
Abstract
Obesity is currently regarded as a global concern, and the key objectives of the global health strategy include its prevention and control. Probiotic supplementation can help achieve these objectives. This study aimed to assess whether a probiotic strain Lactobacillus paracasei ssp. paracasei, Lactobacillus casei 431 (henceforth, L. casei 431) possesses antiobesogenic properties. High-fat diet-induced obese Sprague-Dawley rats were treated with L. casei 431 for 10 weeks, and the outcomes were compared with those of rats treated with the antiobesity medication orlistat. Body weights, epididymal fat, and tissues from mice were assessed. Furthermore, serological and histological analyses were performed. Epididymal fat accumulation was significantly reduced in groups administered L. casei 431 and orlistat. Furthermore, L. casei 431 and orlistat treatments lowered serum alanine transaminase, aspartate aminotransferase, and triglyceride (TG) levels. Hematoxylin and eosin staining of the liver and epididymal adipose tissues showed that the L. casei 431-treated groups exhibited reduced lipid buildup and adipocyte size. Furthermore, sterol regulatory element-binding protein 1c, adipose TG lipase, and lipoprotein lipase messenger RNA (mRNA) levels were upregulated, leading to lipid oxidation and degradation, in L. casei 431-supplemented groups. Furthermore, carnitine palmitoyltransferase 1, a major factor in lipolysis, was consistently upregulated at the protein level after L. casei 431 administration. Collectively, these results demonstrate the potential of L. casei 431 in alleviating obesity in rats through optimizing lipid metabolism and some related biomarkers.
INTRODUCTION
Overweight and obesity are multifactorial metabolic disorders in which energy intake from diet is greater than energy expenditure, resulting in abnormal fat accumulation and adverse health effects. Individuals with obesity are more susceptible to numerous metabolic diseases, including diabetes mellitus, nonalcoholic fatty liver disease, and dyslipidemia, all of which are risk factors for cardiovascular diseases. 1,2
Obesity management encompasses lifestyle changes, regular physical activity, and even surgical procedures, along with medications that alter appetite, affect lipid utilization, or reduce nutrient absorption. Orlistat, for example, is a Food and Drug Administration (FDA) 3 -approved drug for long-term management of obesity. Nevertheless, numerous antiobesity medications as well as bariatric procedures are high priced and associated with adverse reactions. 4 Thus, preventing and managing obesity through nonpharmacological options with minimal side effects appear safer and appropriate. 5
Several human and animal studies have shown that probiotics are promising nonpharmacological alternatives for addressing diverse disorders. 6 Probiotics are viable microorganisms available in foods or dietary supplements that beneficially affect one's health when provided in adequate quantities, 7 and they may act through a number of mechanisms. They promote intestinal health by aiding food digestion and inhibiting pathogenic bacteria through competive exclusion or release of antibacterial metabolites. 8 They also help maintain immune homeostasis by modulating inflammatory responses and gut-related gene expression 9 as well as by exerting antioxidant activities. 10
Probiotics with antiobesity potential have been shown to reduce adipocyte size along with body and fat pad weight, 11 restore lipid and hepatic profiles, 12 and enhance overall metabolic status. 10 There are several types of probiotics, with Lactobacillus being one of the most common. 6,13 Many beneficial effects of Lactobacilli strains, such as improving high-fat diet (HFD)-induced hyperlipidemia and obesity 14 and attenuating the size and load of adipose tissue, 15 have been observed in animal models. Lactobacillus paracasei CNCM I-4270 reportedly decreases weight in mice, even when food intake has not decreased. 16 In addition, more antiobesity effects were observed in clinical trials upon administration of Lactobacillus strains. 6,17
Jones 18 emphasized the positive effects of Lactobacillus casei and L. paracasei (classified into two subspecies comprising subsp. tolerans and subsp. paracasei) strains in clinical studies of obesity, dyslipidemia, and other chronic conditions. Launched by Chr. Hansen A/S (Hørsholm, Denmark), L. paracasei subsp. paracasei. L. casei 431 (henceforth, L. casei 431) was originally isolated from an infant stool and has been used since 1995 as a dietary supplement worldwide, with many related safety and survivability studies. 19 It has also been approved by the U.S. FDA and by the European Food Safety Authority. Moreover, Rizzardini et al. 20 and Jespersen et al. 21 revealed the beneficial effects of L. casei 431 on immune function. However, its antiobesity effects are not yet clearly known.
This study aimed to investigate the antiobesity effects of L. casei 431 on body weight, fat mass distribution, fat accumulation in the liver or adipose tissues; messenger RNA (mRNA) expression; and protein expression in HFD-induced obese rats.
