and mechanisms of apple-derived NVs (apple NVs) in the regulation of bone formation using osteoblastic MC3T3-E1 cells as model systems.
MATERIALS AND METHODS
NV separation
Apples used in this study were the Fuji (Malus pumila Mill) strain purchased from Andong Nonghyup, which were harvested in Andong, Gyeongsangbuk-do. Apple NVs were obtained by using an optimized method (Fig. 1A). 27,28 After apples were washed with clean water, seeds were removed. Flesh along with the pulp and skin were pulverized (300 g) with cold 1 × PBS (phosphate-buffered saline) for 5 min. The supernatant fraction was collected by sequential centrifugations (once at 500 g for 10 min, 2000 g for 20 min and twice at 10,000 g for 30 min). The supernatant was again centrifuged at 100,000 g for 2 h. Finally, the resulting pellet was dissolved in 1 mL of 1 × PBS and used as apple NVs.
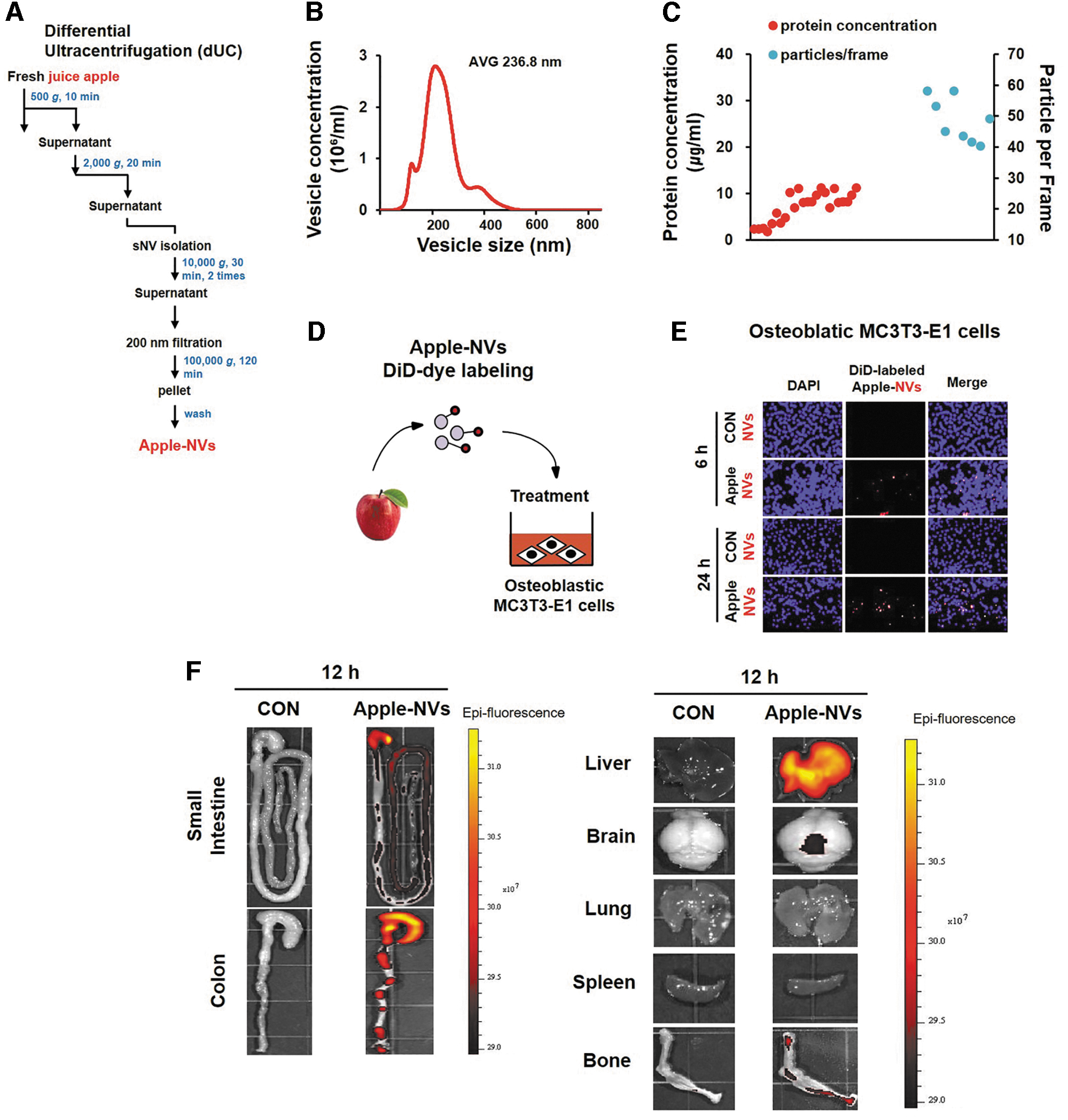
Isolation and characterization of apple NVs.
