
Abstract
This study investigated the protective effects of a complex of Indian gooseberry and barley sprout (IB complex) on oxidative stress and skin damage caused by ultraviolet B irradiation in SHK-I hairless mice. The study examined the impact of IB complex on skin hydration, wrinkle formation, and melanogenesis using enzyme-linked immunosorbent assay, real-time polymerase chain reaction, and western blot analysis. The IB complex reduced skin hydration loss and wrinkle formation, while also demonstrating enhanced antioxidant activities. The IB complex maintained skin hydration via upregulation of hyaluronic acid and ceramide synthesis, including the regulation of hyaluronic acid synthase, long-chain ceramide formation, dihydroceramide desaturase 1 activity, and type I collagen production. The IB complex prevented wrinkle formation via downregulating JNK and upregulating TGF-β pathways. Moreover, IB complex blocked melanin production via inhibition of protein kinase A, cAMP response element-binding protein, and microphthalmia-associated transcription factor pathways. These results suggest that IB complex is a potential agent to protect the skin against photodamage caused by exposure to UVB radiation. The research protocols underwent approval from the Institutional Animal Care and Use Committee of Kyung Hee University (KHGASP-21-577), ensuring compliance with ethical standards.
INTRODUCTION
The skin is the largest organ in the human body and serves as the first line of defense against external environmental factors. It is a complex structure composed of several layers, each with its own unique features and functions. The epidermis, as the outermost layer, is responsible for protecting the body from ultraviolet (UV) radiation, pathogens, and other harmful substances. 1,2
The impact of UVB irradiation on the skin has been extensively studied in recent years, as excessive exposure to this type of radiation causes a wide range of negative effects. Among the most significant effects of UVB radiation on the skin are dryness, wrinkle formation, and changes in melanin production. UVB radiation induces oxidative stress in the skin, leading to the production of reactive oxygen species (ROS) and reactive nitrogen species (RNS). These free radicals cause damage to cellular components, including lipids, proteins, and DNA. This damage disrupts the skin's natural barrier function and leads to increased water loss, resulting in dryness. 3 –5 Furthermore, UVB radiation stimulates the expression of matrix metalloproteinases (MMPs), enzymes that degrade the extracellular matrix (ECM) in the skin.
This degradation leads to a loss of collagen and elastin fibers, which are essential for maintaining skin elasticity and firmness. The resulting structural changes cause wrinkles and other signs of aging. 6 –8 Melanin production is also regulated by molecular mechanisms that are impacted by UVB irradiation. UVB radiation activates the enzyme tyrosinase, which engages in the production of melanin. This intensification in melanin production is a natural protective response of the skin to UV radiation, as melanin helps to absorb and scatter UVB photons. However, excessive melanin production can lead to hyperpigmentation, dark spots, and other forms of skin damage. 9,10
Indian gooseberry (Emblica officinalis), also called amla, has bioactive components for antioxidant, immunomodulatory, gastroprotective, anticancer, antiobesity, and antidiabetic effects. Recently, the consumption of Indian gooseberry extracts has improved in food, pharmaceutical, and cosmetic products to health. 11 –14 Barley sprout (Hordeum vulgare L.) has various flavonoids that are effective in obesity, alcoholic fatty liver, diabetic, inflammation, and cardiovascular diseases. 15 –17 Extracts of Indian gooseberry and barley sprout protect skin damage by UVB irradiation, but research on their mixture is currently insufficient. However, we have previously determined the antiobesity effects of Indian gooseberry and barley sprout complex using cell and animal models. 18 In this study, we investigated whether Indian gooseberry and barley sprout complex might protect the skin against UVB exposure to SHK-I hairless mice.
MATERIALS AND METHODS
Indian gooseberry and barley sprout complex preparation
Indian gooseberry extract powder (I) and barley sprout juice powder (B) were provided from HL Science Co., Ltd. (Uiwang, Korea) and the IB complex was composed of I:B = 2:1. The extraction methods of Indian gooseberry and barley sprout were described previous study. 18 The powders were kept at −20°C until the experiments were performed. The active compound of Indian gooseberry and barely sprout was detected by high-performance liquid chromatography and data were described in the previous study. 19
Experimental animals and treatment
The experiments were permitted by the Institutional Animal Care and Use Committee of Kyung Hee University (KHGASP-21-577). Fifty-six SKH-1 hairless mice (male, 5-week-old) were purchased from SaeRon Bio (Uiwang, Korea). The mice were acclimated in an automatically managed condition room (12/12 h light/dark cycle, 45 to 55% relative humidity, and at 23°C ± 2°C). The mice were separated into seven groups of eight animals each as follows: AIN93G without UVB exposure (normal control, NC), AIN93G with UVB exposure (control, C), AIN93G supplemented 100 mg/kg body weight (BW) of L-ascorbic acid (L-AA) with UVB exposure, AIN93G supplemented 100 mg/kg BW of arbutin with UVB exposure (arbutin), AIN93G supplemented 100 mg/kg BW of IB complex with UVB exposure (IB complex 100), AIN93G supplemented 200 mg/kg BW of IB complex with UVB exposure (IB complex 200), and AIN93G supplemented 400 mg/kg BW of IB complex with UVB exposure (IB complex 400).