MATERIALS AND METHODS
Animals and experimental design
Male Sprague-Dawley rats (6-week-old) were purchased from Orientbio (Gapyeong, South Korea). They were maintained under constant conditions (under a 12 h light/dark cycle, at a humidity of 50% ± 10% and temperature of 23°C ± 2°C), with food and water supplied ad libitum. All experiments were conducted according to a protocol approved by the institutional animal care and use committee of the Food Industry Promotional Agency of Korea (IACUC-20-004).
L. casei 431 was produced by Chr. Hansens (Hoersholm, Denmark) and supplied in a freeze-dried form by VIXXOL Corporation (Gunpo, South Korea). It was diluted in phosphate buffered saline (PBS) for administration.
After acclimatization for a week, animals received experimental treatment for 10 weeks. Rats in the control group (ND) were fed a normal diet (Table 1) (Lab Diet No. 5053; PMI Nutrition International) and orally treated with PBS. The remaining rats were fed a HFD (Table 1) (D12492; Research Diet) and were randomly assigned to four groups, including the HFD group wherein PBS was orally administered, the L. casei 431 (L) group (108 colony forming unit [CFU]/kg per day of L. casei 431 by oral administration), the L. casei 431 (H) group (1010 CFU/kg per day of L. casei 431 orally administered), and the orlistat group (30 mg/kg of orlistat orally administered).
Nutritional Composition of the Experimental Diet
Individual magnetic resonance imaging (MRI) scans using nuclear magnetic resonance (EchoMRI TM-700; Echo-MRI) were performed every 2 weeks to monitor fat deposits, whereas body weight and feed intake were recorded on a regular basis. Before necropsy, final fat distribution in rats was measured using dual-energy X-ray absorptiometry (DXA; iNSiGHT; Osteosys), which has a high accuracy in portraying body composition. 22,23 Blood, epididymal fat, and liver were collected, and stored for further analyses.
Serological analysis
Blood samples were obtained from the vena cava after CO2 anesthesia. Serum samples were stored at 20–25°C for 30 min and centrifuged at 13,572 × g for 10 min. All samples were analyzed in terms of the lipid profile indices including levels of total cholesterol (TC) and triglyceride (TG), and liver function test including levels of aspartate aminotransferase (AST) and alanine aminotransferase (ALT) using a clinical chemistry analyzer (TBA120FR; Toshiba Ltd.).
Histological analysis
Samples of epididymal fat and liver were dissected, rinsed with normal saline, fixed in 10% neutral formalin solution, trimmed, processed, embedded in paraffin, and cut into measuring 5 μm sections. Tissues were stained with hematoxylin and eosin (H&E) (Sigma-Aldrich) and an image was obtained using a slide scanner (Leica) at × 200 magnification. The adipocyte mean areas as well as lipid accumulation in the liver were established using Image-J software (National Institutes of Health, Bethesda, MD, USA).
Real-time polymerase chain reaction
Total RNA was extracted from liver tissues using Trizol reagent. A complementary DNA (cDNA) library was synthesized using a Prime-Script™ reverse transcription reagent kit (TakaRa, Shiga, Japan). mRNA expression levels were measured by analyzing cDNA using an iCycler iQ™ Real-Time PCR (polymerase chain reaction; qPCR) Detection System (Bio-Rad Laboratories, Hercules, CA, USA) with SYBR Green (TOYOBO, Osaka, Japan) and specific rat primers (Table 2).
Primer Sequences for Real-Time Polymerase Chain Reaction
ATGL, adipose triglyceride lipase; LPL, lipoprotein lipase; SREBP-1c, sterol regulatory element-binding protein 1c.
Hepatic protein contents
To identify proteins involved within the hepatic tissue, samples were homogenized in RIPA buffer, centrifuged, and the supernatants were kept for subsequent total protein normalization by Bradford method. 24 Then, carnitine palmitoyltransferase 1 (CPT-1; MyBioSource) and adiponectin (Thermo) were measured using an enzyme-linked immunosorbent assay (ELISA) kit according to the manufacturer's instructions.
Statistical analysis
Changes in body weight, fat weight, and feed intake measurements over time were assessed by two-way repeated measures analysis of variance (ANOVA) with Dunnett's post hoc adjustments. Analyses of qPCR data were performed using Bio-Rad CFX maestro software (ver. 4.0; Bio-Rad Laboratories). The analyses were performed based on the groups (ND, HFD,
Gene expression stability of sterol regulatory element-binding protein 1c (SREBP-1c), adipose triglyceride lipase (ATGL), and lipoprotein lipase (LPL) was analyzed using the geNorm algorithm, included with the CFX maestro software (ver. 4.1; Bio-Rad Laboratories). The expression ratio of the genes of interest was normalized relative to the abundance of the reference gene using the ΔΔCq method. Expression values are presented as means of fold change with the standard error using CFX maestro software (ver. 4.0; Bio-Rad Laboratories).