DiD-DAPI staining
The internalization of apple NVs into preosteoblastic MC3T3-E1 cells was investigated using fluorescence microscopy assisted with DiD-DAPI staining. For DiD labeling, apple NVs (1 mg in 1 mL PBS) were mixed with 1 μL of 5 mM DiD (1,1′-Dioctadecyl-3,3,3′,3′-Tetramethylindodicarbocyanine Perchlorate, Invitrogen) in dimethyl sulfoxide and incubated at 25°C for 30 min. MC3T3-E1 cells were incubated with DiR-labeled apple NVs-DiR for 6 or 24 h. Cells were fixed with 4% paraformaldehyde in the microscope coverslips for 10 min at room temperature (RT). The fixed cells were washed with PBS three times, permeabilized with 1% Triton X-100 for 10 min, washed with PBS again three times before staining with 4′,6-diamidino-2-phenylindole (DAPI) for 15 min. Cell images were obtained using a fluorescence microscope with a digital camera.
Nanoparticle tracking analysis
Nanosight NS300 instrument (Malvern Panalysis Ltd., Malvern, Worcestershire, UK) was used to measure the size and concentration of apple NVs whose sizes ranged from 50 to 1000 nm in suspension.
Cell culture and osteoblast differentiation
MC3T3-E1, a murine preosteoblastic subclone 4 cell line, was purchased from the American Type Culture Collection (CRL-2593, Manassas, VA, USA). They were grown in alpha minimum essential medium (α-MEM; Gibco Laboratories) supplemented with 10% (v/v) fetal bovine serum and 1% (v/v) penicillin-streptomycin (P/S) at 37°C in an incubator with 5% CO2 atmosphere. After reaching 80% confluence, cell culture medium was replaced with a complete differentiation medium (DM) containing ascorbic acid (100 μg/mL) and β-glycerophosphate (10 mM) for osteoblast differentiation. To examine the effect of apple NVs on osteoblast differentiation, cells were grown in a DM containing different concentrations of apple NVs (0, 1, 5, or 10 μg/mL). The culture media were replaced every 3 days to induce differentiation of osteoblasts for 7 days.
Assessment of cell proliferation using the MTT assay
MC3T3-E1 cells in α-MEM were seeded into 96-well microplates at a density of 1 × 103 cells/well and then incubated for additional 24 h. After cells were washed twice with PBS, cell medium was replaced with a DM containing apple NVs at a concentration of 0, 1, 5, or 10 μg/mL for 3 and 7 days, to induce the differentiation and determine the rates of mineralization in osteoblastic MC3T3-E1 cells, as described in a previous report. 29
Alkaline phosphatase activity assay
MC3T3-E1 cells in α-MEM were seeded into 12-well microplates at a density of 1 × 104 cells/well, followed by incubation for another 24 h. After cell medium was replaced with DM with a different concentration of apple NVs (0, 1, 5, or 10 μg/mL), cells were incubated for 3 and 7 days as the previous study. 29,30 Alkaline phosphatase (ALP) activity of each sample was normalized to the protein concentration. Cellular ALP activity was expressed as units (nmol PNPP/mg of protein/min), while medium ALP activity was shown as units/mL (nmol PNPP/mL/min).
ALP staining
ALP activity in the differentiated MC3T3-E1 cells was visualized using ALP staining. In brief, cells incubated with apple NVs (0, 1, 5, or 10 μg/mL) for 3 or 7 days were rinsed with PBS and stained using a mixture of naphthol AS-Mx phosphate disodium salt, N,N-dimethyl formamide, and fast red salt at 37°C for 30 min or until the cells became yellow in appearance. 29,30 After washing with distilled water, the cells were photographed under a light microscope. ALP-positive cells were visualized in red/purple color.
Von Kossa staining
MC3T3-E1 cells (1 × 104 cells/well) were seeded in 12-well plates, followed by incubation in growth media. At 100% confluency, the cells were treated with 0, 1, 5, or 10 μg/mL apple NVs, followed by incubation for 3 and 7 days. Extracellular matrix mineralization was assessed by staining with 5% silver nitrate, incubation under ultraviolate light at RT for 1 h, and then were washed with distilled water. Images were captured using a light microscope. Mineralized nodules are shown as thick, dark brown stripes resembling cuboidal cells.
Alizarin red staining
Mineralized extracellular matrix in MC3T3-E1 cells were monitored using Alizarin red staining to chelate Ca ions as alizarin red forms. Before staining, MC3T3-E1 cells (1 × 105 cells/well) seeded in 12-well plates were incubated in an osteogenic medium containing different concentrations of apple NVs (0, 1, 5, or 10 μg/mL). After 3 and 7 days, cells were stained with 40 mM Alizarin Red S (pH 4.2) at RT for 30 min, as described in a previous study. 29,30 The stained cells were photographed using a phase-contrast microscope followed by a quantitative destaining procedure using 10% cetylpyridinium chloride prepared in 10 mM sodium phosphate (pH 7.0) for 15 min. The extracted stain through the destaining procedure was transferred to a 96-well plate, and absorbance determined at 570 nm using a spectrophotometer.
