
Abstract
Alcoholic liver disease (ALD) is damage to the liver and mainly caused by binge alcohol. ALD have decreased junctional protein expression and modulated intestinal permeability. We investigated whether plant-releasing exosome-like nanovesicles can prevent liver damage and leaky gut from binge alcohol. In this study, we characterized the exosome-like nanovesicles from pomegranate juice and confirmed the round shape of a lipid bilayer. After 14 days of pomegranate-derived exosome-like nanovesicle (PNVs) pretreatment, binge alcohol (6 g/kg/dose) was administered to mice three times orally every 12 h. Exposure to binge alcohol increased levels of oxidative and nitric oxide stress marker proteins such as CYP2E1, 3-Nitrotyrosine, and inducible nitric oxide synthase in both liver and gut damage. Also, binge alcohol significantly elevated the plasma endotoxemia, inflammatory fatty liver, and leaky gut. However, PNVs reduced the oxidative stress and apoptosis marker proteins and prevented the leaky gut and endotoxemia. Markedly, PNV treatment significantly prevented a decrease in the amount of intestinal junctional proteins and an increase in leaky gut in mice exposed to alcohol. These results showed that PNVs can prevent leaky gut and liver damage caused by binge alcohol and suggest that it may be useful hepatoprotective or intestinal protective agents for the first time.
Introduction
Alcoholic liver disease (ALD) is caused by chronic and/or binge drinking. 1 Binge drinking is defined as a drinking pattern that results in a blood alcohol concentration of 0.08% or more. 2 Alcohol metabolism penetrates almost every tissue in the body through an inflammatory cascade containing oxidative stress, causing significant changes in liver or gut damage. 1 The clinical spectrum of ALD includes alcoholic fatty liver disease (steatosis), steatohepatitis, fibrosis/cirrhosis, and finally liver cancer. 3
Alcohol and metabolites directly damage intestinal cells through oxidative stress and interfere with junctional proteins, thereby increasing the permeability of the intestinal barrier, affecting gut microbiota. 4 Increased bacteria can trigger an inflammatory response through endotoxins, increase lipopolysaccharide (LPS) levels due to the release of endotoxins and cytokines, and interact with toll-like receptor-4 (TLR-4) to cause liver inflammation and liver fibrosis. 4,5 It has been proven that dysbiosis and disturbances of the integrity of the intestinal barrier play a role in promoting inflammatory liver damage. 6 Therefore, leaky gut induced by alcohol is very important because it progresses to severe ALD. 7 –10
Plant-releasing exosome-like nanovesicles (PENs) exist the round shape of a lipid bilayer, including lipids, miRNAs, proteins, mRNAs, and second metabolites. 11 –13 PENs have the characteristics of a nano size, stability and low toxicity, offering a potential protective role in liver and gut diseases, and a therapeutic alternative to various diseases as drug carriers. 8,9,12 –18 Also, PENs have intercellular communication, and anti-inflammatory, immunomodulatory, and regenerative properties through the regulation of protein expression. 11 –13 Recently, our group reported antiosteoporosis effect of yam-derived nanovesicles or apple-derived nanovesicles through modulation of osteoblast differentiation. 12,19 Indeed, PENs isolated from ginger, grapefruit, and grapes have been demonstrated to have positive effects on alcohol-induced liver damage by stimulating antioxidants and anti-inflammation. 11,15,16
Pomegranate is a polyphenol-rich fruits with high antioxidant activity. 20 Phytochemicals extracted from pomegranate and pomegranate juice have various bioactive properties such as antioxidant, anti-inflammatory, and antiobesity. 21 –23 Pomegranate juice contains antioxidants such as polyphenols (0.2–1%), tannins, punic acid, and ascorbic acid. 21,23 Especially, the most abundant phenolic compound in pomegranate extract is ellagic acid (EA), which has very high bioavailability and bioactivity at higher levels than free EA. 13,24 In fact, our studies have reported that pomegranate (contained 40% EA) alleviated binge alcohol-induced gut leakiness and liver inflammation. 8 Various bioactive ingredients extracted from pomegranate juice act as antioxidant and anti-inflammatory, and are mentioned as beneficial treatments for liver- and gut-related diseases. 8,10 However, the effects of pomegranate-derived exosome-like nanovesicles (PNVs) on alcohol-induced diseases of liver and gut are not known.
In this study, we successfully isolated and characterized the exosome-like nanovesicles from pomegranate juice and confirmed nano size and lipid layer image of PNVs. Also, we found that PNVs prevent binge alcohol-induced leaky gut and inflammatory liver disease in mice and in vitro (Fig. 1). Therefore, this study showed PNVs can be a beneficial agent for diseases associated with liver inflammation and gut leakiness by binge alcohol.
