
Abstract
Hyperhomocysteinemia is a main risk factor for phenotypic modulation of vascular smooth muscle cells (VSMCs) and atherosclerosis. Phenotypic switching and proliferation of VSMCs are related to the progression of vascular inflammation. Chrysanthemum coronarium L. is a leafy vegetable with various biological functions, such as antioxidative, anti-inflammatory, and antiproliferative effects. In this study, we aimed to identify the mechanisms underlying the therapeutic and preventive effects of C. coronarium L. extract (CC) in regulating homocysteine (Hcy)-induced vascular inflammation in human aortic VSMCs. CC did not exhibit cytotoxicity and inhibited Hcy-stimulated VSMC proliferation and migration. In addition, CC promoted Hcy-induced expression of VSMC contractile phenotype proteins, including alpha-smooth muscle actin, calponin, and smooth muscle 22α. CC also decreased Hcy-induced accumulation of reactive oxygen species and expression of inflammatory markers nicotinamide adenine dinucleotide phosphate oxidase-4 and soluble epoxide hydrolase. These results showed that CC attenuates Hcy-induced inflammatory responses, highlighting its potential as a therapeutic or preventive target for Hcy-induced vascular inflammation.
INTRODUCTION
Atherosclerosis (AS) is a slow progressive cardiovascular disease correlated with high mortality rates worldwide. 1 Vascular smooth muscle cells (VSMCs) are related involved in AS pathophysiology. 2 In inflammatory stimuli or oxidative stress conditions, VSMCs can change to a de-differentiated, proliferative, and migratory phenotype, that is, a contractile to synthetic shift, by suppressing their contractility, thereby promoting proliferation and migration. 3,4 This phenotype switching is associated with excessive proliferation and migration of VSMCs, which is a major factor in the pathological process of AS. 5,6 Although increasing evidence has revealed the phenotypic modulation of VSMCs, the mechanisms underlying this phenotypic transition and its modulation in AS are unknown.
Homocysteine (Hcy) is an amino acid metabolite produced from methionine. Hyperhomocysteinemia (HHcy) is defined as an elevated blood Hcy concentration exceeding 15 μmol/L. For every 5 μmol/L increment in blood Hcy concentrations, there is an independent risk of a 500 μmol/L increase in cholesterol levels, and the cardiovascular disease risk is increased by ∼1/3. 7 Therefore, HHcy is a high-risk factor for AS. 8 Moreover, VSMCs are the major cells of the artery and are converted to foam cells in AS plaques.
Accumulating evidence shows that Hcy-induced oxidative stress, endothelial dysfunction, and VSMC migration and proliferation are associated with the progression of AS. 9 –11 Moreover, HHcy increases VSMC phenotype modulation by increasing VSMC proliferation and inflammation, ultimately promoting the development of AS. 4 Therefore, given their role in the development of AS, understanding the molecular mechanisms underlying Hcy-stimulated VSMC conversion is important. Although a few strategies have been shown to suppress VSMC proliferation, migration, and phenotype conversion in AS, no effective therapy has been established yet.
Glebionis coronaria (Chrysanthemum coronarium L., crown daisy), called “ssukgat” in Korea, is an annual herbaceous weed widely distributed in the Mediterranean region. It is a valuable vegetable and a medicinal and ornamental plant. C. coronarium L. extract (CC) has various biological functions, including antioxidative, antihypercholesterolemic, and antihyperglycemic activities. 12 –14 CC also exhibits anti-inflammatory and antiproliferative activities in vitro. 12,15
We have previously found that CC has preventive properties on osteoporotic bone loss. 16 However, the mechanisms underlying Hcy-mediated suppression of VSMC proliferation, migration, and phenotype modulation remain unexplored. In this study, we hypothesized that CC exhibits inhibitory effects on Hcy-induced vascular inflammation and discuss the underlying mechanisms.