Collagen staining
Cellular matrices were examined using the van Gieson stain method to evaluate extracellular matrix collagen synthesis, as previously described. 29,30 Images of the stained cells were taken using a phase-contrast microscope with a digital camera (IM50, Leica, Germany).
Protein extraction and western blotting
After cells treated with apple NVs were washed twice with cold 1 × PBS, cellular proteins were extracted using RIPA buffer (Boston Bio Products) supplemented with a protease inhibitor cocktail (Sigma-Aldrich) and a phosphatase inhibitor cocktail (Sigma-Aldrich). The cell lysates were centrifuged at 13,000 g at 4°C for 10 min. The protein concentrations of diffetent cell extracts were quantified using a bicinchoninic acid protein assay (Thermo Fisher Scientific). Equal amounts of protein from different cell extracts were separated using sodium dodecyl sulfate-PAGE and transferred onto nitrocellulose membranes. The membranes were blocked for 1 h at RT with 5% nonfat dry milk prepared in Tris-buffered saline containing 0.05% Tween-20 (TBS-T), followed by incubation with the primary antibodies (against each indicated target protein) at 4°C overnight.
Afterward, the blots were washed three times with TBS-T and then incubated with the secondary antibodies conjugated with horseradish peroxidase (Santa Cruz Biotechnology, sc-2004). The signals were visualized using an enhanced chemiluminescence kit (Amersham Biosciences, Piscataway, NJ, USA). In addition, some membranes were stripped to determine the β-actin signal for the same membranes. The relative abundance of each band relative to β-actin was quantified via a densitometric scanning of each band using a Fusion Solo 6x basic 0.84 (Vilber, Collegien, Paris, France).
RNA isolation and quantitative real-time PCR
Total RNA quantity was determined using an RNeasy Mini Kit (Qiagen, Valencia, CA, USA). The isolated total RNA (1 μg) was reverse-transcribed using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA) according to the manufacturer's protocol. Real-time quantitative PCR (qRT-PCR) was carried out following the standard protocols using a fluorescence dye SYBR Master Mix Kit (Applied Biosystems). The PCR plates were sealed and run on a QuantStudio™ 1 real-time PCR instrument (Thermo Fisher) under the following thermocycling conditions: 50°C for 2 min, 95°C for 2 min, followed by 40 cycles at 95°C for 15 sec, and 60°C for 1 min. Target gene expression was analyzed by using the software (Applied Biosystems) after normalization to GAPDH (as the endogenous control).
In vivo imaging for studying biodistribution of orally administration apple NVs
All mice experiments were conducted according to the guidelines approved by the Institutional Animal Care and Use Committee (IACUC) of Andong National University.
Male C57BL/6 mice were administrated DiR dye-labeled apple NVs (1 mg per mouse in 200 μL PBS) and CON without DiR dye (200 μL PBS) by oral gavage and imaged over 12 h. The specific organs (liver, brain, lung, spleen, bone, small intenstine, and colon) were scanned for measuring the intensity of the time-dependent changes of the DiR fluorescence signals using the IVIS Lumina Series III In Vivo Imaging System (Perkin Elmer).
Statistical analysis
All data are presented as the means ± standard errors from at least three independent experiments. Statistical analyses were performed using the Statistical Package for the Social Sciences software package program (SPSS, Inc., Chicago, IL, USA). One-way analysis of variance using Duncan's test was used to assess statistical significance among more than three samples. P < .05 was considered significant.
RESULTS
Internalization of apple NVs into preosteoblastic MC3T3-E1 cells
The mean particle size of the apple NVs was 236.8 nm (Fig. 1B). Nanoparticle tracking analysis revealed the average number of particles per frame to be 43.1 (Fig. 1C). Whether the apple NVs were internalized into the MC3T3-E1 cells after 6 h was investigated using fluorescence microscopy assisted with DiD-labeled exosomes (red indicator) (Fig. 1D). As contrasted with the control group, the apple NVs were demonstrated to enter the osteoblastic MC3T3-E1 cells (Fig. 1E).
To determine the tissue distribution in mice, the in vivo biodistribution of DiR-labeled apple NVs was evaluated using IVIS imaging. The DiR fluorescent signals of orally administered apple NVs were predominantly detected in the GI tract, liver, and bone (Fig. 1F).
Effect of apple NVs on the proliferation of preosteoblastic MC3T3-E1 cells
The effect of apple NVs on the proliferation of MC3T3-E1 cells was assessed using the MTT assay. The cells were treated with apple NVs at three different concentrations (1, 5, and 10 μg/mL) and incubated for 3 and 7 days. All apple NVs significantly increased osteoblastic proliferation at 3 days (P < .05), in a concentration-dependent manner and this effect lasted at least for 7 days (Fig. 2A). These results suggested that apple NVs promote the proliferation of MC3T3-E1 cells.

