
Abstract
The sialic acid N-acetylneuraminic acid (NANA), an essential factor in bioregulation, is a functional food component that is known to have beneficial health effects, but its antiobesity effect has not been clearly understood. Adipocyte dysfunction in obesity involves a decrease in the level of NANA sialylation. In this study, we investigated the antiobesity effect of NANA in mice fed a high-fat diet (HFD) and in 3T3-L1 adipocytes. Male C57BL/6J mice were randomly divided into three groups and administered the following diets: a normal diet, an HFD, and an HFD with 1% NANA supplementation for 12 weeks. NANA supplementation significantly reduced body weight gain; epididymal adipose tissue hypertrophy; and serum lipid, fasting glucose, and aspartate transaminase levels compared with those in HFD mice. The percentage of lipid droplets in hepatic tissue was also decreased by NANA supplementation in HFD mice. The downregulation of Adipoq expression and upregulation of Fabp4 expression induced by HFD in epididymal adipocytes were improved by NANA supplementation. The downregulation of Sod1 expression and increase in malondialdehyde level were induced by HFD, and they were significantly improved in the liver by NANA supplementation, but not in epididymal adipocytes. However, NANA supplementation had no effect on sialylation and antioxidant enzyme levels in mouse epididymal adipocytes and 3T3-L1 adipocytes. Overall, NANA exerts antiobesity and antihypolipidemic effects and may be beneficial in suppressing obesity-related diseases.
INTRODUCTION
Obesity, a global health problem, can trigger various diseases including cardiovascular disease and liver steatosis and conditions such as diabetes and hypertension. The visceral adipose tissue (AT) is an endocrine organ that controls the development of metabolism-related diseases. Obesity causes an increase of visceral ATs, can lead to dysfunctional adipocytes, and is associated with various obesity-related complications/diseases. 1 –5 In our previous study, we found that a decrease in the level of sialylated protein with N-acetylneuraminic acid (NANA) in ATs is related to adipocyte dysfunction caused by obesity. 6
These changes may lead to the development of a novel glycan-targeting strategy for the treatment of obesity-related diseases. Therefore, we speculated that the oral supplementation of NANA might increase NANA sialylated protein levels in ATs and that NANA might have an antiobesity effect.
NANA is distributed throughout the human body and is present at the end of glycan chains on the cell membrane and is an essential factor in bioregulation. It is also a component of some foods and is abundant in mammalian breast milk, edible bird's nest, and red meat. Studies have demonstrated the physiological health benefits of NANA, such as antioxidant activity; NANA has been confirmed to be safe as a supplement. 7 –11 NANA supplementation increases the levels of sialylated glycoproteins and glycolipids in the brain, intestine, and muscles, 12 and improves memory, learning, 13 immune function, 14 and distal myopathy. 15 However, the effect of oral supplementation of NANA on adipocytes has not yet been investigated.
In this study, we analyzed the effects of NANA supplementation on obesity, sialylated protein levels, adipocyte function regulation, and antioxidant capacity using a high-fat diet (HFD)-induced obesity model mice.
MATERIALS AND METHODS
Animals and diet
Fifteen male C57BL/6J mice (age, 6 weeks) were obtained from Charles River Laboratory (Kanagawa, Japan) and randomly housed (five animals per cage) in a temperature-controlled room at 24°C with a 12-/12-h light–dark cycle (lights off at 21:00 h). After 1 week of acclimatization, the mice were assigned to three groups, with five mice each, and administered the following diets: high-fat diet (HFD group), HFD supplemented with 1% NANA (HFD-NANA group), and control diet (CON group). The composition of the experimental diet is given in Table 1. The diet was fed for 12 weeks. AIN-93G (Research Diets, New Brunswick, NJ, USA) was used as the CON diet; HFD comprised a 55% fat energy ratio and was supplemented with lard based on the AIN-93G (Research Diets).
Composition of Experimental Diets
CON diet was prepared based on AIN-93G (American Institute of Nutrition). CON, control diet; HFD, high-fat diet; HFD-NA, HFD mixed with 1% NANA; NANA, N-acetylneuraminic acid; PFC, protein/fat/carbohydrate.
NANA (Nagara Science, Nagoya, Japan) was administered by supplementing 1% NANA in HFD. The mice were allowed free access to food and drinking water throughout the acclimation and experimental periods. Individual mouse weights and food consumption rates were recorded twice weekly. At the end of the 12-week feeding period, the mice were starved for 6 h and then anesthetized through isoflurane inhalation. Blood was sampled through the orbital cavity and centrifuged at 5000 g for 15 min to obtain serum. Biochemical tests were performed by Oriental Yeast (Shiga, Japan). The mice were euthanized through cervical dislocation under anesthesia, and the livers, kidneys, epididymal ATs, renal ATs, and mesenteric ATs were immediately removed and weighed.