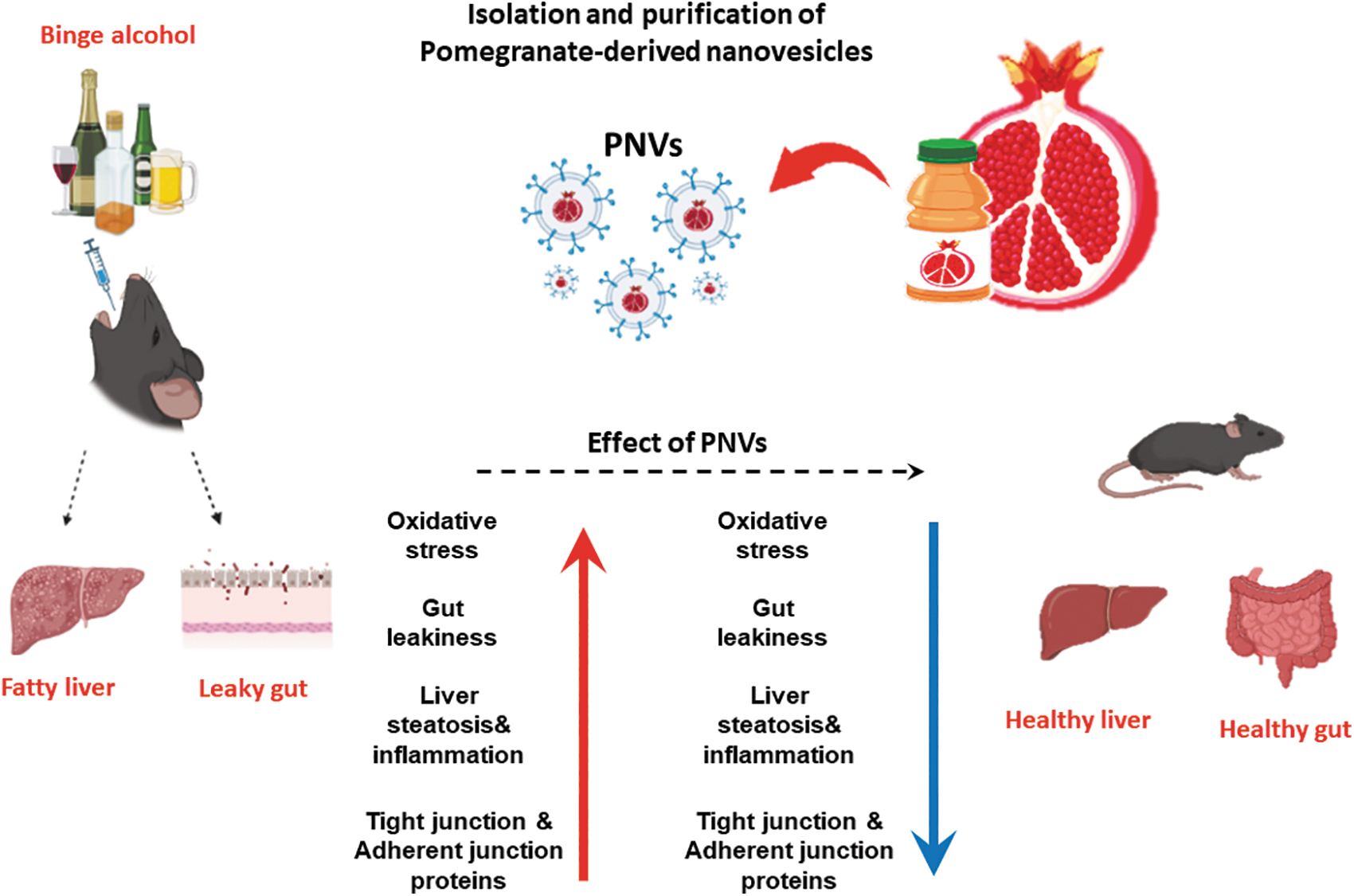
Summary of preventive effects of PNVs against binge alcohol-induced oxidative stress, gut leakiness, and apoptosis through liver-gut axis. The up (red) and down (green) arrows represent an increment by binge alcohol exposure and a decrement of each indicated parameter, respectively, by PNV pretreatment. PNVs, pomegranate-derived exosome-like nanovesicles.
Materials and Methods
Isolation of exosome-like nanovesicles from pomegranate
PNVs were isolated from the fresh pomegranate juice (Geroria's Natural, Tbilisi, GA, USA). 13 Nanovesicles were isolated according to our laboratory method. 12,17 Each exosome pellet was dissolved in phosphate buffered saline (PBS) and characterized nano size. 12,17
Nano-size conformation
The number and size of PNVs were measured by nanoparticle tracking analysis (NTA) using a NanoSight NS300 system (NanoSight, Amesbury, UK). For the transmitter electron microscopy (TEM) analysis, samples were filled into glow-discharge copper grids coated with a continuous carbon film and stained with 0.75% uranyl formate. 25
Animal experiments
All the animal studies were approved and conducted by the guidelines of the Institutional Animal Care and Use Committee (IACUC) of the Andong National University (2019-2-0510-02-01). All female mice were controlled Andong University Animal Care guideline. The experiment was conducted by dividing 6-week-old female mice (C57BL/6J) into EtOH+PNVs (1 mg/kg) groups (n ≥ 7), Control groups (water; n ≥ 7), or EtOH groups (6 g/kg; n ≥ 7). The EtOH+PNVs groups were orally administered 1 mg/kg of PNVs every day during 2 weeks for pretreatment. After PNV oral administration, female mice were exposed to 3 oral doses of binge alcohol (6 g/kg/dose) or dextrose (as control) at 12-h intervals and euthanized 1 h after the last ethanol dose, as described. 8,10,26 PNV dose was set 1 mg/kg to mimic the regenerative activity dose of plant-derived exosome-like nanovesicles. Female mice were chosen since they are more susceptible to alcohol-induced liver and gut injury than male mice. 27,28
Cell culture
The AML12 normal liver cells were cultured in DMEM/F-12 medium at 37°C in a 5% CO2 atmosphere. We treated 100 mM ethanol with PNVs (1 mg/kg) into AML12 cells for 24 h.
T84 colon cells were grown in a humidified incubator under 95% air and 5% CO2 at 37°C in Ham's F-12 medium. And then, we treated 100 mM ethanol with PNVs (1 mg/kg) into AML12 cells for 24 h.