Effect of apple NVs on the proliferation and ALP activity of osteoblastic MC3T3-E1 cells.
Effects of apple NVs on ALP activity in MC3T3-E1 cells
ALP is a secretory enzyme synthesized by osteoblasts and then secreted to the extracellular matrix, which is used as an early marker of osteoblast differentiation. Increased ALP activity is positively correlated with the progressive differentiation of cultured osteoblasts, which is responsible for the calcification of hard bone tissues. Thus, the effects of apple NVs on ALP activity in preosteoblastic MC3T3-E1 cells were investigated (Fig. 2B–D). Compared to those without apple NVs at 7 days, cells treated with apple NVs exhibited increased ALP staining (Fig. 2B), which was consistent with the increased ALP activity (Fig. 2C, D).
In fact, cellular ALP activity was significantly increased in 5 μg/mL apple NV-treated MC3T3-E1 cells for 3 and 7 days. Apple NVs at a concentration range of 1 to 10 μg/mL significantly increased medium ALP activity at days 3 and 7 compared to the control, indicating the release of ALP into the medium after exposure to apple NVs. These results indicate that apple NVs stimulated ALP activity, potentially inducing osteoblast differentiation and extracellular matrix mineralization in osteoporosis.
Effect of apple NVs on the bone nodule formation activity of MC3T3-E1 cells
Calcium deposition is frequently used as a marker of differentiated osteoblasts. 29 Therefore, the effects of exogenous apple NVs on calcium mineralization in MC3T3-E1 cells were evaluated. MC3T3-E1 cells were incubated with apple NVs in an osteogenic medium for 3 and 7 days, and then Von Kossa and Alizarin red S stainings were conducted. Von Kossa staining detects calcium phosphate in the extracellular matrix, while Alizarin red S staining visualizes calcium chelate.