All data are expressed as the means and standard errors of the means. Statistical evaluation was performed using one- or two-way ANOVA followed by Dunnett's post hoc adjustments. All analyses were performed using GraphPad Prism version 9.3.1 (GraphPad Software). P < .05 was determined to be significant.
RESULTS
L. casei 431 moderated fat mass gain in HFD-induced obese rats
HFD led to a significant increase in body weight over the 10-week period. The body weights of all groups increased weekly, particularly in the HFD-fed groups compared with the ND group (Fig. 1A). However, body weights of the L. casei 431-treated groups and the orlistat group were reduced throughout the experimental period, although the change was not statistically significant. At the end of the experiment, the gain in body weight was significantly higher in the HFD group than in the ND group, whereas L. casei 431 and orlistat treatments showed a tendency to lower the body weight gain (Fig. 1B).
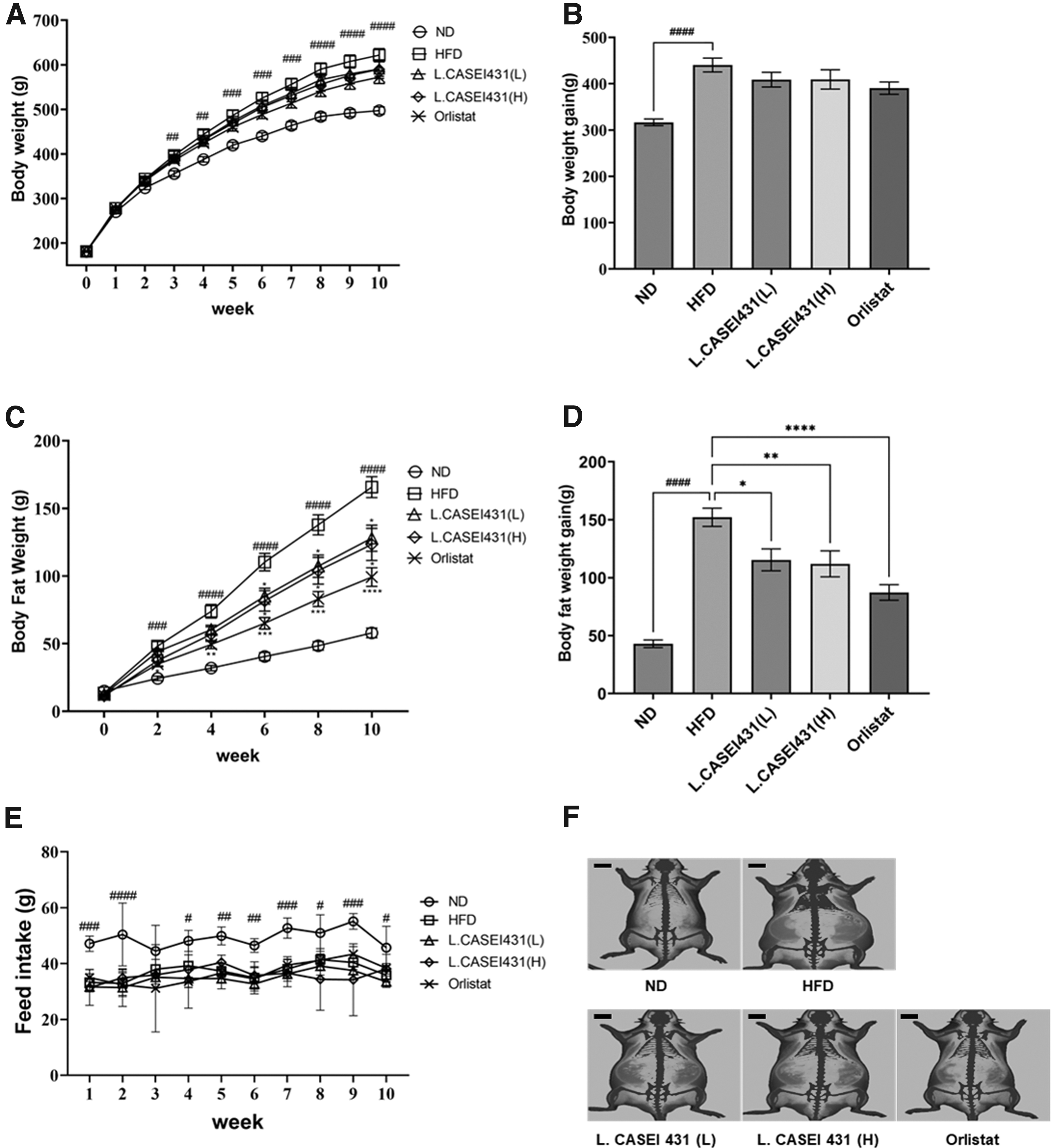
Effects of L. casei 431 supplementation on anthropometric parameters and food intake in the experimental rats. L. casei 431 (L) (108 CFU/kg per day) or L. casei 431 (H) (1010 CFU/kg per day) or orlistat (30 mg/kg) was orally administered to experimental rats fed a high-fat diet for 10 weeks.