Histological analysis
Liver or small intestine tissue was fixed in formalin and embedded in paraffin. And then, tissues were stained with hematoxylin and eosin (H&E) solution (Kyungpook National University core lab). 29 Histological changes was measured under list microscope. 29
Plasma alanine transaminase, triglyceride, and endotoxin measurement
The plasma alanine transaminase (ALT) and hepatic triglyceride (TG) were detected using the commercial detection kits such as standard end-point colorimetric assay kit (BioVision, Milpitas, CA, USA, and Asan Co., Ltd., Gimpo, Korea). Plasma endotoxin levels were detected using the commercial detection kits (endpoint LAL Chromogenic Endotoxin Quantitation Kit; Thermo Fisher Scientific, Waltham, MA, USA). 8,30
Quantitative real-time polymerase chain reaction analysis
Transcription rates of inflammation-related marker genes were measured by quantitative real-time polymerase chain reaction (PCR). Total RNA was isolated using an RNeasy Mini Kit (Qiagen, Valencia, CA, USA) and reverse-transcribed using a High-Capacity cDNA Reverse Transcription Kit (Applied Biosystems, Foster City, CA, USA) following manufacturer's protocol. Real-time quantitative PCR was carried out using the standard protocols using a fluorescence dye SYBR Master Mix Kit (Applied Biosystems), a QuantStudio™ 1 real-time PCR instrument (Thermo Fisher), and its corresponding software (Applied Biosystems). Inflammation-related marker genes were normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Primer sequences were as follows: inducible nitric oxide synthase (iNOS) (Forward) CGGTGCTGTATTTCCTTACGAGGCCAA, iNOS (Reverse) GGTGCTGTCTGTTAGGAGGTCAAGTAA, cyclooxygenase-2 (COX-2) (Forward) GGAGAGATTACAAGATACTGATC, COX-2 (Reverse) ATGGTCAGTAGACTTTTACAGCTC, TLR-4 (Forward) AGCCATTGCTGCCAACATCA, TLR-4 (Reverse) GCTGCCTCAGCAGGACTTC, GAPDH (Forward) TGACGTGCCGCCTGGAGAAA, and GAPDH (Reverse) AGTGTAGCCCAAGATGCCCTTCAG.
Immunoblot analysis
Tissues and cells were homogenized with RIPA buffer using homogenizer. Lysates of tissue and cells were separated by sodium dodecyl-sulfate polyacrylamide gel electrophoresis and transferred to nitrocellulose membranes. These membranes were incubated with CYP2E1 (1:5000 dilution; Abcam), 3-Nitrotyrosine (3-NT) (1:5000 dilution; Abcam), Cleaved caspase-3 (1:1000 dilution; Cell Signaling), p-JNK (1:1000 dilution; Santa Cruz Biotechnology), iNOS (1:5000 dilution; Abcam), ZO-1 (1:5000 dilution; Abcam), claudin-1 (1:1000 dilution; Santa Cruz Biotechnology), claudin-4 (1:1000 dilution; Santa Cruz Biotechnology), occludin (1:1000 dilution; Santa Cruz Biotechnology), JNK (1:1000 dilution; Santa Cruz Biotechnology), BAX (1:1000 dilution; Santa Cruz Biotechnology), Bcl-2 (1:1000 dilution; Santa Cruz Biotechnology), β-catenin (1:1000 dilution; Santa Cruz Biotechnology), E-cadherin (1:1000 dilution; Santa Cruz Biotechnology), α-tubulin (1:1000 dilution; Santa Cruz Biotechnology), and GAPDH (1:1000 dilution; Santa Cruz Biotechnology) corresponding to anti-mouse, as indicated.
Nitrocellulose membranes were incubated with the secondary antibody. Detect images were detected using electrogenerated chemiluminescence solutions (Thermo Fishers). The intensities of immunoreactive target protein bands relative to GAPDH used as a loading control were quantified by densitometry using chemiluminescence imaging system (FUSION SOLO S, vilber, Collégien, France).
Bioluminescence imaging
To evaluate the liver and small intestine distribution of PNVs, DIR dye-labeled PNVs (1 mg/kg) were orally administrated into female mouse for 6, 12, and 24 h. PNVs (50 μg/mL PBS) were mixed with 10 μM iR;DilC18(7) (Invitrogen) in PBS and incubated at room temperature for 30 min. To remove nonlabeled PNVs, DIR-labeled PNVs were separated into pellets using an ultra-centrifuge (100,000 g). DIR dye-labeled PNVs (1 mg/mouse in 200 μL PBS) were orally administrated into female mouse for 6, 12, and 24 h. The intensity of DIR signal from different samples was measured using an in vivo imaging system (VIS; NEWTON 7.0, vilber, Collégien, France). The tissue samples were tested by H&E analysis to evaluate toxicity. 12
Immunofluorescence staining of CYP2E1
For detecting CYP2E1 by immunofluorescence, AML12 normal liver cells were initially plated onto chamber slides. The cells were fixed with paraformaldehyde and incubated with blocking solution. And then, the cells were incubated with CYP2E1 antibody overnight at 4°C. The cells were incubated with secondary antibody (alexa Fluor488-labeled anti-rabbit; Thermo Fisher Scientific). The nucleus of cells was stained with DAPI and mounting solution. Fluorescence images were collected by using a fluorescence microscope (Logos Biosystems, Inc., Anyang-si, South Korea).
Statistical analysis
Data were analyzed using SPSS (version 27.0; SPSS, Inc., Chicago, IL, USA). t-test was used to determine statistical significance (*P < .05, **P < .01, ***P < .001). 8 –10,29,30
Results
Purification and characterization of PNVs
Exosome-like nanovesicles were isolated from pomegranate juice without seed using our laboratory purification technique. 13 The morphology and size of PNVs were characterized by TEM analysis (Fig. 2A). The morphology of PNVs were showed lipid bilayer (Fig. 2A). Size distribution and concentration were measured at 7.6 × 1014 particle/mL, 161.4 ± 3.8 nm using NTA (Fig. 2B). To quantify PNVs, the protein concentration of PNVs was measured using a Bicinchoninic Acid Assay. The isolation of PNVs from 25 g of pomegranate were obtained the protein concentration of 25.73 ± 3 mg/mL (Fig. 2C). Therefore, these results determine that PNVs were nano-size nanovesicles and produced large amount of protein.
