
Abstract
Gulf War Illness (GWI) afflicts US military personnel who served in the Persian Gulf War. Suspect causal agents include exposure to pyridostigmine (PB), permethrin (PM) and N,N-diethyl-m-toluamide (DEET). Prominent symptoms include cognitive deficits, such as memory impairment. In aging animal models, we have documented the beneficial effect of the flavanol (-)-epicatechin (Epi) on hippocampus structure and related function. Using a rat model of GWI, we examined the effects of Epi on hippocampus inflammation, oxidative stress, mitochondrial dysfunction, cell death/survival pathways, and memory endpoints. Male Wistar rats underwent 3 weeks of exposure to either vehicles or DEET, PM, PB, and stress. Subgroups of GWI rats were then allocated to receive orally 15 days of either water (vehicle) or 1 mg/kg/day of Epi treatment. Object recognition tasks were performed to assess memory. Hippocampus samples were analyzed. Epi treatment yields significant improvements in short- and long-term memory versus GWI rats. Hippocampus oxidative stress and pro-inflammatory cytokine levels showed significant increases with GWI that were largely normalized with Epi becoming comparable to controls. Significant increases in markers of hippocampus neuroinflammation and cell death were noted with GWI and were also largely reduced with Epi. Neuronal survival signaling pathways were adversely impacted by GWI and were partially or fully restored by Epi. Markers of mitochondrial function were adversely impacted by GWI and were fully restored by Epi. In conclusion, in an animal model of GWI, Epi beneficially impacts recognized markers of hippocampus neuroinflammation, oxidative stress, cell survival, neurotoxicity and mitochondrial function leading to improved memory.
INTRODUCTION
Approximately 30% of the 700,000 US Veterans of the 1990–91 Persian Gulf War (GW) developed a persistent multisystem syndrome termed Gulf War illness (GWI). GWI comprises a gathering of symptoms that prominently affect the gastrointestinal, nervous, and skeletal muscle systems leading to cognitive deficits, headaches, sleep disturbances, exercise intolerance, and fatigue. 1,2 Most common neurological abnormalities include depression, anxiety, and learning/memory impairments which have been linked to hippocampal dysfunction. 3 –6 GW Veterans continue to experience symptoms and altered function after ∼30 years. The precise etiology is unknown and several hypotheses have been proposed. Co-exposure to specific chemical agents and stress are suspect etiological factors. 2 Military personnel stationed in the battlefield are believed to have consumed daily the reversible acetylcholinesterase inhibitor (AChEi) pyridostigmine bromide (PB) pills as a prophylactic treatment to protect against nerve gas attacks. 1,2 To reduce the risk of infection by diseases transmitted by various vectors, personnel were also exposed to insecticides and insect repellants most commonly permethrin (PM) and N,N-dietyl-m-toluamide (DEET). PM intoxication results from the opening of voltage-gated sodium channels. 7 DEET’s target is unknown but, human poisoning can occur. 7 AChEi and organophosphate toxicity and lethality have been related to the development of oxidative stress (OS) and mitochondrial dysfunction (MD). 2,7,8 Long-term exposure to PB has also been associated with polyneuropathy, neuropsychiatric symptoms such as depression and mood changes, problems with attention, information processing, problem solving, and short-term memory. 9
Rodent models of a GWI-like illness have been developed and rely on the exposure of animals for several weeks to suspect chemical agents and stress. Behavioral studies in GWI rodent models have identified hippocampus-dependent learning and memory sites as targets of chemical exposure leading to diminished function. In brain, decreased neurogenesis, neuroinflammation, OS, and MD were noted. 10,11 Recently, direct in vivo evidence of neuroinflammation in GWI patients was reported. 12 With the aim of improving neurological symptoms, candidate therapeutic compounds have been tested in rodent models (e.g., luminol and curcumin) and select ones in pilot trials with GWI patients (Coenzyme Q10). 13 –15 These compounds appear to target OS, inflammation and/or MD and have yielded initial promising results.
Preclinical, clinical, and epidemiological evidence indicates that the consumption of cacao products can ameliorate age-related cognitive decline. A study in aged rats demonstrated positive effects of a flavonoid-rich cocoa extracts in discrimination and spatial tasks. 16 In normal aged human subjects, the consumption of flavanol rich cocoa for 12 weeks was associated with improvements in cognitive function and was linked to enhanced hippocampus function. 17 Epidemiological studies also associate the sustained consumption of high flavanol cocoa products with improved cognitive function. 18 The most abundant flavanol present in cacao is (-)-epicatechin (Epi), which stimulates nitric oxide (NO) production in endothelial cells and thus, increases blood flow. Epi can cross the blood-brain barrier and potentially directly act on neurons and supporting systems. 19 In mice, Epi favorably impacts anxiety, learning and memory. 20 –22 Such effects are associated with increased hippocampus angiogenesis and neuronal spine density as well as with the upregulation of mRNA levels of proteins associated with learning and downregulation of markers of neurodegeneration. 20 –22 We reported on the beneficial effects of Epi on OS levels and indicators of mitochondrial biogenesis, structure and function in multiple organs including the prefrontal cortex of aged mice. 23 Thus, Epi recapitulates the beneficial actions of cocoa in blood vessels and brain. In a rat model of GWI, we reported on the beneficial effects of two week treatment with Epi on multiple indicators of skeletal muscle structure and function. 24 However, no studies have examined in rodent models of GWI the effects of Epi on functional and structural indicators of brain health.
Therefore, the purpose of this study was to evaluate the effects of Epi treatment on short- and long-term memory in a rat model of GWI and in recognized markers of neuroinflammation, OS and MD in hippocampus.