This study was approved by the Daito Bunka University Animal Experiment Committee at Daito Bunka University (ASH19-001) in Japan; the experimental protocols were performed according to the guidelines of the Japanese Council on Animal Research.
RNA extraction, reverse transcription, and real-time polymerase chain reaction
The total RNA was extracted from tissues and cells using the RNeasy Plus Universal Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer's instructions. The total RNA (1 μg) was reverse transcribed using Superscript III (Life Technologies, Carlsbad, CA, USA). For real-time polymerase chain reaction (PCR), cDNA was mixed with the TaqMan Universal PCR Master Mix (Life Technologies) and amplified. mRNA levels were normalized to the corresponding ribosomal RNA levels. The protocol was as follows: using the two-step PCR method, initial denaturation was performed at 95°C for 30 s; amplification was performed at 95°C for 5 s, 60°C for 34 s for 40 cycles; and annealing was performed at 60°C for 30 min.
TaqMan primer probe sets for mRNA quantification were purchased from Applied Biosystems (Waltham, MA, USA): 18s RNA, 4308329; adiponectin (Adipoq), Mm00456425_m1; peroxisome proliferator-activated receptor gamma (Pparg), Mm00440940_m1; fatty acid-binding protein 4 (Fabp4), Mm00445878_m1; CCAAT enhancer-binding protein β (Cebpb), Mm00843434_s1; St6gal1, Mm00486119_m1; superoxide dismutase (Sod1), Mm01344233_g1; Sod2, Mm01313000_m1; and glutathione peroxidase 1 (Gpx1), Mm00515041_m1.
Antioxidant measurement
Lipid peroxidation was analyzed by measuring the production rate of thiobarbituric acid-reactive substances (TBARSs) and expressed as malondialdehyde (MDA) equivalents, using a TBARS assay kit (Cayman Chemical, MI, USA). The amount of MDA was assessed by measuring the absorbance of samples at 535 nm, and the results are expressed as micromolar per milligram. The antioxidant enzymes Sod1, Sod2, and Gpx1 were analyzed using real-time PCR, as described in the previous section.
Morphological analysis of epididymal ATs and liver tissues
The epididymal ATs and liver tissues were fixed with 10% paraformaldehyde, embedded in paraffin, sectioned to a thickness of 3 μm, and stained with hematoxylin–eosin (HE). For morphological analysis, 50 adipocyte areas per mouse were microscopically examined in a blinded manner using ImageJ version 1.52 (National Institutes of Health, Bethesda, MD, USA). Lipid droplet area in the liver was measured in five random fields per mouse; ≥20% lipid content index was used to diagnose diseased fatty liver. The average value of each group was statistically compared.
Blot analysis of epididymal ATs and 3T3-L1 adipocytes
We analyzed NANA sialylated protein expression using blot analysis with Sambucus sieboldiana agglutinin (SSA) lectin. Experimental details have been reported previously. 6 In brief, epididymal ATs and 3T3 L1 adipocytes were homogenized; then, insoluble debris and thick lipid layer were removed after centrifugation at 2500 g for 15 min. The supernatants were collected, and proteins were separated using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and then transferred on to nitrocellulose membranes (Bio-Rad, Hercules, CA, USA). After blocking, the membranes were incubated with primary antibodies or biotinylated SSA lectin, followed by incubation with secondary antibodies or horseradish peroxidase-conjugated streptavidin (Vectastain ABC Standard Kit; Vector Laboratories, Burlingame, CA, USA).
Proteins were detected using the Western Lightning ECL Pro kit (PerkinElmer Life Sciences, Waltham, MA, USA) and the ImageQuant LAS-4000mini system (GE Healthcare, Chicago, IL, USA). Total protein levels were detected through Coomassie Brilliant Blue staining using an adjustment value.
SSA lectin precipitation
Epididymal AT lysates (300 μg of protein in 500 μL of tris buffered saline containing 1% Triton X-100, a protease inhibitor mixture, and phosphatase inhibitors [Roche Applied Science]) were incubated with 30 μL of SSA-agarose (Honen Co.) for 16 h with gentle shaking at 4°C. The precipitates were washed four times with phosphate-buffered saline and then eluted by boiling with SDS sample buffer. The precipitated proteins were separated using SDS-PAGE, transferred onto nitrocellulose membranes, and then probed with anti-integrin-β1.