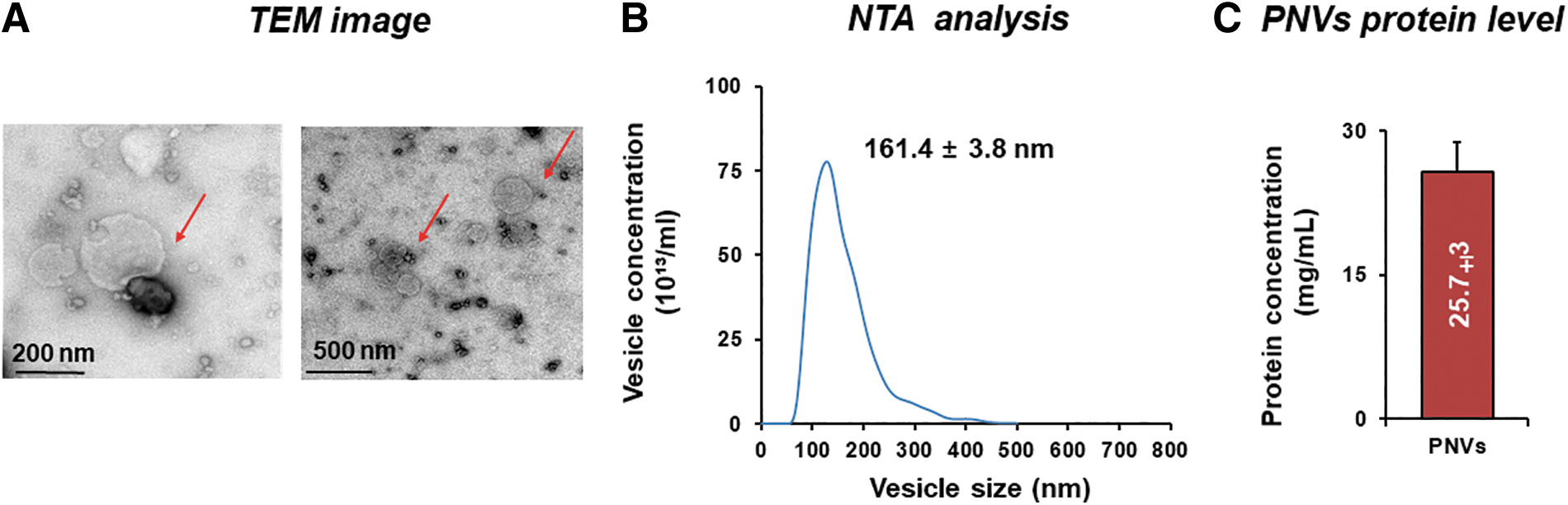
Isolation and characterization of PNVs.
In vivo distribution of orally administered PNVs and organ toxicity analysis
To evaluate the tissue distribution of orally administered PNVs, 1,1′-dioctadecyltetramethyl indotricarbocyanine iodide (DiR)-labeled PNVs were prepared and then utilized for in vivo biodistribution study using in vivo imaging system (IVIS) imaging. After oral administration, DiR fluorescent signals were predominantly detected in the gastrointestinal (GI) tract and liver in a time-dependent manner. However, fluorescent signals were not detected or were detected at a very low quantity in the lung, spleen, or heart (Fig. 3). These results indicate that PNVs can be safely transported through the GI tract and absorbed in the small intestine.

In vivo distribution of orally administered PNVs. DiR-labeled PNVs were orally administered to C57BL/6J female mice. After 6, 12, and 24 h of PNV oral administration, ex vivo images of various tissues, including small intestine, large intestine, lung, heart, liver, and spleen, were acquired with an IVIS spectrum. DiR, 1,1′-dioctadecyltetramethyl indotricarbocyanine iodide; IVIS, in vivo imaging system.
PNVs were orally administered the biological activity dose (1 mg/kg). All animals appeared to have no toxicity after oral treatments with PNVs. Oral administration of PNVs did not cause any liver, kidney, brain, heart, spleen, and lung damage by H&E staining analysis (Fig. 4). These results shown that PNVs may selectively taken up by gut and liver and can be used oral agents.

In vivo organ toxicity of PNVs. The representative images of H&E-stained liver, kidney, brain, heart, spleen, and lung sections from control (PBS) and PNV oral administrated female mice. Scale bars represent 200–100 μm. H&E, hematoxylin and eosin; PBS, phosphate buffered saline.
PNVs attenuated the intestinal oxidative stress marker proteins and alleviated junctional proteins in binge alcohol-exposed mouse
According to a recent report, binge alcohol-induced mice may lead to increased endotoxin LPS levels, which may lead to inflammatory liver disease. 8 H&E stain histology has shown that administration of PNVs prevented abnormal villus stricture caused by binge alcohol exposure (Fig. 5A). In addition, the plasma endotoxin was increased in binge alcohol exposure, but significantly decreased by the administration of PNVs (Fig. 5B).