MATERIALS AND METHODS
Study design and animal model
All experimental protocols were approved by University of California, San Diego’s IACUC (animal) committee, and all methods used were in accordance with Federal and institutional guidelines and regulations. The experimental design is summarized in Supplementary Figure S1. The GWI rat model is based on that developed by Hattiangady et al., with minor modifications. 10 Three month old, male Wistar rats (n = 14) underwent 3 weeks of exposure simultaneously to PB (oral gavage 1.3 mg/kg/day), PM and DEET (skin applications of 0.13 and 40 mg/kg/day respectively, in 70% ethanol) as well as physical restraint (stress) by placing the animals in a plexiglass holder for 5 min/day. Control animals (n = 14) were given only vehicles and no restraint. Animals were then allowed to recover for 3 weeks. During the last 2 weeks of this period, only animals exposed to chemical treatments were randomly allocated into 2 subgroups and provided orally with either vehicle (water) (n = 7), or Epi at 1 mg/kg/day (n = 7) a dose, based on our previous study. 24 Food intake and body weight were recorded every other day during the exposure and recovery periods. After their final functional assessment, animals were euthanized and hippocampus isolated and collected.
Object recognition task (ORT)
To avoid behavior alterations due to exposure to a new environment, animals were transported to the testing room and habituated individually to the open, empty arena to allow for free exploration for 5 min in 2 consecutive days. As illustrated in Figure 1A on the following day, animals were familiarized during 5 min to two identical objects (A + B) placed in the open field at 10 cm from each wall. Then, rats were sent back to their home cages for 1 h and reintroduced to the arena after one object was exchanged (A + C) and allow them to explore for 5 min (short-term memory). After 24 h, object C was exchanged for object D and animals allowed a 5 min exploratory period (long-term memory). Exploration was considered as pointing the nose toward an object at a distance of less than 1 cm and/or touching it with the nose. Turning around or sitting close to the object was not considered as exploratory behavior. Trials were video recorded using a digital camera. ORT scoring was performed by two different individuals blinded to the experimental conditions. Object recognition was defined as time spent between novel (TN) and the time sum of both objects novel and familiar (TF) [(TN)/(TN+TF)].
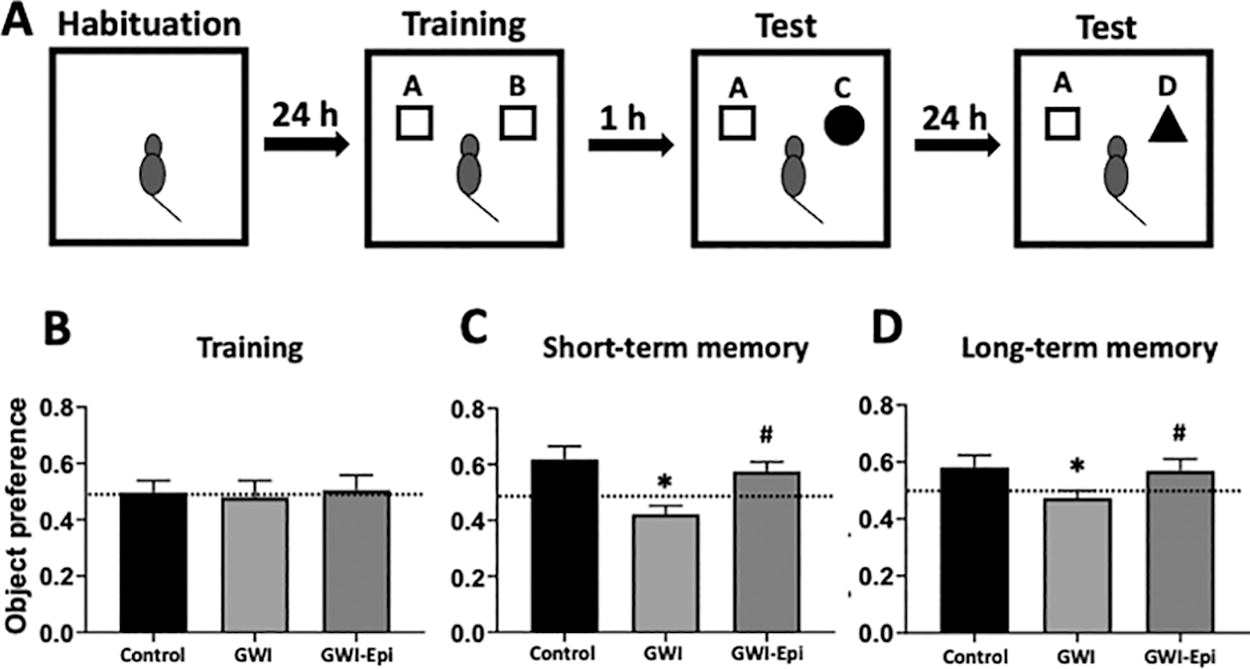
Object recognition test of control and Gulf War Illness (GWI) animals treated with vehicle or (-)-epicatechin (Epi) for 2 weeks. Schematic representation of the object recognition test
Antibodies
Anti-cytochrome c (sc-13156) and glial fibrillary acidic protein (GFAP, a marker of neuroinflammation, sc-9065) antibodies were from Santa Cruz. To measure memory surrogates, anti-phosphatidylinositol-3-kinase (PI3K, 4263), phosphorylated (p)-PI3K (4228), protein kinase-B (AKT, 9272), and p-AKT (directed vs. serine 473, 4058) were purchased from Cell Signaling. Mitofilin (a cristae protein, ab110329), Transcription Factor A Mitochondrial (Tfam; a regulator of biogenesis, ab138351), porin (mitochondrial membrane protein, ab14734), Triggering receptor expressed on myeloid cells 2 (TREM2; a marker of microglial health, ab95470), and ionized calcium-binding adapter molecule 1 (Iba1; a marker of microglial activation, ab178846) were obtained from Abcam. Anti-glyceraldehyde-3-phosphate dehydrogenase (GAPDH, 2118) used for Western blot loading normalization as well as anti-rabbit (7074) and anti-mouse (7076) horseradish peroxidase-conjugated secondary antibodies were from Cell Signaling.
Immunohistochemistry
Hippocampus samples were fixed in a 4% paraformaldehyde-phosphate buffered saline (PBS) pH 7.4 solution and embedded in paraffin, then 5 mm thick sections were obtained and mounted onto glass slides. For immunolabeling, slides were incubated in 0.05% trypsin at pH 7.8 for 12 min at 37°C for antigen retrieval, then incubated with antibodies directed against GFAP or porin overnight at 4°C then rinsed and incubated with a secondary rabbit mouse peroxidase antibody. Color development followed exposure of slides to 0.05% diaminobenzidine hydrochloride and 0.02% H2O2 in buffer. Negative control slides were prepared by substitution of the primary antibody with mouse IgG. Digital images of the sections were obtained at 40× magnification. GFAP and porin positive cells were quantified by blinded operators using 12 randomly chosen digital images of the same size derived from 6 nonconsecutive hippocampus sections.