Lipid accumulation in and viability of 3T3-L1 cells
3T3-L1 cells were cultured as previously reported. 6 In brief, 3T3-L1 preadipocytes were cultured at 2 days postconfluence (day 0) and then induced to differentiate in Dulbecco's modified Eagle medium (DMEM) containing 10% fetal bovine serum (FBS), 1 μg/mL insulin, 1 μM dexamethasone, and 0.5 mM 3-isobutyl-1-methylxanthine. At 2 days postdifferentiation (day 2), the medium was replaced with DMEM containing 1 μg/mL insulin and 10% FBS. During the next 12 days, the medium was replaced every alternate day with fresh medium. Lipid accumulation was assessed through staining with Oil Red O (Sigma-Aldrich, St. Louis, MO, USA). The effect of NANA on cell viability was assessed using the WST-8 assay (Dojindo, Japan).
Statistical analysis
The results are presented as means ± standard errors. Statistical evaluations were performed using a one-way analysis of variance, followed by Scheffe's multiple comparison post hoc test, with SPSS version 25.0 (IBM, Armonk, NY, USA). Results with P < .05 were considered significant.
RESULTS
Bodyweight gain, energy intake, and organ weights
During the 12-week experimental diet-administration period, the body weight of HFD-fed mice significantly increased compared with that of CON-fed mice, but body weight gain was significantly suppressed upon NANA supplementation (Table 2). Energy intake and the absolute weight of the organs did not differ among the groups.
Body Weights, Food Intakes, and Organ Weights
The values represent the means ± standard errors (n = 5).
P < .05 versus CON.
P < .05 versus. HFD by Scheffe's multiple comparison test.
Serum biochemical marker
The HFD-induced increases in serum lipid, total cholesterol (TC), triglyceride (TG), and low-density lipoprotein (LDL) levels were significantly suppressed by NANA supplementation (Table 3). Notably, the low-density lipoprotein levels were reduced by NANA supplementation to a level similar to that of the CON group. The fasting glucose and aspartate transaminase (AST) levels increased in the HFD group, and they were suppressed by NANA supplementation.
Serum Biochemical Parameters
The values represent the means ± standard errors (n = 5).