MATERIALS AND METHODS
Chemicals and reagents
Human aortic VSMCs (HA-VSMCs) and Dulbecco's modified Eagle's medium (DMEM) were provided from ATCC (Manassas, VA, USA). Fetal bovine serum (FBS), penicillin, streptomycin, bicinchoninic acid (BCA) assay kit, and CellROX® Deep Red Oxidative Stress reagent were provided from Gibco (Thermo Fisher Scientific, Waltham, MA, USA). Hcy, 2′,7′-dichlorofluorescein diacetate (DCFH-DA), and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) solutions were obtained from Sigma-Aldrich (St. Louis, MO, USA).
Bromodeoxyuridine (BrdU) was provided from Millipore (Burlington, MA, USA). A transwell chamber assay (8 μm pore size) was purchased from Costar (Corning, Inc., Corning, NY, USA). Radioimmunoprecipitation assay (RIPA) buffer was obtained from Beyotime (Los Altos, CA, USA). Anti-α-smooth muscle actin (α-SMA), anti-calponin, anti-smooth muscle-α, and reactive oxygen species (ROS) assay kits were provided from Abcam (Cambridge, UK).
Intercellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule (VCAM)-1 were provided from Santa Cruz Biotechnology (Dallas, TX, USA). Nicotinamide adenine dinucleotide phosphate oxidase-4 (Nox4) was obtained from OriGene (Rockville, MD, USA). Soluble epoxide hydrolase (sEH) was provided by Cayman (Ann Arbor, MI, USA). β-actin and secondary antibodies were purchased by Cell Signaling Technology (Danvers, MA, USA).
Preparation of CC
The air-dried plants (1 kg, traditional Korean market, Jeonju) were washed and extracted once with 9 volumes of 50% ethanol for 24 h. CC was filtered and evaporated by freeze-drying at −70°C. The recovery yield was ∼8% (w/w).
Cell culture and cell viability
HA-VSMCs were maintained in DMEM with 10% FBS at 37°C in a 5% CO2. Cells were seeded at 3000 per well of a 96-well plate and pretreated with several CC concentrations (10, 25, 50, and 100 μg/mL) for 24 h. MTT solution (0.5 mg/mL) was added to VSMCs for 2 h and the signal was quantified in absorbance at 450 nm using a microplate reader (Molecular Devices, San Jose, CA, USA).
Cell proliferation
The BrdU cell proliferation assay was used to detect VSMC proliferation. VSMCs were seeded in 96-well plates and pretreated with the indicated CC d (10, 25, and 50 μg/mL) with or without Hcy for 24 h. The BrdU assay was estimated following the manufacturer's protocols to analyze the cell proliferation.
Scratch wound-healing assay
VSMCs were seeded at 300,000 per well of a six-well plate. Next, cells scratched with a 200 μm sterile tip and VSMCs were incubated with the indicated CC concentrations with or without Hcy. Photographs were taken using a microscope (Leica, Wetzlar, Germany) at 0 and 24 h. The wound healing was measured based on the wound area at 24 h compared with 0 h.
Transwell migration assay
For the transwell migration assay, VSMCs were cultured in the upper surface of the membrane with 8 μm-pore membranes (Corning) and incubated with several CC concentrations with or without Hcy for 24 h. VSMCs that moved to the lower surface were fixed with 4% paraformaldehyde and stained with 0.1% crystal violet. The migrated VSMCs was analyzed under a light microscope. The mean was calculated from five random visual fields.
ROS measurement
To assess intracellular ROS generation, VSMCs were maintained with several CC concentrations and Hcy for 24 h. VSMCs were treated with 10 μM DCFH-DA at 37°C in the dark for 30 min. The fluorescence signals were detected using a fluorescence reader (Molecular Devices). Images were taken with a fluorescence microscope (Olympus Corporation, Tokyo, Japan). In addition, after incubation with the CC with or without Hcy for 24 h, HA-VSMCs were maintained with CellROX Deep Red Oxidative Stress reagent for 30 min. Cells were then washed and fluorescence signals were analyzed at 640/665 nm with a microplate reader (Molecular Devices).