Mitochondrial protein isolation
Mitochondria were isolated from hippocampus using a Mitochondria Isolation Kit (Thermo Scientific, 89801) as per instructions. Briefly, hippocampi were washed 3× with cold PBS. Then tissue was homogenized in PBS with proteases and phosphatases inhibitor using a polytron. Homogenates were centrifuged at 1000× g for 3 min at 4°C. Pellet was recovered and suspended in BSA/reagent A solution, vortexed for 5 sec, and incubated 2 min. Ten milliliter of mitochondria isolation reagent B were added and incubated in ice for 5 min while vortexing for 5 sec every min. Mitochondria isolation reagent C (800 mL) were added and mixed manually and centrifuged at 700× g for 10 min at 4°C. Pellet was discarded and supernatant centrifugated at 3000× g for 15 min at 4°C. Supernatant (cytosolic fraction) was recovered for further analysis, while mitochondrial pellet was recovered and washed with washing buffer by centrifuge at 12,000× g for 5 min at 4°C. Once isolated, proteins were extracted, quantified by Bradford assay using the Bio-Rad assay dye concentrate (500006), and analyzed by Western blotting.
Western blotting
Tissue samples were homogenized using a polytron in lysis buffer with protease and phosphatase inhibitor cocktails. Homogenates were sonicated and centrifuged. Total protein content was measured in supernatants using the Bradford method. Samples (30 μg) were loaded onto a 4–15% polyacrylamide gel and subsequently electrotransferred to a membrane. Membranes were cut horizontally and were incubated in blocking solution followed by overnight incubation at 4° C with primary antibodies. Membranes were washed and incubated at room temperature with horseradish peroxidase conjugated secondary antibodies. Blots were developed using an enhanced chemiluminescence detection kit (Western blotting detection reagent from Amersham, RPN2236). Band densitometric analyses of digital images was performed using the ImageJ software and signals normalized according to GAPDH bands (used as loading controls). In select cases, membranes were reused and reblotted. After Western blot development, membranes were rehydrated with buffer for 5 min and stripped with Restore-Western blot stripping buffer (Thermo Scientific, 21059) for 5 min, then washed 3 × 5 min with buffer before the blocking procedure and re-exposure to antibodies. All Western blots are provided as Supplementary Data S1.
Pro-inflammatory cytokines
Tissue samples were homogenized with a polytron in 50 mM EDTA pH 8 with protease inhibitor and centrifuged. Supernatant was recovered and assayed for cytokine quantification using a Milliplex magnetic-bead fluorescent immunoassay for tumor necrosis factor (TNF)-α, interferon (IFN)-γ, and interleukin (IL)−1β (Millipore Sigma). The concentration of cytokines was calculated using the standard curve generated and validated using the internal controls of the assay.
Oxidized proteins
As markers of tissue OS, we measured oxidized proteins, protein carbonylation, and lipid peroxidation. Oxidized proteins were measured by Western blot analysis of dinitrophenylhydrazine derived carbonyl groups using an Oxyblot protein oxidation detection kit (Sigma-Aldrich, S7150). Protein samples (20 µg) reacted with the substrate for 15 min, followed by neutralization with a solution containing glycerol and β-mercaptoethanol. Samples were electrophoresed on 4–15% polyacrylamide gel and electrotransferred to a membrane and processed as described above.
Protein carbonylation
Tissue samples were homogenized using a polytron homogenizer in cold buffer and centrifuged. Supernatants were collected and incubated for 15 min with streptomycin sulfate at a concentration of 1% and centrifuged. Total protein carbonylation was measured in supernatants using a colorimetric protein carbonyl assay kit following manufacturer’s instructions (Cayman, 10005020) using a spectrophotometer at 360 nm.
Lipid peroxidation
Lipid peroxidation (marker of OS) was measured by malondialdehyde quantitation. Tissue samples were homogenized using a polytron in lysis solution containing protease inhibitors placed on ice and centrifuged. Supernatants were collected and used to measure malondialdehyde bound to thiobarbituric acid by a colorimetric assay kit according to the manufacturer’s instructions (Cayman, 700870) at 360 nm using a spectrophotometer.
Citrate synthase
As an indicator of mitochondrial function, we evaluated citrate synthase activity in hippocampus using a kit (ScienCell Labs, 8318). Tissue samples were homogenized with a polytron in cold extraction buffer with protease inhibitors. Homogenates were centrifuged, supernatants were recovered and used to measure enzyme activity as the rate of production of the mercaptide ion based on conversion of acetyl-coenzyme A (CoA) to oxaloacetate and CoA-SH. CoA-SH in the presence of 5,50-dithiobis (2-nitrobenzoic acid) produces mercaptide ion. Samples were analyzed in a spectrophotomer at 412 nm. Samples were tested in duplicate and measured at room temperature.
ATP levels
A luciferase-based bioluminescence kit was used to quantify ATP levels (Cayman, 700410). ATP levels were measured in 5 mg of liquid nitrogen frozen hippocampus that was pulverized and then homogenized in kit buffer. Tissue homogenates were diluted in sample buffer and used to measure ATP levels within 10 min of the luciferase reaction as per instructions using a luminometer.
Caspase activity
The activity of caspases as surrogates of apoptosis was measured using caspase-3 (Cat. 39401) and caspase-9 (catalog 65608) kits (Abcam). Hippocampal samples were homogenized and processed as per manufacturer instructions. Caspases 3 and 9 recognize specific sequences and cleave the labeled substrate developing a color reaction which is measured using a microplate reader al 405 nm wavelength.
Data and statistical analysis
Results are expressed as mean ± standard error (SEM). Statistical analysis was pursued using one way analysis of variance followed by a Welch’s post-hoc tests to account for the unequal sample sizes in the control (n = 14) versus the GWI subgroups (n = 7/subgroup) using Graphpad Prism version 8.0. Statistical significance was defined when P < 0.05.