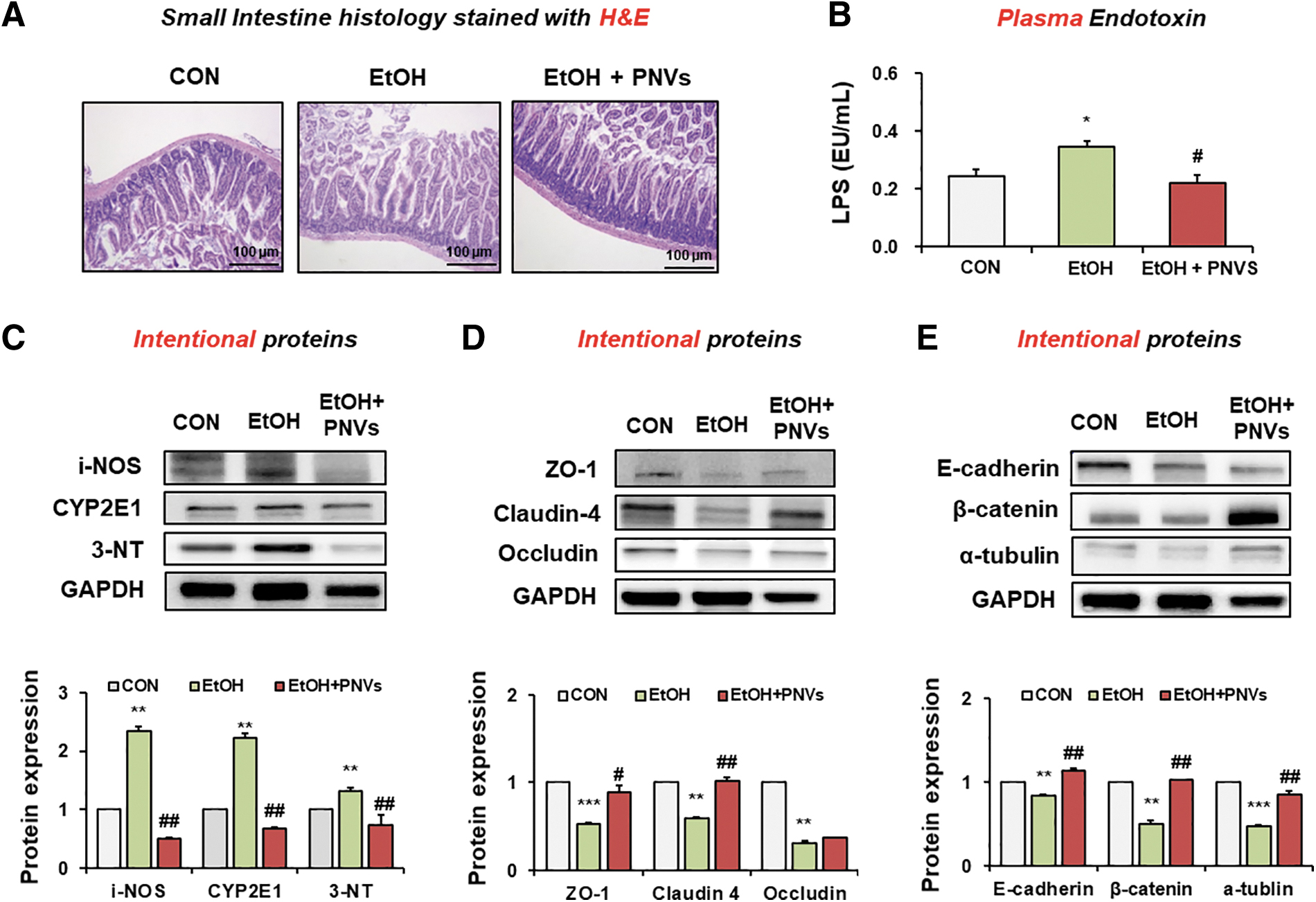
PNVs prevented leaky gut and oxidative stress in binge alcohol-exposed mouse.
Many studies have demonstrated that oxidative stress indicated by the elevated levels of CYP2E1, iNOS, and nitrated proteins promotes alcohol-mediated gut-liver damage. 8,10,29 Oxidative stress maker proteins in the small intestine were increased in alcohol-induced mice. However, PNVs significantly reduced the iNOS, CYP2E1, and 3-NT protein expressions (Fig. 5C).
Binge alcohol exposure modulated gut barrier damage by reducing junctional protein expression, which is involved in endotoxemia through oxidative streess. 1,8 The junctional proteins such as ZO-1, claudin 4, occludin, E-cadherin, β-catenin, and α-tubulin in the small intestine were also prevented in PNV treatment (Fig. 5D, E). Thus, administration of PNVs significantly reduced endotoxemia and oxidative stress maker proteins and elevated the junctional proteins by binge alcohol exposure.
PNVs enhanced epithelial barrier proteins in T84 colon cells
We confirmed that PNVs protect against alcohol-induced epithelial barrier damage even at the in vitro level. The junctional proteins such as ZO-1, claudin 1, occludin, E-cadherin, β-catenin, and α-tubulin were prevented in PNV-treated T-84 colon cells by 100 mM alcohol exposure (Fig. 6A, B). These in vitro results are consistent with the in vivo mouse results. Therefore, our results indicate that PNVs can protect the disrupted tight junction structure and prevent leaky gut.