Immunoblotting
Total protein was extracted from lysed cells in RIPA buffer. We measured the concentration of protein using a BCA assay. Proteins were separated by sodium dodecyl-sulfate polyacrylamide gel electrophoresis, transferred to polyvinylidene difluoride membranes. Blots were immunoprobed with specific primary antibodies against anti-α-SMA, anti-calponin, anti-SM22-α, anti-ICAM-1, anti-VCAM-1, anti-Nox4, anti-sEH, and anti-β-actin overnight. The blots were then probed with horseradish peroxidase-conjugated secondary antibodies. Signals were analyzed using the ChemiDoc™ Imaging System (Bio-Rad Laboratories, Hercules, CA, USA). Densities of specific bands were estimated using Image J software.
Quantitative reverse-transcription polymerase chain reaction
Total RNA was extracted from VSMCs using TRIzol reagent and converted to cDNA. Quantitative reverse-transcription PCR (qRT-PCR) was performed using 2 × Taq polymerase chain reaction MasterMix and the iTaq universal SYBR Green I Supermix (Bio-Rad Laboratories). The thermal cycling conditions were as follows: initial denaturation at 95°C for 3 min, followed by 39 cycles of 30 s at 95°C, 30 s at 55°C, and 30 s at 72°C. The melting curve was acquired by increasing the temperature from 65°C to 95°C with a temperature transition rate of 0.1°C/s using the Bio-Rad RT-QPCR software. The internal reference gene was glyceraldehyde 3-phosphate dehydrogenase (GAPDH). The specific primers used are provided in Table 1.
List of Primers Used in the Study
GAPDH, glyceraldehyde 3-phosphate dehydrogenase; IL, interleukin; MCP-1, monocyte chemoattractant protein-1; TNF-α, tumor necrosis factor-alpha.
Statistical analysis
Data are shown as the means ± standard deviation. Statistical analysis was determined using GraphPad Prism software version 10.0 (GraphPad, Inc., La Jolla, CA, USA). Statistical differences were determined using one-way analysis of variance. P < .05 was regarded as significant.
RESULTS
CC inhibits Hcy-induced VSMC proliferation
To evaluate the cytotoxic effect of CC on VSMCs, we assessed the cell viability after treatment with various CC concentrations using the MTT assay. Compared with control cells, treatment with the CC (10, 25, 50, and 100 μg/mL) for 24 h did not show cytotoxicity in HA-VSMCs (Fig. 1A). Moreover, the effect of the CC on VSMC proliferation was assessed using the BrdU assay; HA-VSMCs were pretreated with the CC at different dosages (10, 25, and 50 μg/mL) and then with Hcy. We found that Hcy significantly induced VSMC proliferation. However, CC suppressed Hcy-induced VSMC proliferation in a concentration-dependent manner (Fig. 1B).

CC inhibits Hcy-induced VSMC proliferation. Cells were pretreated with CC at different concentrations (10, 25, and 50 μg/mL) for 30 min and then with Hcy for 24 h.
CC inhibits the Hcy-induced VSMC migration
To evaluate the effect of CC on VSMC migration, HA-VSMCs were incubated with the CC (10, 25, and 50 μg/mL) and then with Hcy. Wound-healing and transwell assays were performed. Our results showed that Hcy significantly promoted VSMC migration. Although treatment with 10 μg/mL of CC did not inhibit VSMC migration, 25 and 50 μg/mL CC inhibited migration in a dosage-dependent manner (Fig. 2). These findings show that the CC suppresses Hcy-induced VSMC migration.

CC inhibits Hcy-induced VSMC migration. Cells were pretreated with CC at different concentrations (10, 25, and 50 μg/mL) for 30 min and then with Hcy for 24 h.
CC abrogates Hcy-induced expression of VSMC contractile markers
To identify the effect of the CC on VSMC differentiation, VSMCs were pretreated with the CC (10, 25, and 50 μg/mL) and then with Hcy, and the levels of contractile phenotype proteins, including α-SMA, smooth muscle 22α (SM22α), and calponin, were assessed. Hcy-triggered VSMC phenotype modulation from a differentiated to a de-differentiated condition, shown by decreased α-SMA, calponin, and SM22-α expression, whereas these effects were reversed by the CC (Fig. 3). These findings indicate that the CC ameliorates Hcy-stimulated VSMC differentiation.