As indicated in Figure 1C, the fat weight of the HFD group was significantly higher than that of the ND group starting from the second week of the experiment; in L. casei 431- and orlistat-treated groups, a tendency of suppression of the increase in fat depot was observed. The difference in fat weight between the HFD group and the orlistat group became noticeable after the fourth week and that of the L. casei 431-treated group became significant after the sixth week. Figure 1D shows significantly lowered fat gain values in L. casei 431 and orlistat groups compared with those in the HFD group.
Although food intake differed significantly between the ND- and HFD-fed groups, the difference was not significant (Fig. 1E). As shown in Figure 1F, measurement of the fat density distribution (red), using DXA, showed a highly distributed fat depot in the HFD group. Conversely, L. casei 431- and the orlistat-treated groups showed relatively low fat distribution.
L. casei 431 exerts an effect on liver and epididymal fat histopathology
As shown in Figure 2A and B, liver histological analyses revealed severe accumulation of adipose tissue around the central vein in the HFD group compared with that in the ND group. Moreover, numerous lipid droplets appeared in the cytoplasm of the hepatocytes. However, this ectopic fat accumulation was attenuated in all the supplemented groups especially the L. casei 431 (H) and orlistat groups. Interestingly, no significant differences were noted among the liver weights at the end of the experiment, although an increasing tendency was observe in the liver weight of the HFD group (Fig. 2C).
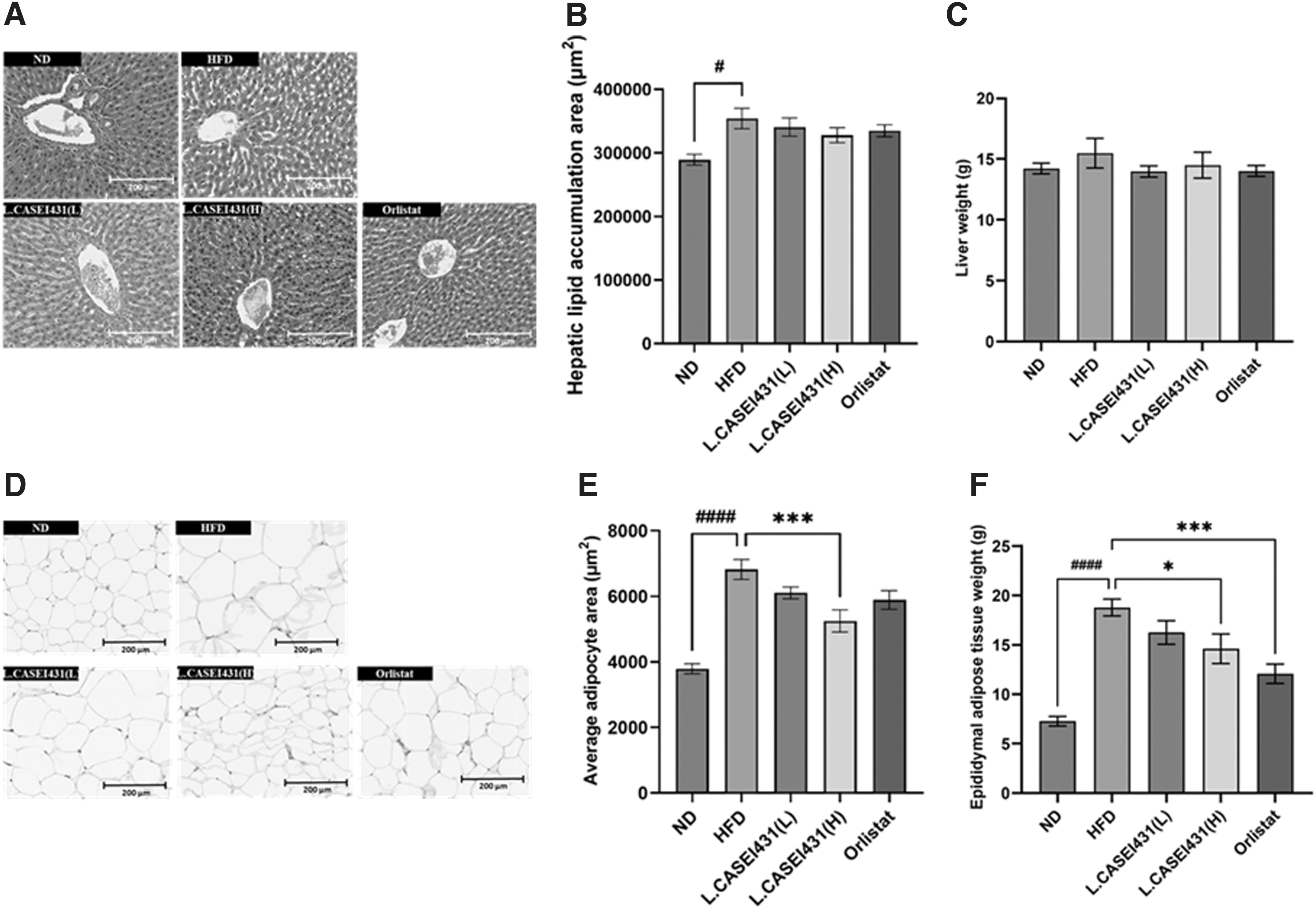
Effect of L. casei 431 supplementation observed by H&E staining in liver and epididymal adipose tissue in the experimental rats. L. casei 431 (L) (108 CFU/kg per day) or L. casei 431 (H) (1010 CFU/kg per day) or orlistat (30 mg/kg) was orally administered to experimental rats fed a high-fat diet for 10 weeks. Histological sections were obtained from the liver tissue and epididymal white adipose tissues, and stained with H&E.