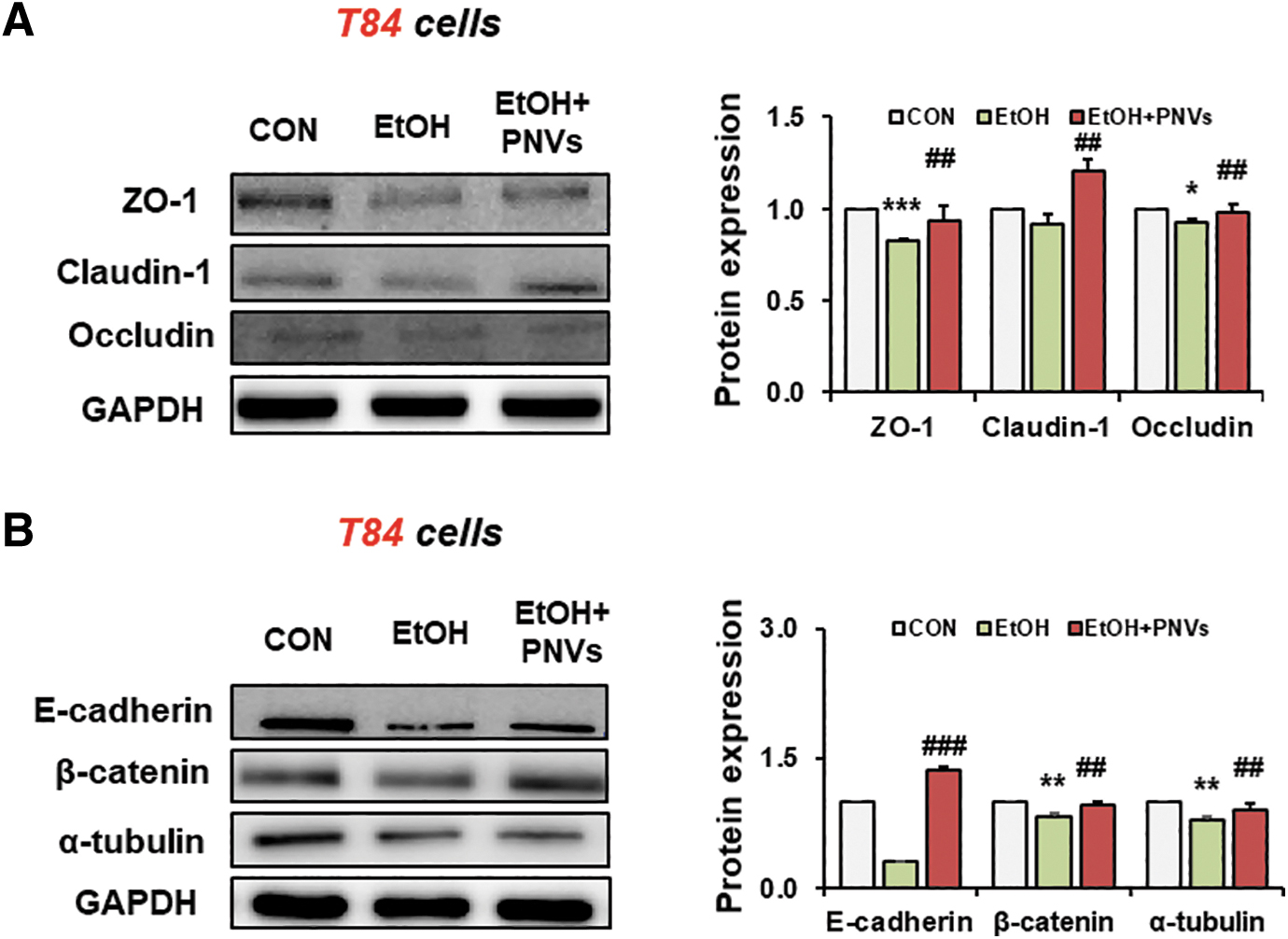
PNVs enhanced junctional proteins in T84 colonic cells.
PNVs attenuated the hepatic oxidative stress and apoptosis marker proteins in binge alcohol-exposed mouse
To confirm the hepatoprotective effect following PNVs against liver damage in alcohol-induced mouse model, H&E-stained histological analysis showed fat accumulation and liver damage in the EtOH group compared to the control group. However, administration of PNVs reduced these changes in liver (Fig. 7A). The plasma ALT and aspartate aminotransferase (AST) levels were elevated by binge alcohol, but administration of PNVs decreased plasma liver toxicity marker (Fig. 7B, C). The hepatic TG level was significantly increased in binge alcohol, but the administration of PNVs significantly decreased these levels (Fig. 7D).
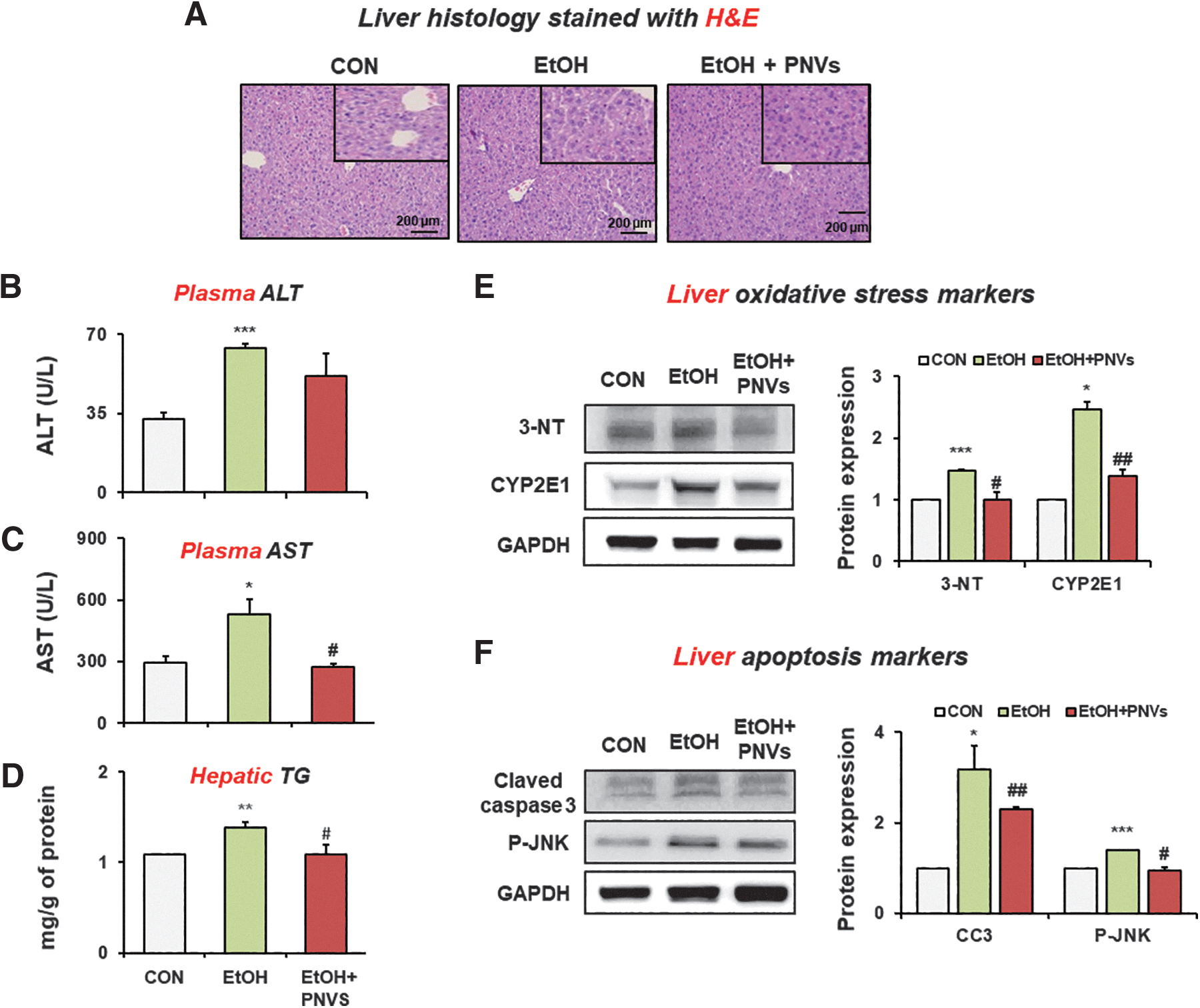
PNVs attenuated binge alcohol-induced acute liver injury in mice.