RESULTS
No differences were noted in food intake or body weight among all groups (data not shown). ORT (Fig. 1A) was used to evaluate short- and long-term memory in rats. Training of the animals (Fig. 1B) did not show object preference score differences between the groups. GWI animals demonstrated an impairment in object preference score both in short (Fig. 1C) or long-term memory (Fig. 1D). Epi treatment yielded short and long-term memory scores comparable to those of controls which, were significantly improved versus GWI. Pro-inflammatory cytokine levels were assessed in samples of rat hippocampus and results are shown in Figure 2. In GWI animals, pro-inflammatory cytokines levels increased significantly by ∼80% for IL-1β (Fig. 2A), 240% for TNF-α (Fig. 2B) and 65% for IFN-γ (Fig. 2C) versus controls. Epi treatment suppressed IL-1β, TNF-α, and IFN-γ levels to values comparable to those of controls and were different versus GWI. Results derived from OS level assessments are shown in Figure 3. GWI animals significantly increased lipid malondialdehyde content versus controls by ∼270% (Fig. 3A, from 1.4 ± 0.6 to 3.8 ± 0.7 nmol/mg). Epi treatment significantly suppressed malondialdehyde levels to 2.2 ± 0.9 nmol/mg versus GWI animals. Total protein carbonylation (Fig. 3B) was significantly increased by ∼270% in GWI (48 ± 10 nmol/mg) versus controls (17.6 ± 7 nmol/mg). Epi treatment demonstrated a trend (P = 0.06) toward reduced protein carbonylation levels to 31 ± 0.9 nmol/mg of protein versus GWI animals. Changes in protein oxidation levels as detected by Westerns (Oxyblots, Fig. 3C) paralleled those noted for protein carbonylation. As indicators of neuroinflammation, the relative protein levels of the glial associated activators Iba1 and GFAP were evaluated by Westerns (Fig. 4). GWI animals showed a ∼70% and ∼110% increase in the markers of neuroinflammation Iba1 and GFAP levels, respectively, versus controls. Epi treatment significantly reversed Iba1 levels to values similar to controls, while GFAP protein levels demonstrated a trend (P = 0.06) to be decreased by ∼80% (Fig. 4A). These results were consistent with values derived from GFAP immunohistochemistry and related quantification (Fig. 4B). Caspase 3 and 9 activity were used as surrogates of apoptosis and results are shown in Figure 5. In GWI animals, caspase activity was increased (∼170% caspase 3 and ∼200% for 9) versus controls. Epi treatment significantly decreased activity for both caspases versus GWI animals. As shown in Figure 5B, cytochrome c levels increased significantly in the cytoplasm of GWI animals versus controls suggesting increased levels of apoptosis. Epi treatment significantly restored cytochrome c levels versus GWI rats. As a measure of cell survival pathway activity, TREM2, PI3K and AKT levels were evaluated. Figure 6 reports on decreases in TREM2 content by ∼70% in GWI animals versus controls which was partly restored with Epi. The active (phosphorylated) forms of PI3K and AKT were decreased in GWI by ∼40% versus controls and were completely restored with Epi. To determine the impact on mitochondrial endpoints, samples of hippocampus were analyzed by Westerns for changes in relative protein levels for mitofilin and the regulator of mitochondrial biogenesis, Tfam (Fig. 7A). In GWI animals, significant decreases ∼60% versus controls occurred for both proteins. By immunohistochemistry, the content of the mitochondrial outer membrane protein porin also demonstrated decreased levels in GWI animals (∼45%) versus controls (Fig. 7B). Epi treatment partially restored the levels of mitofilin, Tfam, and porin. To document changes in mitochondrial function, we used as surrogates citrate synthase activity and ATP levels (Fig. 7C and D). Citrate synthase activity significantly decreased by ∼ 50%, while ATP levels had a reduction of ∼ 45% in GWI versus controls. These changes were fully restored with Epi treatment.

Pro-inflammatory cytokine levels in the hippocampus of control, Gulf War Illness (GWI) and (-)-epicatechin (Epi) treated GWI rats. Protein levels are reported for interleukin (IL)−1β
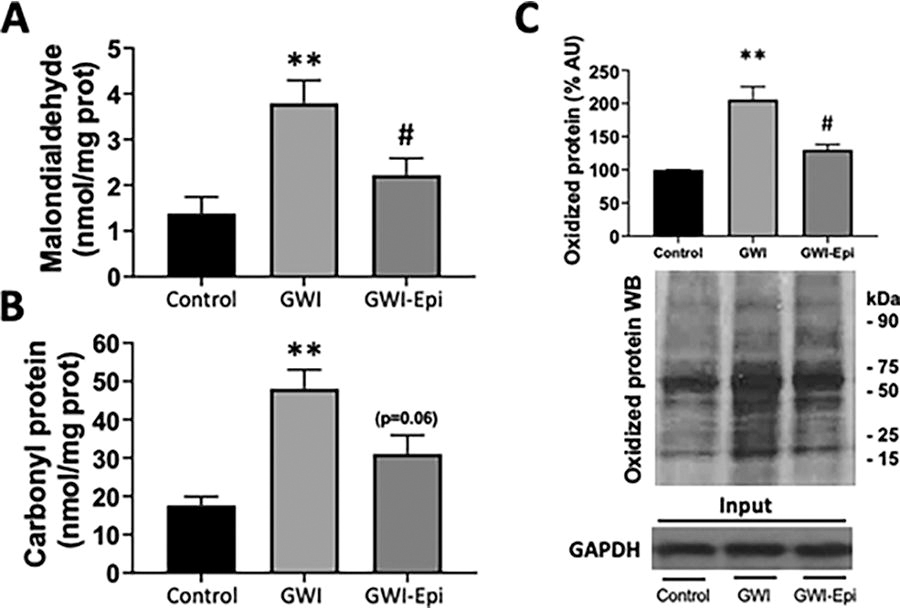
Oxidative stress levels in the hippocampus of control, Gulf War Illness (GWI) and (-)-epicatechin (Epi) treated animals.