P < .05 versus CON.
P < .05 versus HFD by Scheffe's multiple comparison test.
ALT, alanine transaminase; AST, aspartate transaminase; FG, fasting glucose; HDL, high-density lipoprotein; LDL, low-density lipoprotein; NEFA, nonesterified fatty acids; T-Cho, total cholesterol; Trig, triglyceride.
Visceral AT weight and adipocyte area
The weights of epididymal, perirenal, and mesenteric ATs (per gram of body weight) were measured as an index of visceral adiposity. The epididymal and perirenal AT weights were markedly higher in the HFD group than in the CON group (61.7 mg/g vs. 36.2 mg/g and 17.7 mg/g vs. 11.0 mg/g body weight, respectively; P < .05, Fig. 1A); whereas there was no difference in mesenteric AT weight between the groups. This increase in epididymal AT weight in the HFD group was suppressed upon NANA supplementation.
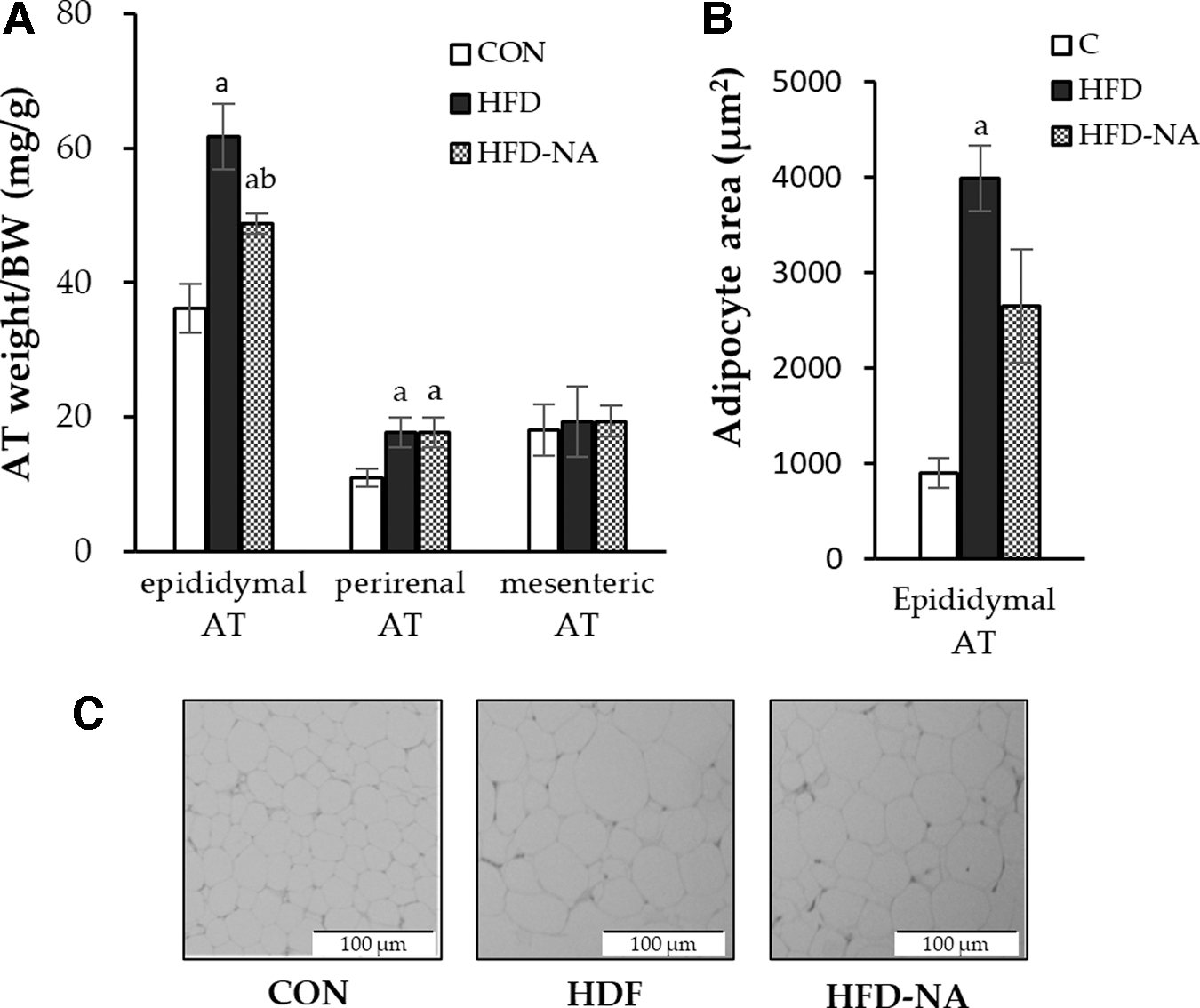
Weight and histological observation of visceral ATs.
Therefore, we focused on epididymal AT and assessed the effect of NANA supplementation morphologically using HE staining. Epididymal ATs showed significant adipocyte hypertrophy, which was evaluated using the adipocyte area in the HFD group; it was mitigated by supplementation with NANA (Fig. 1B, C).
mRNA levels of lipogenesis-related and antioxidant enzymes in epididymal ATs
The effects of NANA supplementation on mRNA expression in epididymal ATs were analyzed (Fig. 2). NANA supplementation improved the function of adipokines in epididymal ATs. The level of Fabp4, which encodes a fatty acid-binding protein specifically in adipocytes, promoting lipid synthesis and inducing insulin resistance, 16,17 was increased by HFD intake but downregulated by NANA supplementation. The expression of Adipoq, which suppresses obesity-related diseases, 2,4,5,18,19 was decreased by HFD intake, but significantly upregulated by NANA supplementation (Fig. 2A). Furthermore, the expression levels of Pparg and Cebpb, the genes involved in adipocyte differentiation, were upregulated upon HFD intake; changes induced by NANA supplementation were not significant, and NANA did not directly affect adipocyte differentiation.
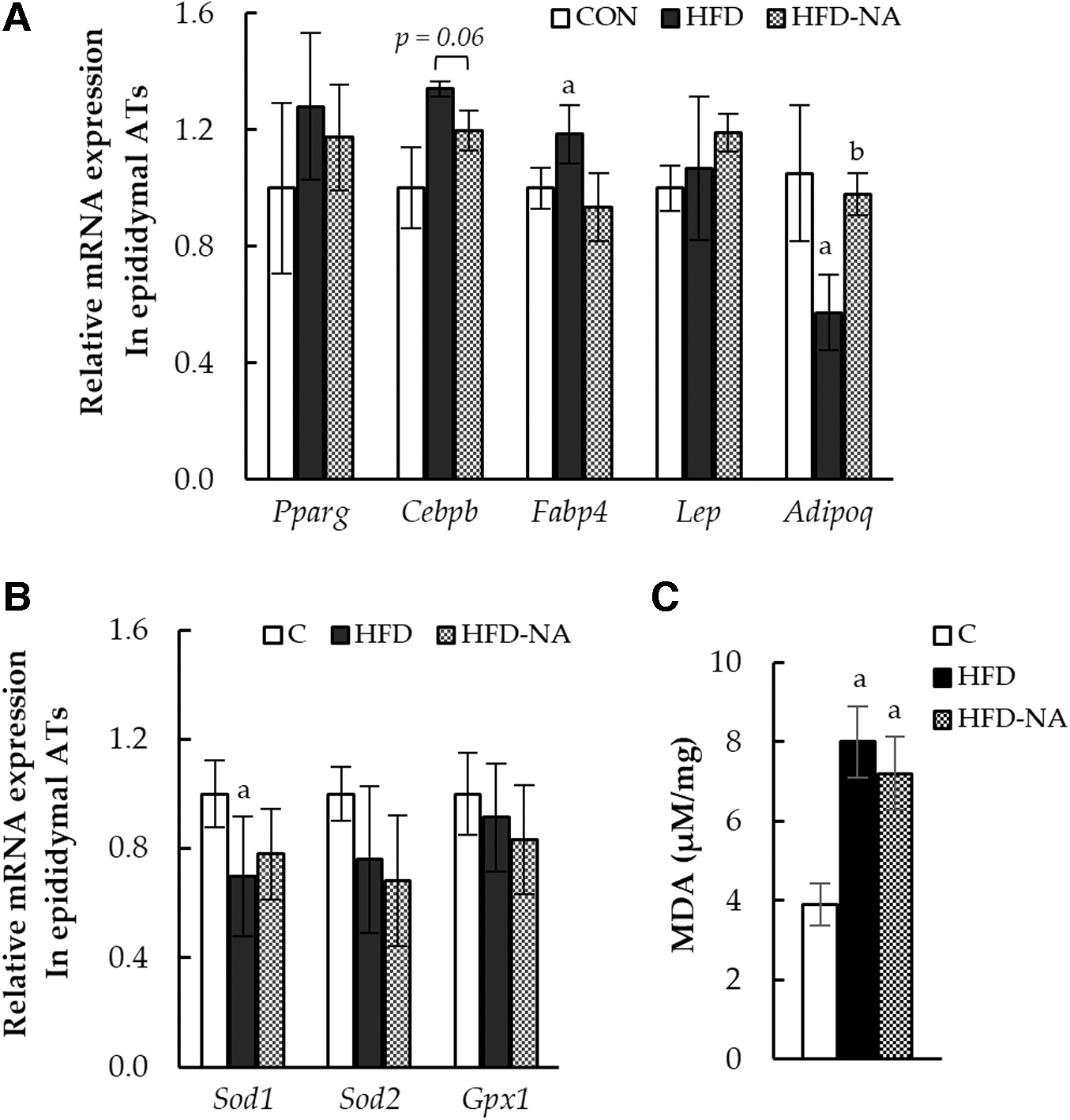