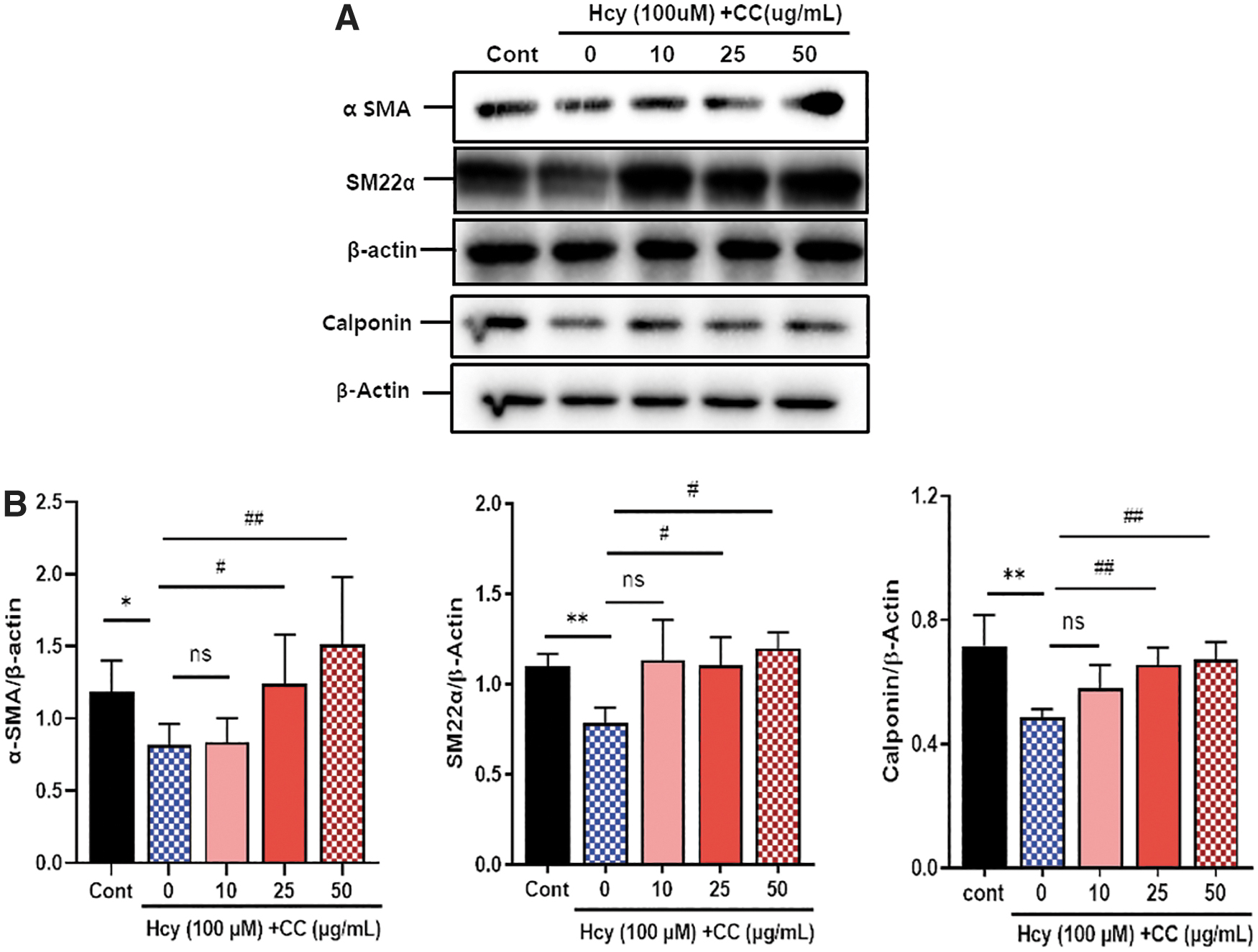
CC abrogates Hcy-induced VSMC differentiation. The cells were pretreated with the CC at different concentrations (10, 25, and 50 μg/mL) for 30 min and then with Hcy for 24 h.