An increase in Von Kossa staining intensity in the mineralized nodules was confirmed after 7 days treatment with 5 and 10 μg/mL apple NVs (Fig. 3A). Alizarin red S staining revealed higher calcium deposition in apple NVs-treated MC3T3-E1 cells than nontreated cells (Fig. 3B). Calcium deposition was further assessed, quantitatively (Fig. 3D): on day 3, the concentration of mineralized nodules significantly increased in all apple NVs-treated groups in a dose-dependent manner. On day 7, significant increases of the concentration of mineralized nodules were observed in the 5 μg/mL (111.32%) and the 10 μg/mL apple NV-treated group (115.69%). These results indicate that apple NVs potentially improved mineralization during osteoblast differentiation.
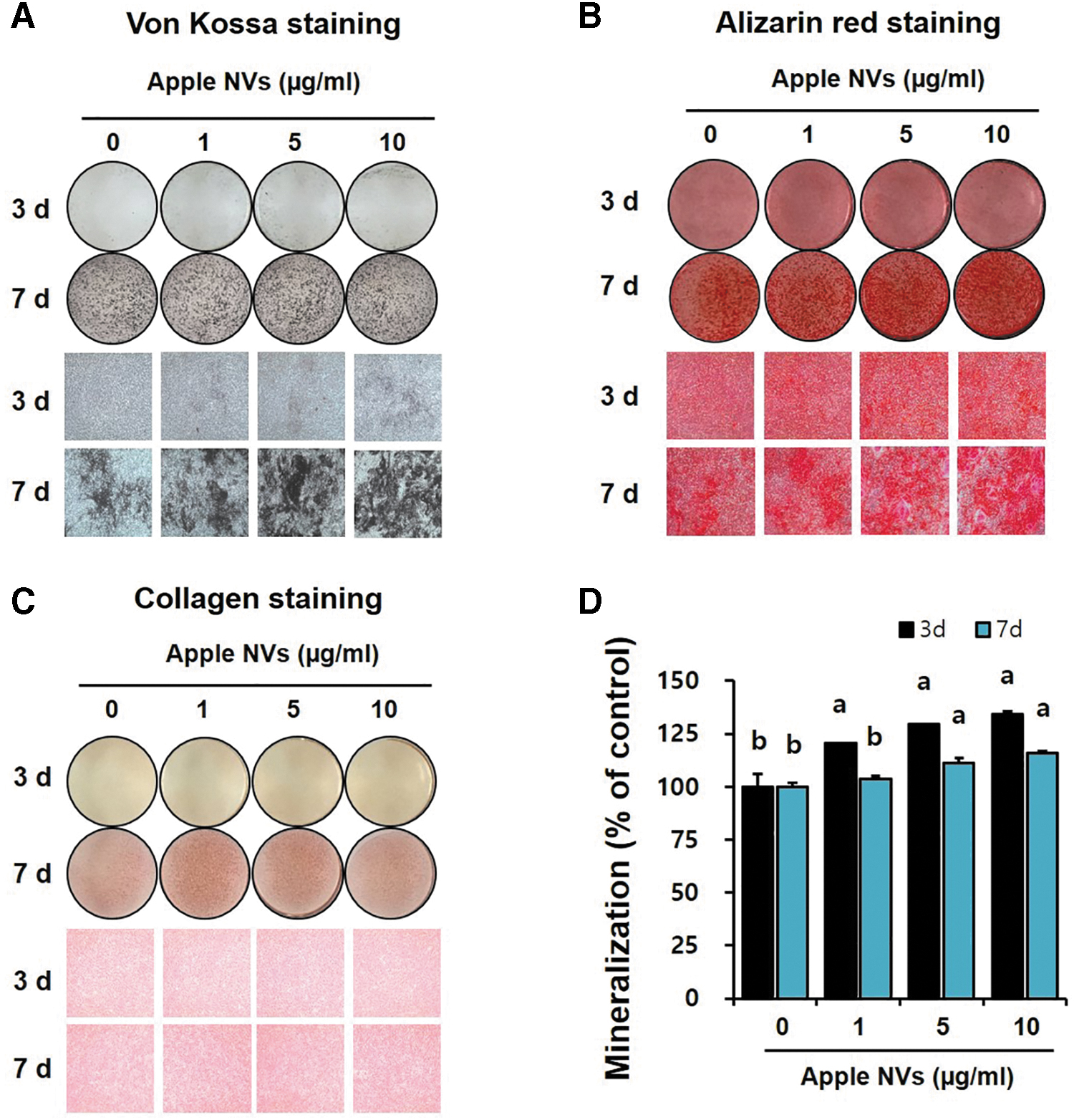
Effect of apple NVs on bone nodule formation in osteoblastic MC3T3-E1 cells. The representative images of three replicates in 12-well plates are shown.
As the increase in collagen synthesis and the accumulation of extracellular matrix proteins represent the maturation of the bone matrix and osteoblasts, 30 collagen deposition in the cell matrix was investigated using van Gieson's staining (Fig. 3C). Collagen synthesis was increased in the apple NVs-treated group, compared to that in the control group on day 7. Overall, apple NVs promoted the differentiation and mineralization of preosteoblastic MC3T3-E1 cells.
Effects of apple NVs on the expression of osteogenic markers
To investigate how apple NVs stimulate growth and differentiation in osteoblastic MC3T3-E1 cells, transcript-level expression of osteoblast differentiation marker genes was analyzed using qRT-PCR. Significant upregulation of Runx2, ALP, and OPN was observed in the MC3T3-E1 cells treated with apple NVs, compared with the control group (Fig. 4A). Next, the levels of the corresponding proteins in apple NV-treated cells were evaluated using western blotting. On day 3, the apple NV-treated groups showed significant and dose-dependent increases in the levels of Runx2, ALP, and OPN (Fig. 4B). These results suggest that apple NVs enhanced the expression of bone-related genes and proteins, leading to effective inducion of osteoblast growth and differentiation.
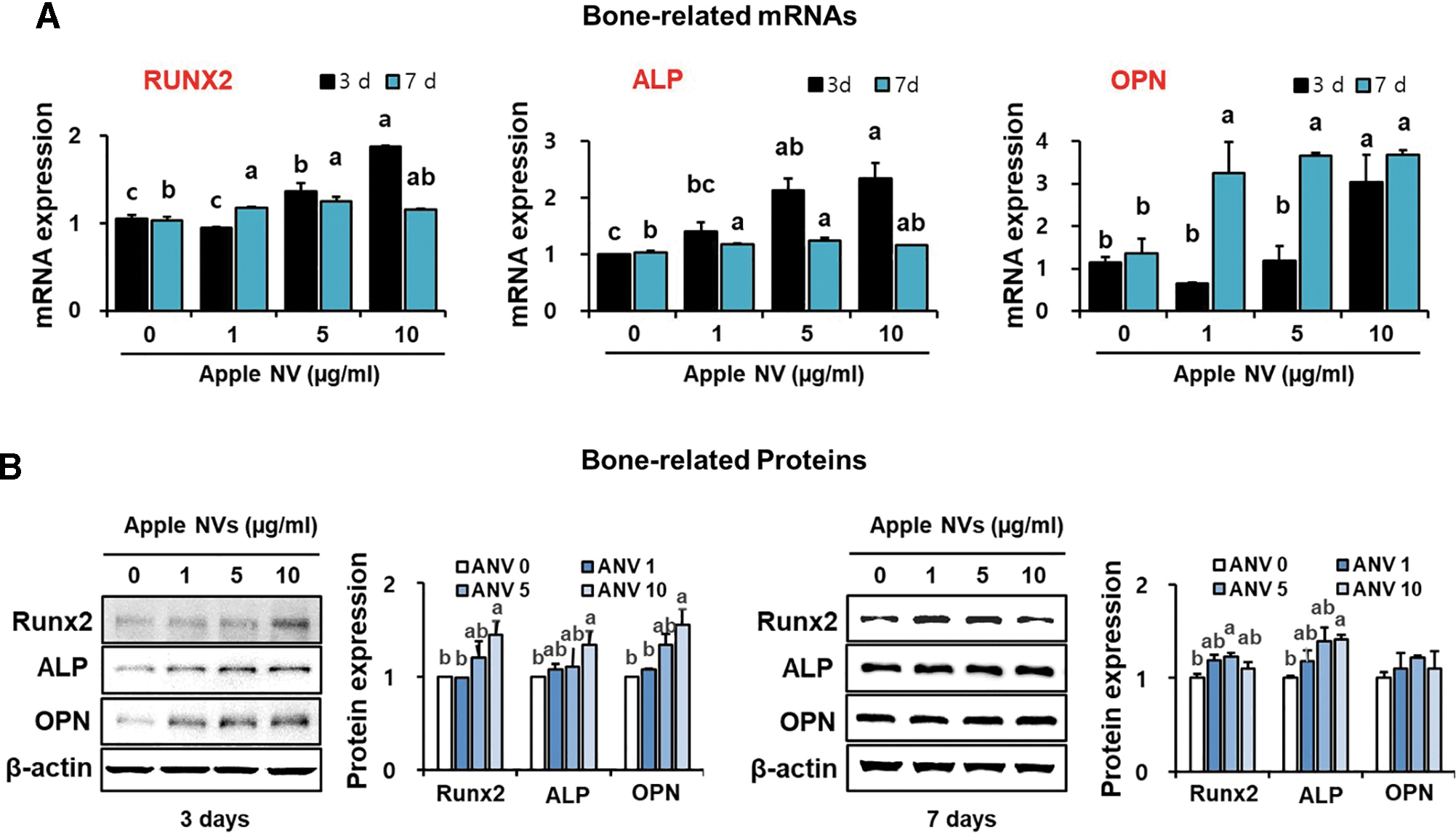
Relative expression of bone-related genes and proteins after treating osteoblastic MC3T3-E1 cells with apple NVs.