mRNA levels of lipogenesis-related and antioxidant enzymes in epididymal ATs.
Regarding antioxidant enzyme-encoding genes, the HFD group showed a significant decrease in Sod1 expression, which was not observed upon NANA supplementation (Fig. 2B). In addition, the MDA level, which was measured to evaluate the production of lipid peroxides, markedly increased with HFD intake, and NANA supplementation did not have a significant effect on the MDA levels (Fig. 2C).
St6gal1 expression and α2,6-sialylated protein levels in epididymal ATs
Glycosyltransferase st6gal1, an enzyme that sialylates NANA, and the role of sialylated proteins were analyzed. The mRNA expression of St6gal1 in epididymal ATs decreased upon HFD intake, and it was not affected by NANA supplementation (Fig. 3A). To detect the α2,6 sialylated protein (st6gal1 product) level, as a NANA sialylation indicator, lectin blots were analyzed using SSA. The level of α2,6-sialylated protein in epididymal ATs was decreased by HFD intake, and no change was observed upon NANA supplementation (Fig. 3B, C).

Expression of St6gal1 and sialylated protein in epididymal ATs.
Integrin-β1 is known as an α2,6-sialylated protein in mouse epididymal ATs, and its activity is impaired by the loss of α2,6-sialylation; these phenomena cause adipocyte hypertrophy and dysfunction. 6 We found that the level of α2,6-sialylated integrin-β1 in epididymal ATs was significantly reduced by HFD intake, and no change was observed upon NANA supplementation (Fig. 3D, E).
Histological lipid droplet percentage and antioxidant capacity in the liver
To observe the effects of NANA supplementation on the liver, the hepatic lipid index was calculated as the percentage of lipid droplets per area using HE-stained hepatic tissues. The lipid droplet index significantly increased with HFD intake, but it was decreased upon NANA supplementation (Fig. 4A, B). Regarding antioxidant enzyme-coding genes, Sod1 expression was markedly decreased by HFD intake, which was significantly inhibited by NANA supplementation (Fig. 4C). The MDA level, measured as a product of hepatic lipid oxidation, was increased by HFD intake, but was inhibited by NANA supplementation (Fig. 4D).