In addition, oxidative stress markers such as CYP2E1 and 3-NT were increased in the EtOH-exposed mouse group, but were significantly decreased by the oral administration of PNVs (Fig. 7E). The hepatic apoptosis markers such as cleaved-caspase 3 and phosphorylated (p)-JNK were increased in the EtOH-exposed mouse group, but were reduced by the oral administration of PNVs (Fig. 7F). These results confirm that PNVs can prevent liver damage by inhibiting oxidative stress and apoptosis marker protein expression.
PNVs protected the oxidative stress and apoptosis marker proteins caused by alcohol in AML 12 liver cells
To confirm the in vivo results, we tested PNV treatment effect on AML12 normal liver cells by 100 mM ethanol. Our results showed that the elevation of oxidative stress and apoptosis markers protein by alcohol treatment was reduced in PNV-treated AML12 cells (Fig. 8A, B). The Bax/Bcl-2 ratio was significantly increased in the PNV-treated AML12 cells (Fig. 8B). Furthermore, PNVs significantly suppressed the elevated mRNA levels of iNOS, COX-2, and TLR-4 in alcohol-exposed AML12 cells (Fig. 8C–E). Fluorescence image analysis showed higher CYP2E1 protein expression in alcohol-exposed AML12 liver cells. Treatment of PNVs reduced the CYP2E1 protein expression (Fig. 8F). Therefore, PNVs can prevent alcohol-induced hepatic oxidative stress and inflammation in vitro.
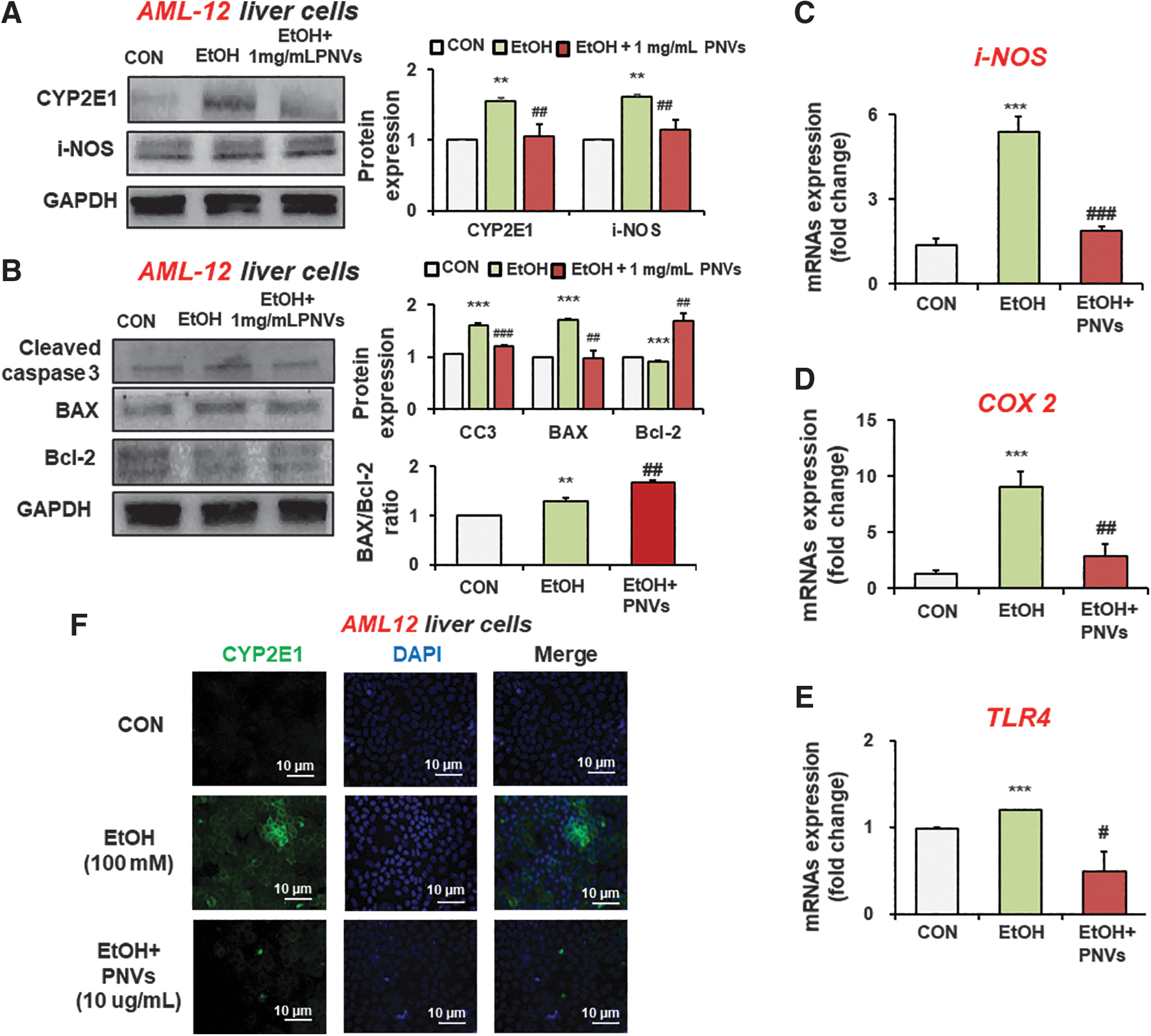
PNVs alleviated hepatocyte damage in alcohol-exposed AML-12 liver cells.