Bone-matrix protein (BMP) 2/4 signaling pathway was further investigated to elucidate the molecular mechanisms associated with osteoblast differentiation and activity, which is critically important in bone formation. The mRNA expression of BMP 2/4 was significantly increased in apple NVs-treated osteoblastic MC3T3-E1 cells on day 3, however, this mRNA expression was not changed on day 7 (Fig. 5A). On day 3, the apple NVs significantly elevated the transcript- and protein-level expression of Smad1 in osteoblastic MC3T3-E1 cells in a dose-dependent manner (Fig. 5A).
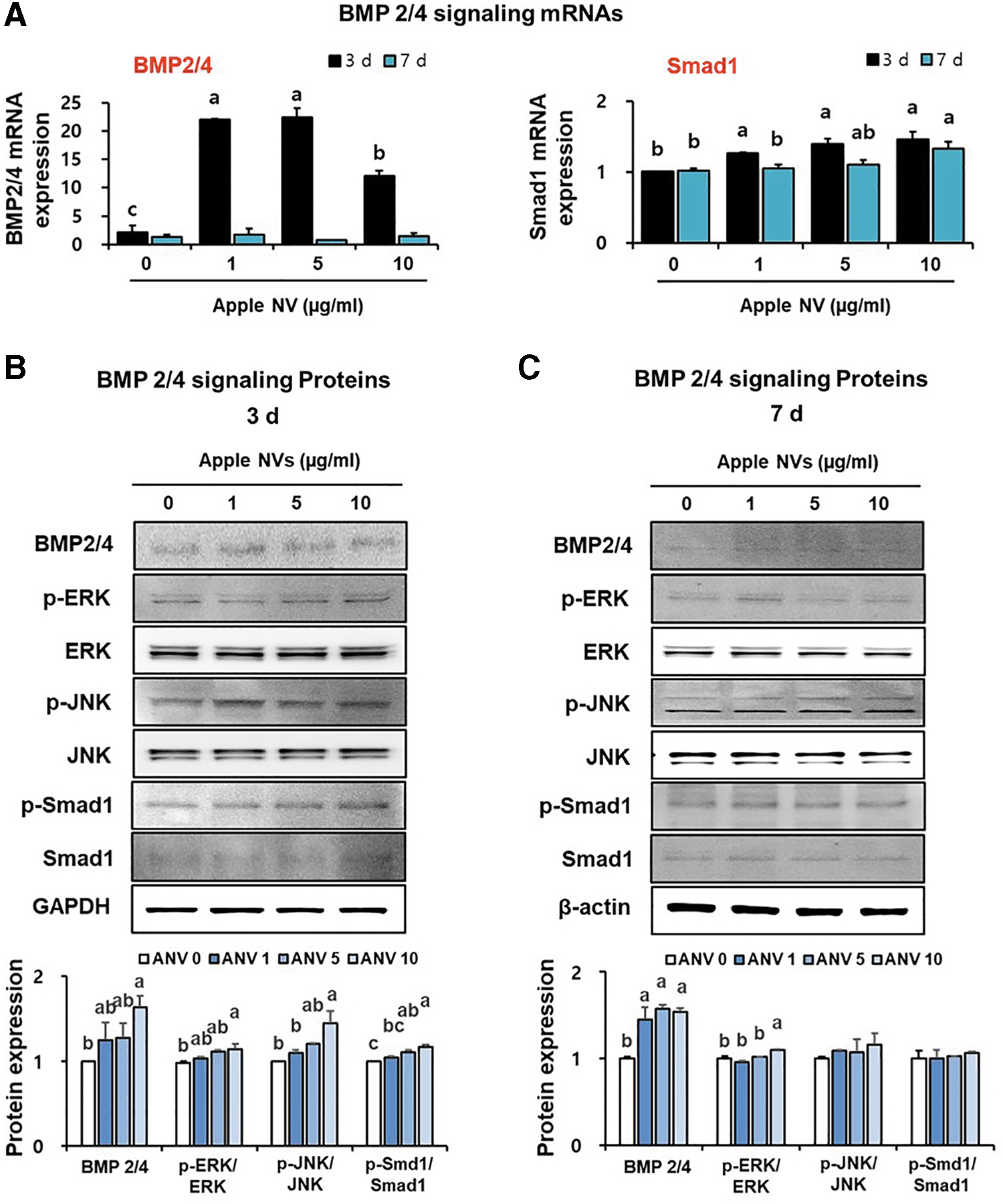
Activation of BMP 2/4 signaling proteins after treating osteoblastic MC3T3-E1 cells with apple NVs.
Whether the mitogen-activated protein kinase (MAPK) cascade contributes to the acquisition of osteoblast markers through BMP-2 was also investigated (Fig. 5B, C). Interestingly, two main MAPK cascades, that is, ERK1/2 and JNK1/2, were significantly activated in apple NV-treated group (10 μg/mL), compared to those in the control group on day 3 in response to BMP 2/4 (Fig. 5B). Taken together, apple NVs were demonstrated to stimulate osteoblast differentiation through the BMP-2/Smad1 and BMP-2/MAPK signaling pathways, which upregulate downstream osteoblast marker genes.
DISCUSSION
Bone tissue undergoes a continuous process of remodeling or bone turnover to maintain normal skeletal structure and function, 31 which depend on many types of cells and molecular factors. Osteoblasts and osteoclasts are two primary cell types involved in these processes. 32 Osteoclasts are responsible for aged bone resorption, while osteoblasts are in charge of new bone formation. 33 Bone resorption and formation are well-balanced under normal physiological conditions, 34 however, the disruption of bone homeostasis results in induced osteoporosis and other bone diseases. 35 Many herbal medicines and natural plant compounds that increase bone formation have recently attracted much attention 9,10 for the prevention and treatment of osteoporosis, which is a result of higher rates of bone resorption than formation. 5 –8