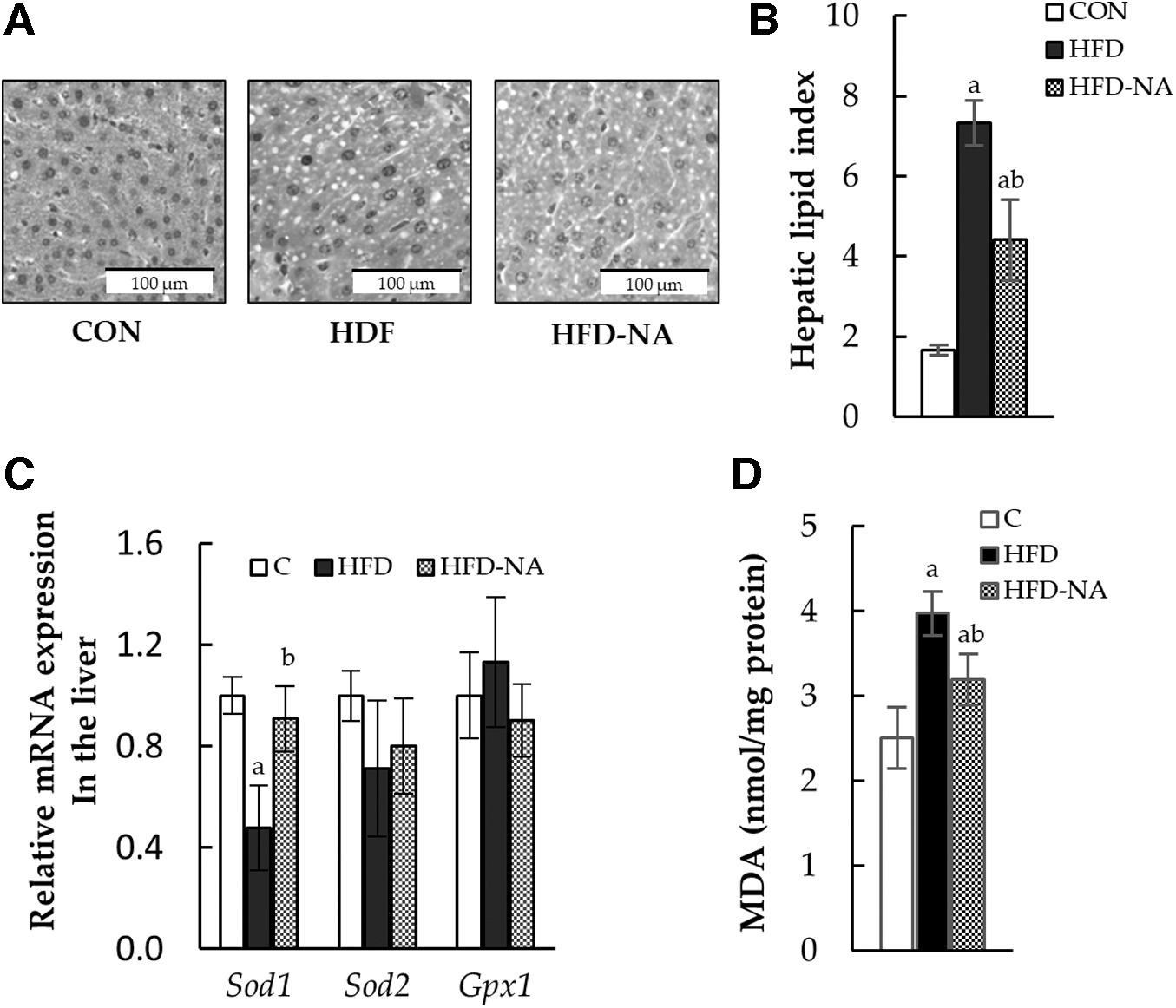
Histological lipid droplet percentage, antioxidant enzyme-coding gene mRNA expression, and MDA level in the liver.
Lipid accumulation, antioxidant gene expression, and sialylated protein levels in 3T3-L1 cells
The effects of NANA supplementation on adipocytes were investigated using 3T3-L1 cells (Fig. 5A). NANA did not exert cytotoxicity at any of the doses tested, and the cell viability remained at ∼90–100% in the nontoxic range (Fig. 5B). There were no changes in lipid accumulation in 3T3-L1 cells when supplemented with NANA at 0.1–10 μg/mL, as observed using Oil Red O staining (Fig. 6C, D).

Effect of NANA supplementation on 3T3-L1 adipocytes.