Histological measurement of fat index in the epididymal adipose tissue showed that epididymal adipocytes in the HFD group were irregular and larger than those in the ND group. However, these changes were not apparent in the other groups, especially in the L. casei 431 (H) and orlistat groups (Fig. 2D). In addition, adipocyte size in the HFD group notably increased compared with that in the ND group, but this adipocyte hypertrophy was significantly reversed after supplementation with a high dosage of L. casei 431 (Fig. 2E).
Upon completion of the experimental treatments, the weights of epididymal white adipose tissue were dramatically high in the HFD group. Contrastingly, orlistat and L. casei 431 (H) treatments suppressed epididymal adipose tissue accumulation when compared with those in the HFD group (Fig. 2F).
L. casei 431 improves serological parameters
The HFD group had higher serum TG, TC, ALT, and erase (AST) levels than the ND group. After 10 weeks of supplementation, ALT levels in the plasma of the L. casei 431- and orlistat-treated groups were significantly reduced in a dose-dependent manner (Fig. 3A), whereas AST concentrations were markedly diminished only in the L. casei 431 (H)- and orlistat-treated groups (Fig. 3B). The HFD group showed the highest concentrations of TC and TG. Interestingly, the L. casei 431 (H) treatment led to a significant reduction in TG levels, whereas orlistat treatment markedly reduced TC levels (Fig. 3C, D).
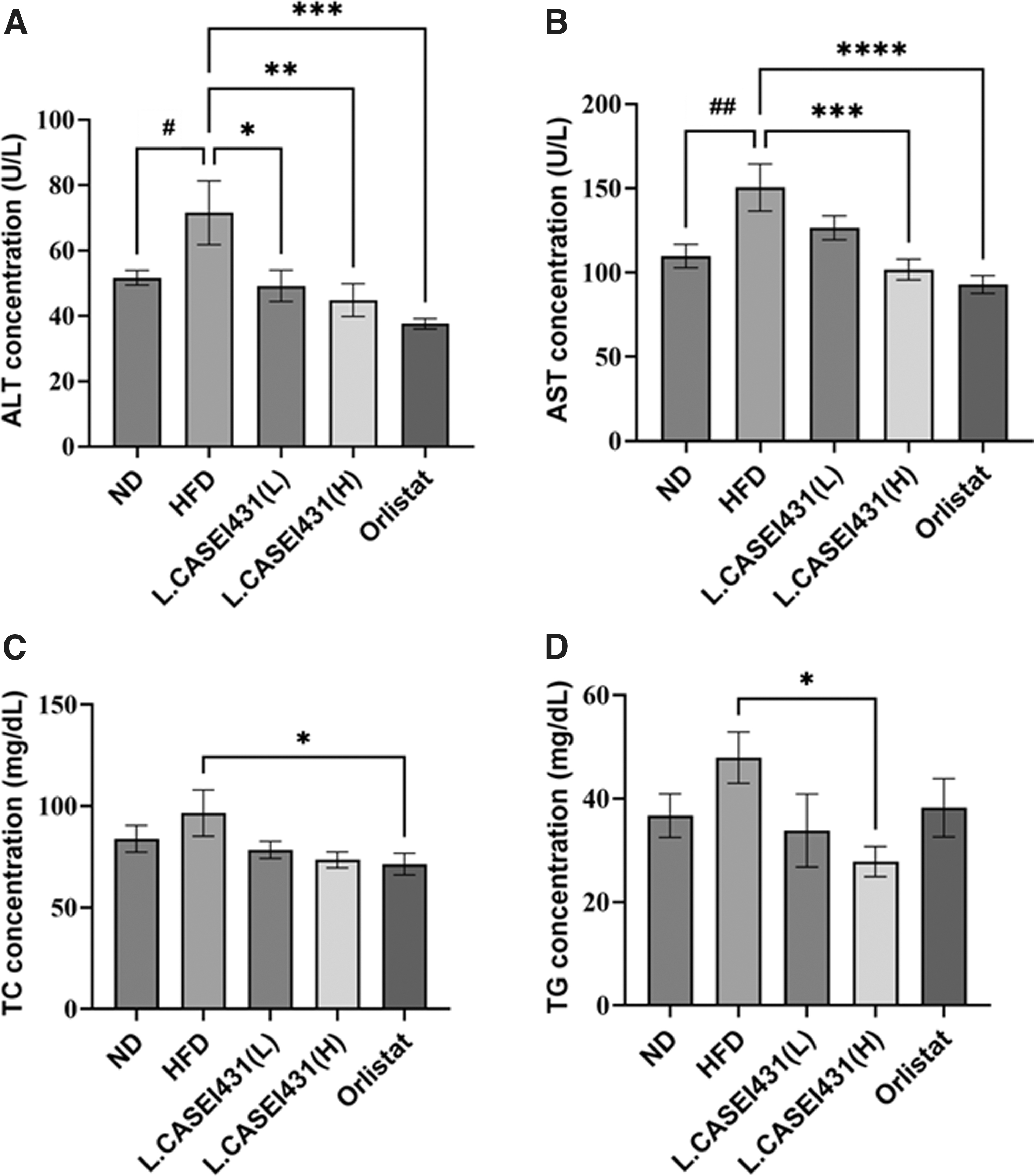
Effect of L. casei 431 on blood chemistry in the experimental rats. L. casei 431 (L) (108 CFU/kg per day) or L. casei 431 (H) (1010 CFU/kg per day) or orlistat (30 mg/kg) was orally administered to experimental rats fed a high-fat diet for 10 weeks. High concentration of L. casei 431 significantly increased blood triglyceride levels.