CC decreases Hcy-induced ROS production
To assess the effect of the CC on ROS production, HA-VSMCs were incubated with the CC (10, 25, and 50 μg/mL) and then with Hcy, and ROS levels were assessed using DCFH-DA staining and CellROX Deep Red Oxidative Stress reagent. Hcy-treated cells exhibited significantly higher levels of DCFH-DA fluorescence (P < .001) and CellROX Deep Red Oxidative Stress staining (P < .001) compared with control cells. However, CC significantly reduced Hcy-stimulated ROS production in a dosage-dependent manner (Fig. 4). Our findings show that the CC suppressed Hcy-induced ROS production.

CC decreases Hcy-induced ROS production. Cells were pretreated with the CC at different concentrations (10, 25, and 50 μg/mL) for 30 min and then with Hcy for 24 h.
CC inhibits Hcy-stimulated inflammation and Nox4 and sEH expression
To assess the effects of the CC on Hcy-stimulated inflammation and Nox4 and sEH expression, HA-VSMCs were pretreated with the CC (10, 25, and 50 μg/mL) and then with Hcy. We analyzed the protein levels of Nox4, sEH, ICAM-1, and VCAM-1 by immune blotting and the mRNA levels of tumor necrosis factor (TNF)-α, interleukin (IL)-6, IL-1β, and monocyte chemoattractant protein-1 (MCP-1) using quantitative reverse-transcription polymerase chain reaction. Hcy significantly increased Nox4, sEH, ICAM-1, and VCAM-1 protein levels, as well as TNF-α, IL-6, IL-1β, and MCP-1 mRNA levels. However, these effects were markedly suppressed by the CC (Fig. 5). These findings show that the CC inhibits Nox4 and sEH expressions and vascular inflammation.
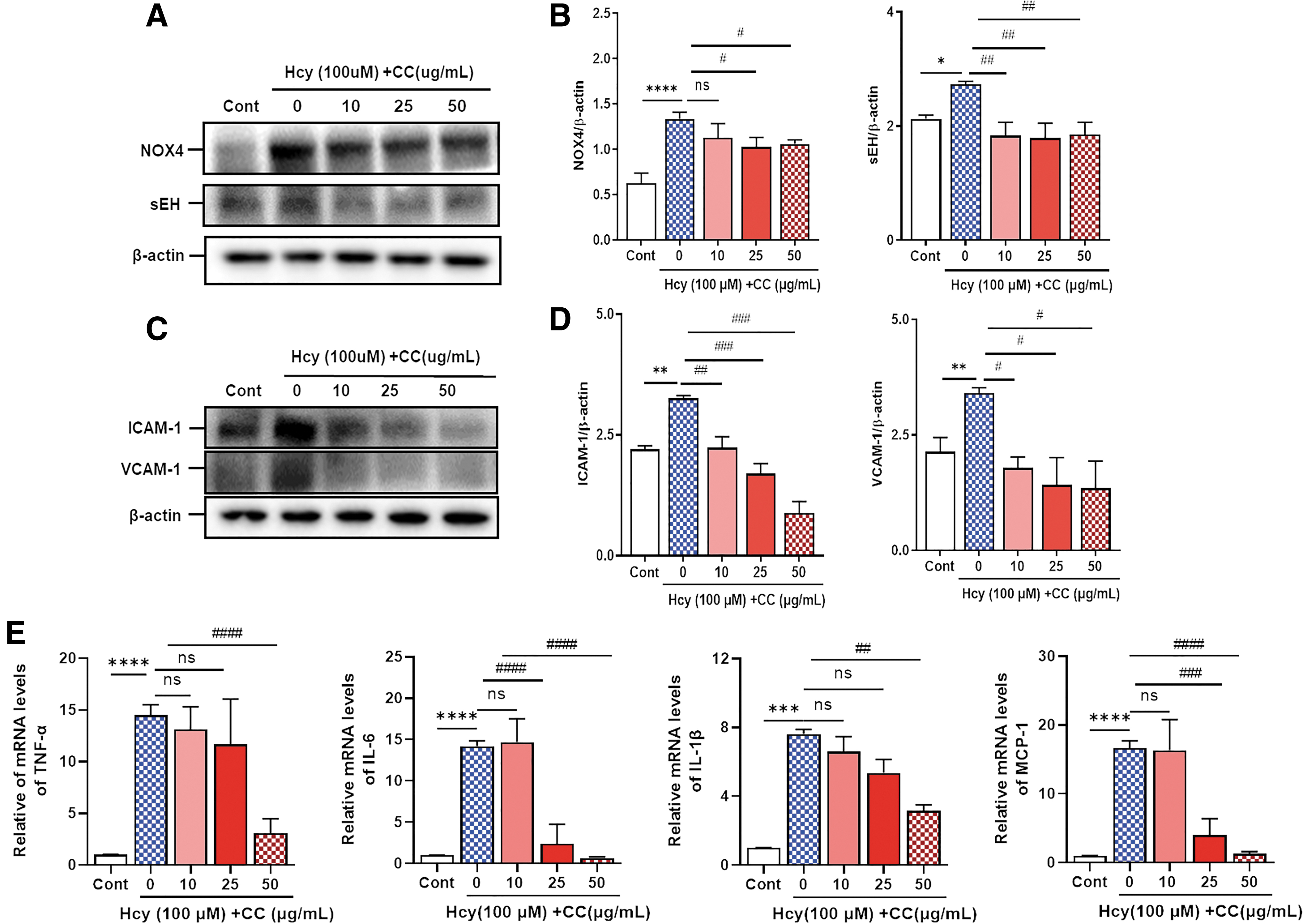
CC reduces Hcy-induced inflammation in VSMCs. Cells were pretreated with CC at different concentrations (10, 25, and 50 μg/mL) for 30 min and then with Hcy for 24 h.
DISCUSSION