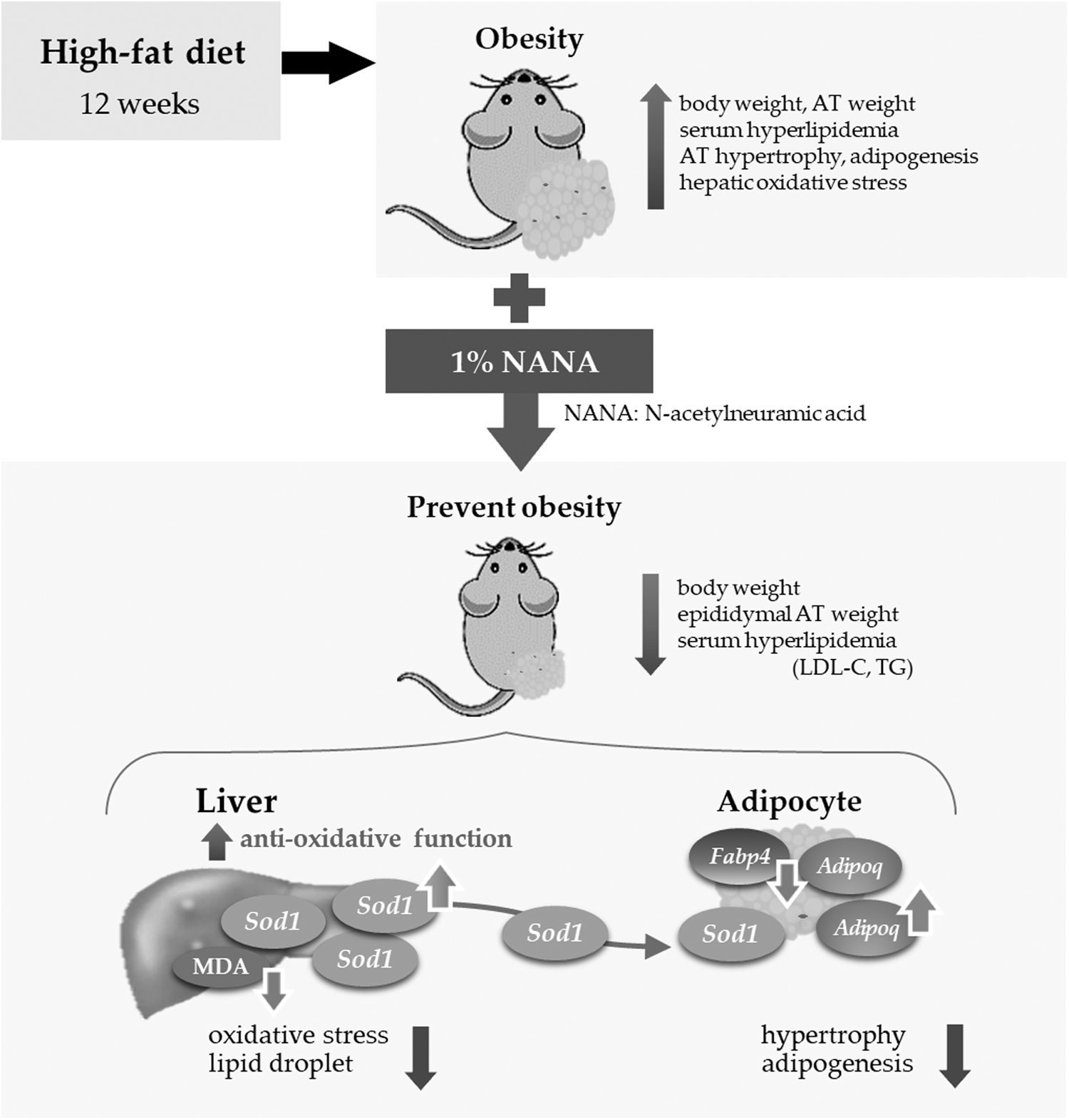
Schematic of the effect of NANA on obesity. By supplementing NANA, body weight and epididymal AT weight changes, and serum hyperlipidemia were prevented. These effects of NANA might be through enhanced hepatic antioxidant capacity upon Sod1 upregulation.
DISCUSSION
To the best of our knowledge, this the first study to demonstrate the antiobesity effect of NANA supplementation, which attenuates bodyweight gain, adipocyte hypertrophy, hyperlipidemia, and hyperglycemia in HFD-fed mice. Our results indicated that the antioxidant properties of NANA may have resulted in the alleviation of obesity, which could prevent obesity-related complications including diseases such as diabetes mellites, hypertension, asteatosis, and hepatic lipidemia.
Diet-derived NANA is considered to function through certain pathways in vivo, such as antioxidant action 8,20,21 and the conversion of NANA compounds to sialylated proteins. 20,22,23 Therefore, to elucidate the mechanism by which NANA exerts antiobesity effects, we examined the antioxidant capacity and sialylation in adipose and liver tissues.