Once bone-matrix synthesis is initiated in osteoblast culture models, such as osteoblastic MC3T3-E1 cells derived from the calvaria of newborn mice, 36 cells can differentiate to osteoblasts with increased expression of the transcripts and proteins associated with osteoblast markers. The transcription factor Runx2 is a critical regulator of the maturation and stabilization of osteoblasts, with increased ALP activity, expression of BMP genes, and mineralization in immature mesenchymal and osteoblastic cells. 37,38 ALP, a glycoprotein enzyme, is an early marker of osteogenic differentiation from preosteoblasts to mature osteoblasts, by stimulating the differentiation and maturation of osteoblasts. 39,40 OPN, which appears during early proliferation and later differentiation, is associated with osteoblast maturation between adhesion before mineralization and matrix synthesis. 41 Mature osteoblasts are characterized by the production of ALP, alteration of procollagen to collagen, and the deposition of extracellular matrix calcium, all of which lead to subsequent mineralization of bones. 40,41
In this study, we investigated the beneficial effects of apple NVs on bone formation and underlying mechanisms. Specifically, stimulatory effects of exogenously administered apple NVs on the proliferation and differentiation of osteoblasts were evaluated using the osteoblastic MC3T3-E1 cells as an in vitro model of osteoblast differentiation. Cells treated with apple NVs exhibited higher ALP staining signals than untreated cells at 7 days, and this result was consistent with the elevated ALP activity (Fig. 2B–D). As increased collagen synthesis and extracellular matrix mineralization stimulate the maturation of the bone matrix and osteoblasts, the effects of apple NVs on collagen synthesis in the cell matrix and the mineralization of the extracellular matrix were examined in osteoblastic MC3T3-E1 cells. Collagen synthesis and the production of mineralized nodules were increased in the apple NV-treated group (Fig. 2D). These results indicate that apple NVs promote the differentiation and mineralization of osteoblastic MC3T3-E1 cells.