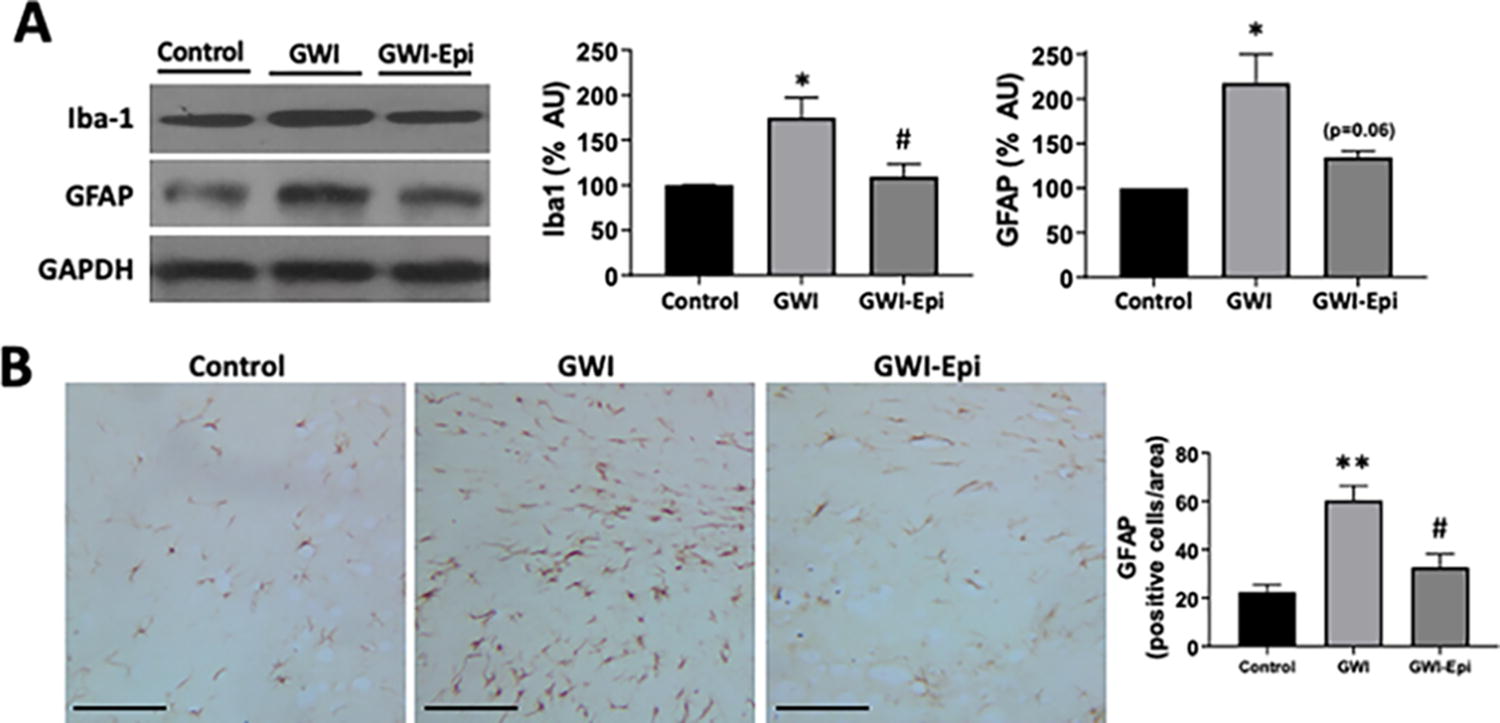
Protein levels of Iba1 and glial fibrillary acidic protein (GFAP) in the hippocampus of control, Gulf War Illness (GWI), and (-)-epicatechin (Epi) treated rats. Representative Western blot images observed for Iba1 and GFAP and their loading control glyceraldehyde-3-phosphate dehydrogenase (GAPDH) are shown in panel A. Control values were set as 100%. Representative GFAP immunostaining and the quantification obtained
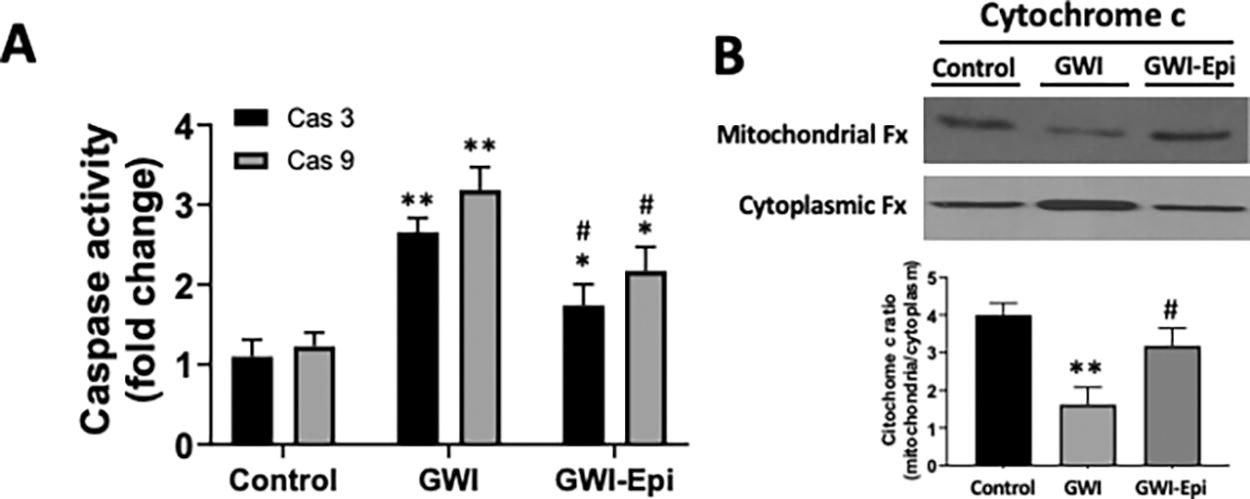
Caspase 3 and 9 activity and protein levels of cytoplasmic and mitochondrial cytochrome c in the hippocampus of control, Gulf War Illness (GWI), and (-)-epicatechin (Epi) treated rats. Relative changes noted in Caspase (Cas) 3 and 9 activity
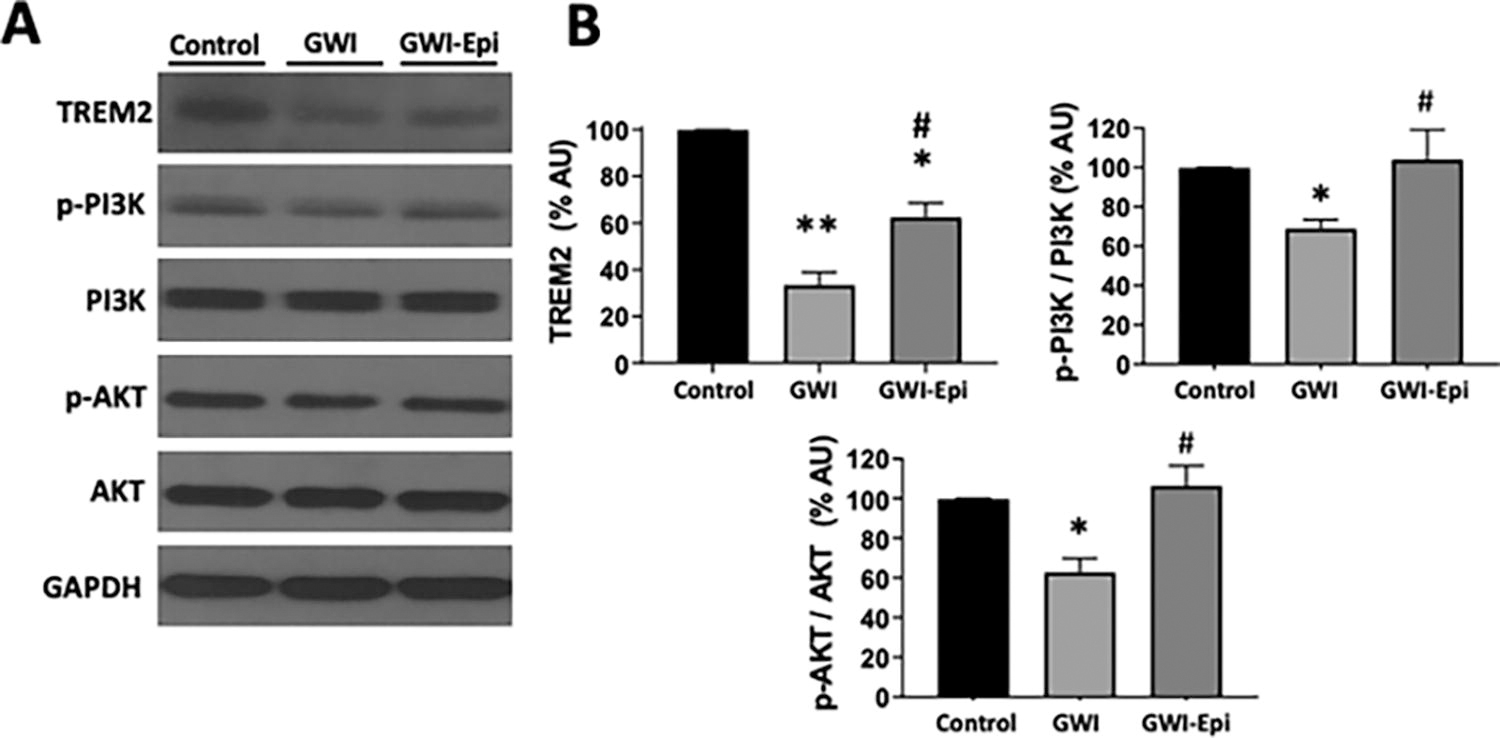