AS is a slow progressive vascular disease of the arteries. There is abundant evidence that abnormal VSMC proliferation and migration are pathological events that mediate AS development 17 –19 and that the phenotypic transition of VSMCs is a requirement for their proliferation and migration. 10,20 Moreover, HHcy induces phenotypic switching of VSMCs. 21 Ji et al. 22 reported that liraglutide attenuates Hcy-triggered VSMC proliferation, migration, and phenotypic conversion in vitro. Furthermore, artemisinin attenuates the progression of AS by regulating the phenotypic change of VSMCs. 23
Thus, in this study, we focused on the suppression of VSMC phenotypic transition as a potential pharmacological target for AS in vitro. We have recently shown that 1-trifluoromethoxyphenyl-3-(1-propionylpiperidin-4-yl) urea (TPPU) attenuates the progression of vascular inflammation by inhibiting VSMC phenotypic switching in vitro and in vivo. 24 In this study, our findings suggested that the CC affected Hcy-induced proliferation, migration, and de-differentiation in HA-VSMCs, suggesting that the CC attenuates the progress of vascular inflammation by modulating VSMC phenotype conversion.
Increasing evidence indicates that AS is most commonly caused by Hcy 25 ; HHcy triggers the initial development of AS 26,27 and can stimulate VSMC proliferation and migration, which promotes VSMC inflammation. 28 Moreover, HHcy promotes phenotypic transition. Physiologically, mature VSMCs have a contractile phenotype, which is expressed by high levels of contractile markers α-SMA, SM22α, and calponin.
However, Hcy stimulates contractile VSMCs to switch to a synthetic type. 29,30 Similarly, we have previously found that HHcy promotes VSMC phenotypic transition. 24 Our findings showed that Hcy increased VSMC proliferation and migration and reduced the levels of contractile proteins, such as α-SMA, calponin, and SM22-α, whereas CC supplementation normalized these parameters.