In addition, treatment with apple NVs significantly increased the expression of bone-related genes and proteins (Runx2, ALP, and OPN) in osteoblastic MC3T3-E1 cells. These findings are in agreement with the results of a recent study, 6 which reported that the dihydrochalcone phlorizin and phloretin in apple peels stimulated the proliferation and osteogenesis of bone marrow stromal cells. Overall, our findings demonstrate the osteogenic functions of apple NVs in osteoblastic MC3T3-E1 cells via increased osteoblast differentiation and mineralization.
Underlying mechanisms of the stimulatory effect of apple NVs on bone-related gene and protein synthesis were also investigated. Osteogenic signaling pathways, such as the bone morphogenetic protein 2 (BMP-2) and Wnt/β-catenin pathways, play an important role in regulating bone metabolism. BMPs represent a unique group of proteins within the transforming growth factor-beta (TGF-β) superfamily of genes, which play a pivotal role in the regulation of heart, nerve, cartilage, and bone development. 42,43 Among BMP family proteins, BMP-2 strongly induces bone formation by binding to two receptors belonging to subfamilies of transmembrane serine/threonine kinases (the type-I and type-II receptors). 43 –45 Smad family proteins have been identified as the downstream propagators of the BMP signals. 43 –45 Activation of the BMP-2/Smad (Smad1/5/8) signaling pathway increases the mRNA transcripts of the osteogenic transcription factor Runx2, which further promotes the expression of bone-related genes, including ALP, OPN, OCN, and type-I collagen. 43 –45
In addition to the proteins of the Smad family, the activation of the MAPK cascade is also involved in TGF-β superfamily-mediated signal transduction. 46 BMP-2 activates the BMP-2/MAPK signaling pathway by binding to the type-I and type-II BMP receptors, transmitting the signal to the nucleus through extracellular signal-regulated protein kinase (ERK 1/2), c-Jun N-terminal kinase (JNK 1, 2/3), and p38. A previous study by Jaiswal et al reported that these three MAPK signaling pathways are specifically involved in the osteogenic differentiation of stem cells. 47
In our study, the transcript- and protein-level expressions of BMP-2 and Smad1 were significantly increased in osteoblastic MC3T3-E1 cells after treatment with apple NVs (Fig. 5A, B). These results suggest that the BMP-2/Smad1 signaling pathway plays a critical role in the apple NV-mediated maturation and differentiation of preosteoblasts. Furthermore, the effects of apple NVs on other vital downstream propagators of BMP signaling, such as the MAPK signaling pathway, were analyzed (Fig. 5); the activations of two of the main MAPK cascades, that is, ERK1/2 and JNK1/2, were significantly increased in response to BMP 2/4 in a dose-dependent manner in apple NVs-treated osteoblastic cells.
Apples contain various polyphenolic compounds, such as catechin, epicatechin, phloridzin, phloretin, procyanidin, chlorogenic acid, hydroxycinnamates, rutin, and quercetin. 15,16 Polyphenols have been reported to have a variety of biological activities, including antioxidant, anti-inflammatory, and antitumor effects. 15 Recently, phenolic compounds have been reported to protect the bone by enhancing osteoblastogenesis and reducing osteoclastogenesis via several mechanisms involving antioxidants and osteoimmunological activity. 48 However, despite this study, 48 further mechanistic studies are required to elucidate the potential participation of polyphenols in the osteogenic functions and mechanisms of apple NVs.
In conclusion, apple NVs stimulate ERK1/2, JNK1/2, and Smad1 signaling through the activation of BMP-2, which continuously increase bone formation by promoting preosteoblast cell growth, differentiation, and mineralization. Therefore, apple NVs serve as potential antiosteoporosis agents. However, further in vivo studies are needed to fully demonstrate the therapeutic potential of apple NVs in promoting bone health.
Footnotes
AUTHORs' CONTRIBUTIONS
Y.J.S., H.J.S., and Y.E.C. designed the study. Y.J.S., H.J.S., D.H.K., S.H.L., and J.H.K. performed the experiments, analyzed data, made the figures, and performed the statistical analysis. I.S.K., C.E.J., J.I.K., J.H.L., D.K.K., M.C.B., and Y.E.C. drafted the initial version of this article, and then all authors contributed to the editing of the article. All authors have read and approved the final version of this article.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This work was supported by National Research Foundation of Korea (NRF) grant funded by Korea government (MSIT) (No.2021R1C1C1008117, 2021R1A5A2021614, 2022R1C1C100633411). This work was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2018R1A6A1A03024862). Following are results of a study on the “Leaders in Industry-university Cooperation (LINC)” Project, supported by the Ministry of Education and the National Research Foundation of Korea (NRF).