Protein levels of neuronal survival pathways of control, Gulf War Illness (GWI), and (-)-epicatechin (Epi) treated rats. Representative Western blot images observed for triggering receptor expressed on myeloid cells 2 (TREM2), active (phosphorylated) forms of PI3K and AKT
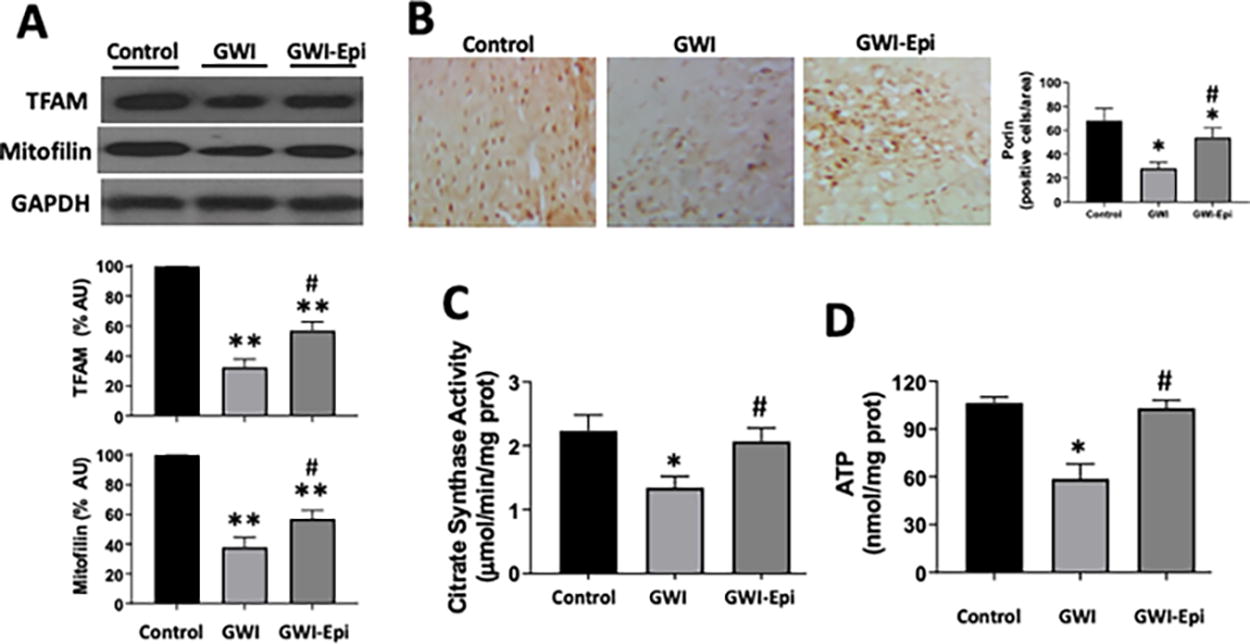
Mitochondrial protein levels in hippocampus of control, Gulf War Illness (GWI), and (-)-epicatechin (Epi) treated rats. Representative Western blot images observed for TFAM and mitofilin
DISCUSSION
Results from this study demonstrate that in an animal model of GWI induced cognitive dysfunction and neurodegeneration, Epi treatment yields improvements in short- and long-term memory, hippocampus OS and pro-inflammatory cytokine levels, markers of neuroinflammation, apoptosis, modulators of cell survival, and mitochondrial structure and function.