Effect of L. casei 431 on lipid synthesis-related gene factors in the liver
To understand the mechanism of the antiobesity effect of L. casei 431, qPCR analysis of mRNA expression in the liver was performed. As shown in Figure 4A, mRNA expression levels of SREBP-1c, which is related to lipogenesis in the HFD group, were significantly higher than those in the ND group. However, L. casei 431 and orlistat treatment significantly downregulated the mRNA expression levels to near-normal levels.

mRNA expression of hepatic SREBP-1c, ATGL, and LPL in the experimental rats. L. casei 431 (L) (108 CFU/kg per day) or L. casei 431 (H) (1010 CFU/kg per day) or orlistat (30 mg/kg) was orally administered to experimental rats fed a high-fat diet for 10 weeks.
Effect of L. casei 431 on lipid hydrolysis-related gene expression in the liver
To gain more understanding of the effects of L. casei 431 against fat accumulation, we analyzed the expression of lipid hydrolysis genes in the liver. Figure 4B indicates that the mRNA expression of ATGL related to lipolysis in the L. casei 431 (H) group showed a significant increase when compared with that of the HFD group. In addition, the expression of LPL significantly increased in the L. casei 431 (H) group (Fig. 4C).
Effect of L. casei 431 on lipid oxidation-related protein factors in the liver
To gain more insights into the antiobesity effect of L. casei 431, fatty acid oxidation-related genes, such as adiponectin and CPT-1, in hepatic tissue were examined by ELISA. As indicated in Figure 5A, CPT-1 in the protein levels in the L. casei 431 (H) and orlistat groups was significantly higher than that in the HFD group. Furthermore, adiponectin showed an increasing tendency in the L. casei 431 (H)-treated group (Fig. 5B).