Discussion
For further application of edible plant-derived exosome-like nanovesicles from food industry, PENVs have numerous benefits of nano size, biocompatibility, stability, low toxicity, and large production. PENVs have recently been applied in a variety of ways to treat inflammatory diseases, cancer, and ALDs. Pomegranate reduced the intestinal oxidative stress, nitration, acetylation, and ubiquitination in binge alcohol-exposed rats. 8 In previous studies, it was known that pomegranates contain organic acid, EA, sucrose, ascorbic acid, citric acid, and malic acid, and bioactive compounds such as phenolics and flavonoids, and principally anthocyanins. 11,13 It is known to be effective against cancer, cardiovascular disease, diabetes, ALD, and women's diseases. 31 –33 In this study, pomegranate juice isolated the exosome-like nanovesicles and characterized the nano size and lipid layer. Our results determined that PNVs prevented alcohol-induced liver and gut damage for the first time (Fig. 1).
In this study, PNVs were successfully isolated nano-size vesicles using our laboratory method. The morphological characterization of PNVs was confirmed by TEM analysis. NTA was performed to determine the size and distribution of PNVs. Our data determined that PNVs showed nano size and lipid bilayer by NTA and TEM, as recently reported. To evaluate biological properties, the biodistribution and toxicity of PNVs were first tested by edible PNV oral administration. In addition, our data show that PNVs were no toxicity in liver, kidney, brain, heart, spleen, and lung by H&E staining. Recently, pomegranate extracellular vesicles (EVs) showed the noncytotoxicity of Caco-2 or THP-1 cells. 13 These results are consistent with several reports that EVs from edible plant are nontoxic.
In this study, we investigated whether PNVs prevent binge alcohol-induced leaky gut and liver damage. It is well established that chronic or excessive amounts of alcohol intake can induce damage trough gut-liver axis. 34 Several research have shown that alcohol destroys the intestinal epithelial barrier, increases intestinal permeability, and accelerates liver damage by activating TLR-4, by increasing plasma endotoxin concentration. 4,5 One of the main damage by binge alcohol exposure is involved in alcohol-induced gut leaky through modulating the oxidative stress marker protein expression.
Previous studies have shown that PNVs prevented the junctional proteins such as ZO-1, claudin-4, occludin, E-cadherin, β-catenin, and α-tubulin and inhibited oxidative stress and apoptosis marker proteins by binge alcohol exposed leaky gut. 8 In this study, our results show that PNVs markedly reduced the plasma endotoxin and elevated the reduction of junctional proteins by binge alcohol-induced gut permeability. We confirmed that PNVs increased the reduction of junctional protein expression in T84 colonic cells treated with alcohol. PNVs prevented the reduced amounts of oxidative stress and apoptosis marker proteins by binge alcohol-exposed leaky gut. Therefore, these findings indicate that PNVs have a significant protective effect on binge alcohol-induced leaky gut.
In this study, PNVs reduced the plasma ALT and AST as an indicator of liver damage caused by alcohol. Also, hepatic TG and fat accumulation in the liver by H&E showed elevation of binge alcohol; however, PNVs reduced these levels. PNVs prevented the increased amounts of oxidative stress such as iNOS, CYP2E1, and 3-NT by binge alcohol. Also, we confirmed these results in AML12 liver normal cells. These results found that PNVs significantly showed hepatoprotective effects against binge alcohol-induced liver damage.
In summary, exosome-like nanovesicles from edible pomegranate juice were successfully isolated, purified, and characterized using optimized methods. PNVs showed nontoxicity and large production form pomegranate juice. Markedly, PNVs demonstrated a protective mechanism against leaky gut and liver damage caused by binge alcohol. Taken together, PNVs can be used as a potentially hepatoprotective or intestinal protective agent by binge alcohol-induced gut and liver damage for the first time.
Footnotes
Authors' Contributions
J.S.K., and Y.E.C. designed the study. J.S.K. and D.H.K. performed the experiments, analyzed data, made the figures, and performed the statistical analysis. M.C.G., W.H.S., D.K.K., and Y.E.C. drafted the initial version of this article, and then all authors contributed to the editing of the article. All authors have read and approved the final version of this article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by National Research Foundation of Korea (NRF) grant funded by Korea government (MSIT) (No. 2021R1C1C1008117, 2021R1A4A1033078, 2018R1A5A2025286, and 2022R1C1C1008912). This work was also supported by the Ewha Womans University Research Grant (2021).