Cognitive dysfunction in Veterans is reported as one of the most debilitating hallmark symptoms of GWI. A recent meta-analysis was performed by Janulewicz et al., compiling a review of 238 studies including American, British, and Danish Veteran cohorts. 25 Across all neuropsychological tests, results showed that Gulf-deployed Veterans performed worse with reductions in visuospatial performance, learning and memory tests and in attention and executive function outcomes. As reported by structural and functional neuroimaging and biomarker studies, alterations in specific brain regions may account for impaired cognitive functions in GW Veterans, such as reduced volume and increased mean diffusivity in white brain matter, 26,27 decreases in metabolite levels 5 and in cerebral blood flow, 28 reduced gray matter volume, 29 and altered gray matter activity in response to stimuli. 28,30
The hippocampus is a region of the brain recognized to play key roles in learning and memory, and this structure appears to be highly sensitive to inflammatory processes. 31 Inflammation of the hippocampus is known to block neurogenesis 32 which is critical for learning and memory. 33 Furthermore, evidence indicates that high levels of inflammation can trigger hippocampus atrophy by promoting neuronal death. 34 In studies of GW Veterans, associations have been reported between neurocognitive symptoms, high levels of inflammatory cytokines, and reduced hippocampal volume. 35 –37 In a study of 246 GW Veterans, O’Donovan reported reduced levels of hippocampal volume as per magnetic resonance imaging results which, associated with higher levels of soluble receptor II for TNF-α. 36 A study performed by Alshelh et al., reported on in vivo imaging of a neuroinflammation marker using a radiolabeled marker which binds to a translocator protein that is known to be upregulated in activated microglia/macrophages and astrocytes. 12 Veterans with GWI and healthy controls were examined. GW Veterans demonstrated widespread cortical elevations in signals in areas including precuneus, prefrontal, primary motor, and somatosensory cortices, thus suggesting the presence of an active inflammatory process even at 30+ years after war exposure. Evidence for generalized inflammation has also been reported even in the very chronic stage of the disease. In a 2016 study published by Johnson et al., plasma C-reactive protein, leptin, brain-derived neurotrophic factor, and matrix metalloproteinase-9 were significantly higher in the GW Veteran group versus controls. 38 Parkitny et al., also reported that for GW Veterans, higher fatigue severity days were associated with significantly greater IL-1β and IL-15 plasma levels. 39
Underlying the inflammatory process can be the development of MD and OS. An analysis of indirect clinical evidence was presented in a 2012 report where it was stated that GWI is compatible with a paradigm by which uncompensated exposure to oxidative/nitrative stressors accompanies and triggers MD, yielding altered bioenergetics and multiple adverse downstream effects. 40 In further support of this hypothesis a study by Koslik et al., evaluated the presence of MD in skeletal muscle of GWI patients by examining post-exercise phosphocreatine-recovery time constant using 31Phosphorus Magnetic Resonance Spectroscopy which is recognized as a “robust and practical” index of in vivo mitochondrial function. 8 Recovery times were significantly prolonged in GW Veterans vs. controls. In support of this mechanism, there are the results from a pilot trial where in 85% of male GF Veterans coenzyme Q10 supplementation increased a quality of life of measure and physical function versus placebo. These results led to the recent implementation of a phase III trial in GWI patients. Further support comes from a study by Chen et al., where blood samples were used to quantify mitochondrial and nuclear DNA lesion frequency and mitochondrial DNA copy number from blood mononuclear cells. 41 Samples were also used to analyze nuclear DNA lesion frequency and mitochondrial complexes I and IV activity. Both mitochondrial DNA lesion frequency and copy number were elevated in GW Veterans versus controls. While complex I and IV activity were similar between groups, greater lesion frequency was associated with reduced complex I and IV activity. Thus, clinical evidence supports the hypothesis of the involvement of OS, MD, and neuroinflammation in triggering cognitive dysfunction.
Studies in animal models of GWI demonstrate that exposure to suspect GW agents and stress, leads to persistent cognitive and mood dysfunction. 42 In animal models of GWI, cognitive dysfunction is associated with brain OS, MD, astrocyte hypertrophy, microglial activation, persistent neuroinflammation, leaky blood-brain barrier, and diminished hippocampal neurogenesis. 7,10,11,14,15,43 Cognitive dysfunction was typified by the inability of animals to discern changes in an object location test 44 and by impaired recognition memory, tasks that largely depends on the hippocampus. 45 In several of these models, OS and increased proinflammatory cytokines in blood parallel changes in brain thus, indicating the presence of systemic inflammation. 15 In rats, exposure to GWI chemical with or without stress also leads to reduced hippocampal volume by ∼10% and neuron loss. 11 Furthermore, reductions in hippocampus neurogenesis and in the integration of newly generated neurons are reported.
Investigations have also evaluated chronic changes in the expression of genes related to increased OS, MD, and inflammation in the hippocampus of GWI rats. 46 OS and MD were evident 6 months after exposure from changes in the expression of genes involved in OS-response, reactive oxygen species metabolism, oxygen transport and antioxidant activity. Elevated OS were confirmed as per increases in malondialdehyde, 4-hydroxynonenal and protein carbonyl levels. 15 A study revealed that cognitive dysfunction and neuroinflammation as typified by activated microglia, reactive astrocytes and increased TNF-α and IL-1β levels persist even 10 months after exposure to chemicals and stress. 47 Thus, long term changes in animal models parallel the persistence of symptoms in GW Veterans.