Moreover, Liu et al. 9 suggested that Hcy stimulates vascular inflammation and increases ICAM-1 and VCAM-1 expression in VSMCs and mouse aorta. In our study, the CC inhibited Hcy-stimulated adhesion molecules, such as ICAM-1 and VCAM, and the mRNA expression of inflammatory markers, such as TNF-α, IL-6, IL-1β, and MCP-1, in VSMCs. Therefore, our findings indicate that the effects of CC may be related to the suppression of vascular inflammation.
ROS plays a role in promoting inflammation. NOX are the main factors of ROS, among which Nox4 is required for the phenotypic switching of VSMCs. 29,30 Moreover, epoxyeicosatrienoic acids (EETs) have protective effects against vascular diseases, and sEH, which metabolizes EETs to less active diols, is associated with VSMC phenotype modulation. 24 Therefore, several lines of evidence show that sEH inhibition can suppress the progression of vascular inflammation in vivo and VSMCs.
In addition, Hcy stimulates oxidative stress in VSMCs; Hcy-induced intercellular ROS is mediated by modulating the proliferation, migration, and phenotype switching of VSMCs and is associated with vascular inflammation. Liu et al. 9 have reported total ROS and H2O2 production in Hcy-treated VSMCs, showing that ROS neutralization can abolish the effects of Hcy on VSMCs. Among ROS factors, Hcy has been known to modulate p47phox, Nox1, Nox2, and Nox4 in VSMCs. Nox4 is the main NADPH oxidase in VSMCs. 9,31
Notably, TPPU, an sEH inhibitor, can suppress Hcy-induced vascular inflammation through the Nox4 pathway. 24 Hcy also increases sEH expression, and Hcy-induced sEH regulates Nox-4-mediated ROS production, indicating that sEH and Nox4 are partially associated with Hcy-induced inflammation in VSMCs. 9,29,30 Therefore, several studies have focused on inhibiting sEH and/or Nox4 to protect against vascular inflammation. 32,33 In this study, these findings suggested that the CC inhibited Hcy-induced ROS production and sEH and Nox4 expression in VSMCs, suggesting that the CC might attenuate vascular inflammation by regulating Nox4 and sEH expression.
Epidemiological evidence has indicated that the intake of vegetables, plants, or their derived flavonoids delays the development of AS. 12 CC contains caffeic and chlorogenic acids, 15 and in our previous study we showed that CC contains rutin, cynarin, 3,4-dicaffeoylquinic acid, and 3,5-dicaffeoyl-4-succinoyl quinic acid. 16 Rutin possess several functional effects, such as anti-inflammatory and antioxidative properties, and inhibits premature senescence, proliferation, and migration of VSMCs and in AS in vivo. 34,35 Caffeic acid has beneficial activities, including antioxidant and anti-inflammatory effects, and inhibits VSMC proliferation. 36,37 Chlorogenic acid has antioxidant, cardioprotective, and anti-inflammatory effects. 38
Furthermore, 3,4-dicaffeoylquinic acid reduces VSMC proliferation and migration. 39 Therefore, we investigated if the CC, which contains various bioactive compounds, would possess a suppressive activity on Hcy-stimulated inflammation in VSMCs. Herein, treatment with CC suppressed VSMC proliferation, migration, and phenotypic modulation during Hcy-induced inflammation, suggesting that the CC may exert a potential therapeutic effect for vascular inflammation-associated AS.
There are some limiting factors in this study. First, this study did not determine the effect of the CC on VSMC proliferation, migration, and phenotype switching in vivo. Second, this study did not define the preventive effects of several CC-derived compounds. Third, further studies will be required to investigate the mechanistic differences in the effects of the CC and CC-derived components.
In conclusion, our findings revealed that the CC inhibited Hcy-induced VSMC proliferation, migration, and phenotypic conversion in vitro, suggesting its potential as a therapeutic strategy for vascular inflammation-associated AS.
Footnotes
AUTHORs' CONTRIBUTIONS
Conceptualization, writing—review and editing, and funding acquisition by M.J.S. Methodology and writing—original draft by Y.K. and M.J.S. Investigation by A.S.L. and Y.K. Visualization by H.J.H. and S.H.L. Software by H.J.H.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This study was supported by a grant from the Korea Food Research Institute (E0210102-03).