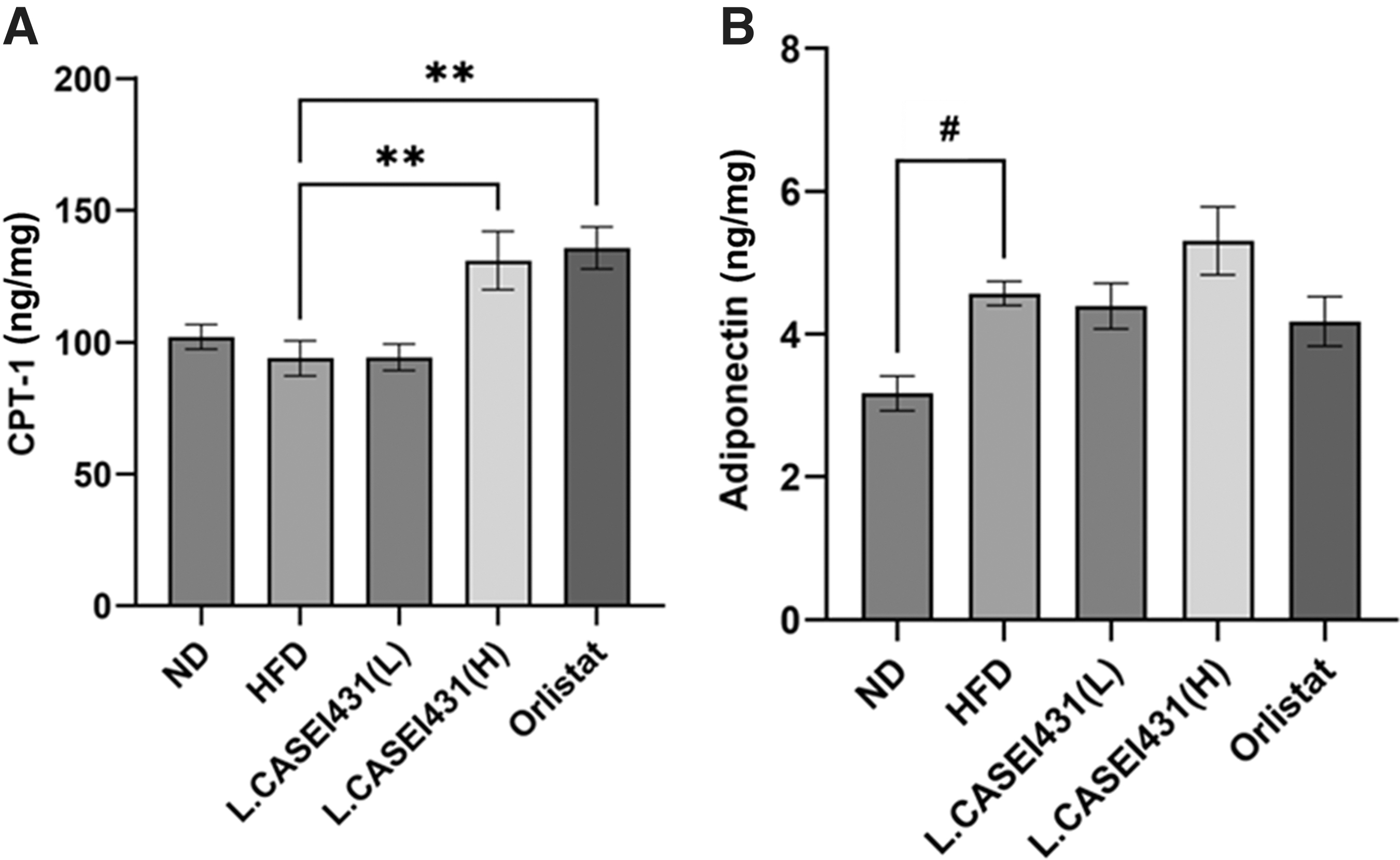
Effect of L. casei 431 on hepatic CPT-1 and adiponectin in the experimental rats examined by ELISA. L. casei 431 (L) (108 CFU/kg per day) or L. casei 431 (H) (1010 CFU/kg per day) or orlistat (30 mg/kg) was orally administered to experimental rats fed a high-fat diet for 10 weeks. Protein expression levels of
DISCUSSION
Incorporation of probiotics into the dietary strategies for managing obesity has lately emerged as a safe treatment, 25 as most probiotics are naturally present within the gut microbiota. The effects of probiotics on obese models include an improvement in morphological characteristics and obesity-related metabolic shifts along with an increase in metabolic and inflammatory biomarkers. 26 The majority of current research focuses on using Lactobacillus spp. to treat and prevent obesity. 6 However, basic research on probiotics showed that each strain must be considered on its own merits as outcomes from specific strains may not be shared by even strains of the same species. 27,28
Herein, L. casei 431 was administered for 10 weeks, and the results were compared with those of the antiobesity medication orlistat in HFD-fed rats. Successful alleviation of obesity was observed after examination of fat depot weight, epididymal fat weight, and markers in the liver and epididymal adipose tissue, despite no significant differences in food intake. In addition, a decrease in serum lipids was observed; particularly, TG levels in the L. casei 431 group were lower than those in the orlistat group.
Long-term HFD consumption increases body weight and fat mass. 29 In this study, HFD rats showed the greatest weight gain compared with ND rats, demonstrating that obesity was effectively induced. Although body weight loss is regarded as the most significant goal in obesity interventions, fat mass loss is a more specific and health-promoting goal. Willoughby et al. 30 suggested that dietary interventions that inhibit fat mass accumulation while preserving lean body mass may be metabolically beneficial. Herein, the body weight statuses in all HFD-fed groups were not statistically different, but the L. casei 431- and orlistat-treated groups exhibited significantly low fat mass compared with the HFD group, as shown by echo-MRI.