With the aim of identifying safe and effective agents aimed at treating GWI, natural compounds have been tested in pre-clinical models of GWI. Curcumin has potent anti-inflammatory and anti-oxidant effects via its pleiotropic actions including the NF (nuclear fator)-κB and signal transducer and activator transcription (STAT)3 signaling pathways. 48 In a study by Kodali et al., male rats exposed for 28 days to suspect chemicals and stress were subsequently administered curcumin as a daily injection and compared to vehicle. Animals treated with curcumin exhibited improved cognitive and mood function, hippocampal neurogenesis and reduced inflammation. 14 Shetty et al., evaluated the efficacy of luminol a compound that apparently promotes redox homeostasis for improving cognitive and mood function in GWI rats. 15 Four months later after chemical and stress exposure, GWI rats received different doses of luminol or vehicle for 8 weeks. GWI rats receiving higher doses of luminol displayed better cognitive and mood function in association with reinstatement of redox homeostasis as per the return of several OS markers including malondialdehyde and protein carbonyls to control levels in brain and blood. Luminol treatment also resulted in anti-inflammatory and pro-neurogenic effects, as per the apparent reduced density of astrocyte hypertrophy and activated microglia, and increased neurogenesis. Luminol also normalized the concentration of multiple pro-inflammatory cytokines in blood including IL-1β and TNF-α. Altogether, the effects of Epi parallel those reported for these compounds.
We previously reported on the beneficial effects of Epi on multiple markers of nervous system health and function in normal, young 3 month old 49 and in 26 month old male mice. 50 Using healthy 3 month animals, we reported on the beneficial effects of 15 days of treatment with Epi on multiple markers of frontal cortex neurogenesis, capillary density and short term memory. In aged mice, we examined the capacity of Epi to mitigate aging associated hippocampus OS, inflammation, hyperphosphorylation of tau protein, soluble β-amyloid protein levels, cell survival, memory, anxiety-like behavior levels and systemic inflammation. Animals were subjected to 4 weeks of Epi treatment (1 mg/kg/day by gavage) and samples of hippocampus processed. Assessments of protein carbonyls and malondialdehyde levels demonstrated significant increases (∼3-fold) with aging that were partially suppressed by Epi. The protein levels of the GFAP, Iba1, pro-inflammatory cytokines IL-1β, IL-3, -5, -6, and -15, cyclooxygenase 2, TNF-α, and IFN-γ increased with aging and were significantly decreased with Epi. The anti-inflammatory cytokines IL-10 and −11 decreased with aging and were restored with Epi. Epi also reversed the aging effects on the hyperphosphorylation of tau, increased soluble β-amyloid levels (∼2-fold), cellular death (as per caspase 3 and 9 activity) and reduced nerve growth factor and TREM2 levels. Measures of anxiety like-behavior and memory demonstrated improvements with Epi treatment.
We have reported on the beneficial effects of Epi on skeletal muscle structure and function including in pilot clinical studies in heart failure patients and Becker muscular dystrophy. 51 –53 Given this precedent, we used our rat model of GWI to examine the potential of Epi to reverse skeletal muscle atrophy and dysfunction and decrease mediators of inflammation. 24 GWI rats were provided Epi for 2 weeks (1 mg/kg/day by gavage) or water for controls. GWI animals lost muscle mass and function accompanied by increases in modulators of atrophy, degradation markers and plasma pro-inflammatory cytokines TNF-α, IL-1β, and IFN-γ levels. Treatment with Epi yielded either a significant partial or full normalization of the above stated indicators relative to healthy, normal controls but more importantly muscle (grip) strength and treadmill performance was restored. Thus, it is plausible that Epi may exert actions in GF Veterans that may specifically target fatigue.
LIMITATIONS
Gulf War Veterans also included female troops. This study was conducted only using male animals. It is possible that the phenotype observed and effects of treatment may differ in female animals and have implications for their possible extrapolation to female Veterans. This, study was also implemented in young animals whereby the effects of suspect agents and treatment were examined. As the Gulf War occurred more than 30 years ago, the current presentation of GWI in Veterans includes an aging component that is not paralleled in the current study design.
SUMMARY AND CONCLUSIONS
In this study, which focuses on cognitive impairment, we demonstrate a capacity of Epi to trigger improvements in short and long-term memory in GWI rats yielding values similar to healthy controls. The beneficial effects noted on cognitive function can be accounted by the pleiotropic nature of Epi induced actions. Increases in hippocampus OS and pro-inflammatory cytokine levels were fully normalized with Epi thus, becoming comparable to controls. GWI induced increases in markers of hippocampus neuroinflammation and cell death were significantly reduced with Epi furthermore, the neuronal survival signaling pathways were partially or fully restored by Epi. Recognized markers of mitochondrial function were also fully restored by Epi. Given the extensive body of clinical evidence supporting the beneficial actions of cocoa flavanols on skeletal muscle and nervous system endpoints as well as their tolerance and safety profile, the design of clinical trials using Epi in GW Veterans warrant consideration.
Footnotes
AUTHORS’ CONTRIBUTIONS
I.R.-S.: Investigation, Data curation, Writing—Original draft preparation, Reviewing, and Editing. V.N.Y.: Investigation, Data Curation, and Writing—Reviewing and Editing. J.E.-R.: Investigation, Data curation, Writing, Reviewing, and Editing. I.R-G.: Investigation and Data curation. C.P.-F.: Investigation and Data curation. P.M.-L.: Investigation and Data curation. R.O.-R.: Investigation and Data curation. J.E.-M.: Investigation and Data curation. G.C.: Supervision, Data Curation, Formal Analysis, and Writing—Reviewing and Editing. F.V.: Conceptualization, Funding Acquisition, Project Administration, Supervision, Data Curation, and Writing—Reviewing and Editing.
AUTHOR DISCLOSURE STATEMENT
F.V. (co-founder) and G.C. are stockholders of Epirium Bio Inc.
FUNDING INFORMATION
This study was partly supported by DoD PR150090, GW180044, NIH DK98717, AG47326, VA I01BX3230, I01BX5894 to F.V. and IPN 20195988 and CONACYT 283938 to I.R.-S.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Data S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.