In the epididymal ATs, NANA supplementation ameliorated the upregulated expression of Fabp4 and the downregulated expression of Adipoq, an adipokine dysfunction known to be a typical symptom of obesity. Fabp4, which is a fatty acid-binding protein specifically expressed in adipocytes, promotes lipid synthesis and induces insulin resistance. 16,17 Adipoq has an important role in suppressing obesity-related diseases by controlling insulin resistance, hypertension, hyperlipidemia, and asteorosis. 2,4,5,16,19
These results indicate that the improvement in serum lipid and glucose levels in mice supplemented with NANA might be regulated by the expression of these adipokines. In addition, the expression of Pparg and Cebpb, the genes involved in adipocyte differentiation and upregulated upon HFD intake, was not significantly affected by NANA. Sod1 expression and the MDA level, which was measured to evaluate the production of lipid peroxides, were also not affected by NANA supplementation in epididymal adipocytes (Fig. 2C). Obesity and obesity-related diseases are regulated by glycation. 24,25
In visceral ATs, the downregulation of sialic glycation is involved in the dysfunction caused by obesity. 6 Hence, in this study, we investigated the expression of sialylated proteins and St6gal1 in epididymal ATs, to determine whether the antiobesity effects of NANA supplementation were due to increased sialylation (Fig. 5). However, the expression remained unaltered.
Through in vitro assays using 3T3-L1 cells, we also investigated antioxidant gene expression and sialylated protein levels under treatment with 1 μg/mL NANA; the NANA concentration was within the same range as human serum sialic acid concentration (0.1–2.0 μg/mL). 26,27 The results were similar to those of the mouse epididymal adipocytes, that is, the expression of Sod1 and levels of MDA, sialylated proteins, and st6gal were not affected by NANA supplementation (Fig. 5E–G). Orally administered sialic acid is excreted at a high rate (>70% within 30–60 min), and is almost completely excreted through urine within 6 h. 22,27,28
In this in vivo animal study, 1% NANA was mixed with HFD and orally administered; therefore, the serum NANA level was high, approximately at the same level as that used in in vitro assays. Both in vitro and in vivo NANA supplementation experiments indicated that adipocyte differentiation and adipocyte hypertrophy were not induced by the direct antioxidant activity, and there was an increase in NANA sialylation levels in adipocytes. In the liver, NANA supplementation attenuated lipid droplet accumulation and notably inhibited oxidative stress by altering the expression of Sod1 and MDA levels, which were the same as those in CON mice (Fig. 4).
Liu et al. reported that in obese mice overexpressing Sod1, lipid accumulation decreases and insulin sensitivity increases. 29 Our observations indicate that the same mechanism might have been involved and that Sod1 upregulation by NANA supplementation regulates hepatic lipid metabolism and insulin sensitivity. We further speculate that this caused the reduction in hepatic lipogenesis and serum glucose level. As obesity increases oxidative stress, which induces obesity-related complications, 3,30 NANA could be effective in preventing obesity-related complications by upregulating Sod1 expression.
Anti-inflammatory effects of NANA on the aorta and liver have been reported. 21,31 The anti-inflammatory effect of NANA on adipocytes remains poorly understood, although hepatic lipid level, hypertrophy, and adipocytokine may have been regulated through the anti-inflammatory effect. The decrease in serum AST level with NANA supplementation also indicates an inhibitory effect on liver inflammation. However, we did not measure inflammatory markers such as tumor necrosis factor-α, warranting further investigation.
The antiobesity effect of NANA might be related to the antioxidant activity of the liver and adipocytokine regulation; however, as serum sialic acid is rapidly excreted in urine, 32 high sialic acid levels, as observed in this study, may be difficult to maintain. To elucidate this mechanism in more detail, it is necessary to consider the markers of inflammation and other antioxidants (e.g., GSH/GSSG levels), and the effect of NANA supplementation on hepatic sialylation and on other organs.
This study demonstrated that NANA supplementation has an antiobesity effect, suppressing body weight gain; reducing AT fat and hepatic lipid accumulation; and reducing TC, LDL, TG, AST, and fasting glucose levels. These effects were found to be related to the antioxidative effect of NANA. NANA has potential to be developed as a safe functional food that prevents and alleviates obesity.
Footnotes
ACKNOWLEDGMENTS
The authors thank Mr. Shida and other members of the Nutrition Laboratory, Daito Bunka University, for their support in this study. They are grateful to Dr. Fukushima for helpful discussions and comments on the article. They thank Editage (
AUTHOR CONTRIBUTIONS
T.K. conceived and designed the experiments, wrote the article, and acquired funding; T.K. and Y.O. performed the experiments; T.K. analyzed the data; and S.O. supervised the study.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This study was supported by the Japan Society for the Promotion of Science Grant-in-Aid for Scientific Research (C) 18K11138 (to T.K.).