In contrast, a high dose of L. casei 431 markedly reduced the weights of epididymal fat. Notwithstanding the fact that only epididymal fat was examined in this study—a limitation we acknowledge—the fat distribution was verified using echo-MRI and DXA, which are accurate and precise techniques applied in both humans 31,32 and animals. 22,23 Interestingly, the HFD group and the L. casei 431 supplemented groups did not differ in the feed intake, suggesting that body weight reduction was induced by L. casei 431 supplementation. Afterward, we speculate that L. casei 431 administration somehow contributed to a reduction in fat accumulation before body weight loss in accordance with a previous study on obese rats treated with a Lactobacillus probiotic strain. 15
Excessive calories result in fat accumulation in ectopic tissues, including skeletal muscle, liver, and heart tissue, which is a risk factor for obesity-related metabolic disorders. 33 In this study, ectopic lipid deposition in the liver was markedly pronounced in the HFD group compared with that in the ND group. Similar to a previous study on HFD-induced obese rats treated with bacterial strains, 34,35 no significant increase was observed in the liver weight. Furthermore, notably, adipose hypertrophy was suppressed after L. casei 431 (H) and orlistat treatments.
Serological biomarkers of liver injury, such as ALT levels, were markedly downregulated in all the supplemented groups, whereas AST levels were only downgraded by L. casei 431 (H) and orlistat. In brief, it can be suggested that L. casei 431 (H) and orlistat can effectively prevent hepatic steatosis.
Dyslipidemia, observed in around 60–70% of obese subjects, 36 refers to abnormal levels of blood fats, including but not limited to serum TG and TC. The findings of this study showed an improved serum lipids profile after treatment with L. casei 431. In fact, TG levels were sensitively reduced by L. casei 431 (H) treatment, whereas TC levels were sensitively reduced in the orlistat group compared with those in the HFD group. To gain more insights into the effect of L. casei 431 on attenuation of fat accumulation, the expression levels of genes and proteins related to obesity and energy homeostasis were analyzed.
Several mechanisms are involved in alleviating obesity, among which the inhibition of lipid synthesis and the increase in lipid oxidation were first considered. SREBPs are transcription factors that regulate the expression of genes that affect cholesterol and fatty acid production and are implicated in adipogenesis. 37 Overexpression of nuclear SREBP-1c in adipose tissue reportedly causes lipodystrophy, insulin resistance, and type 2 diabetes. 38,39 Herein, L. casei 431 downregulated SREBP-1c mRNA levels in the liver and thus, enhanced suppression or disruption of the lipogenic pathway.
CPT-1 and adiponectin, known to affect lipid oxidation, together with genes encoding lipid hydrolysis factors, such as LPL and AGTL, were examined in liver tissue. ATGL is a rate-limiting enzyme for lipolysis in adipocytes that operates by eliminating free fatty acids. ATGL also initiates lipolysis by removing free fatty acids to form a diacylglycerol. 40 LPL, in contrast, is among the principal enzymes for TG lipolysis that is highly expressed in tissues that demand substantial amounts of free fatty acid.
Thus, LPL can increase the oxidation of fat by highly expressing it in tissues that require high levels of free fatty acids, such as the heart, skeletal muscle, and adipose tissue. 41 These findings were consistent with those of this study as L. casei 431 decreased fat accumulation by increasing the expression of fat synthesis inhibition-, decomposition-, and oxidation-related factors, especially owing to the activation of lipolysis. CPT-1, a rate-limiting enzyme in mitochondrial fatty acid oxidation, was also significantly increased in the L. casei 431 (H) and orlistat groups.
Furthermore, adiponectin, which enhances hepatic insulin action and suppresses fatty acid influx into the liver, 42 and thus, is a hormone that increases fatty acid oxidation, increased in the L. casei 431 (H) group. Interestingly, increased levels of adiponectin are reportedly associated with fat weight reduction. 43
The mechanism by which L. casei 431 acts to improve some aspects of obesity remains unclear. Further experiments before randomized clinical trials are essential to gain an in-depth understanding of the mechanisms of action of L. casei 431, with a focus on its effects in counteracting obesity. In our study, the effects of L. casei 431 were comparable with those of orlistat, which was used as the positive control. This indicates that L. casei 431 has weight gain suppressing and hypolipidemic abilities that are required for prevention or treatment of obesity.
Footnotes
AUTHORs' CONTRIBUTIONS
Y.J.S., J.-M.B. and M.J.B. conceived and designed the study, Y.J.S. and H.-R.C. performed the experiments and analyzed the data, Y.J.S. and P.M. wrote and edited the article, and H.H.K., S.H.K. and T.H.K. contributed to investigation.
AUTHOR DISCLOSURE STATEMENT
The authors have no potential conflicts of interest to disclose.
FUNDING INFORMATION
This research was supported by a grant from the VIXXOL Corporation.