UVB irradiation
All groups, except NC, were exposed to UVB three times per week using a UVB lamp (Sankyo Denki Co., Yokohama, Japan). The UVB intensity started at 150 mJ/cm2 (first week) and progressively increased to 300 mJ/cm2 (second week), 450 mJ/cm2 (third week), and 600 mJ/cm2 (4–8 weeks).
Measurement of hydration, transepidermal water loss, and hyaluronic acid of skin and histological observations
The hydration, transepidermal water loss (TEWL), hyaluronic acid (HA), thickness, and wrinkle formation of the dorsal skin were measured using the methods in the previous study. 20,21
Measurement of antioxidant enzyme activities, tyrosinase, nitric oxide, and cyclic adenosine monophosphate
The activities of SOD (superoxide dismutase 1 ELISA kit; BioVision, Inc., Milpitas, CA, USA), CAT (Catalase Activity Colorimetric/Fluorometric Assay Kit; BioVision, Inc.), GPx (Glutathione Peroxidase Activity Colorimetric Assay Kit; BioVision, Inc.), and tyrosinase (Tyrosinase Activity Assay Kit; Abcam, Cambridge, UK), and the levels of NO (Nitric Oxide Assay Kit; Abcam) and cyclic adenosine monophosphate (cAMP ELISA kit; Enzo Life Sciences, PA, USA) were measured according to manufacturer's manuals.
Isolation of total RNA and real-time polymerase chain reaction
Isolation of total RNA from dorsal skin tissue and real-time polymerase chain reaction (PCR) analysis for expression of the genes of hyaluronic acid synthase (HAS), long-chain ceramides (LCB1), dihydroceramide desaturase 1 (DEGS1), fibrillin-1, pro-inflammatory cytokines (tumor necrosis factor-alpha [TNF-α], interleukin [IL]-1β, IL-6), transforming growth factor beta receptor 1 (TGFβR1), procollagen C-endopeptidase enhancer (PCOLCE) protein, collagen type 1 alpha 1 (COL1A1), and glyceraldehyde 3-phosphate dehydrogenase were performed according to previous methods. 20,21 The primer sequences were described Table 1.
Primer Sets Used for Real-Time Polymerase Chain Reaction
M, mouse; HAS, hyaluronic acid synthase; LCB1 (SPT), long-chain base biosynthesis protein 1 (serine palmitoyltransferase); DEGS1, delta 4-desaturase sphingolipid 1; COL1A, collagen type 1 alpha; IL, interleukin; TGF-β RI, transforming growth factor beta receptor 1; PCOLCE, procollagen C-endopeptidase enhancer; Pro-COL1, procollagen type 1; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; TNF-α, tumor necrosis factor-alpha.
Isolation of protein and western blot analysis
Extraction of protein from dorsal skin tissue and western blot analysis for the protein expression of CerS4 (LASS4), IκBα, phospho-IκBα, p65, phospho-p65, COL1A1, c-Jun N-terminal kinase (JNK), phospho-JNK (p-JNK), c-Fos, phospho-c-Fos, c-Jun, phospho-c-Jun, MMP-1, MMP-2, MMP-3, and MMP-9, cyclooxygenase-2 (COX-2), Smad3, phos-pho-Smad3, protein kinase A (PKA), phospho-PKA, cAMP response element-binding protein (CREB), phospho-CREB, microphthalmia-associated transcription factor (MITF), tyrosinase-related protein (TRP)-1, TRP-2, and β-actin were conducted according to previous methods. 20,21
Statistical analysis
All results are illustrated as mean ± standard deviation. The data were statistically determined using Duncan's multiple range tests after one-way analysis of variance using SPSS (SPSS PASW Statistic v.23.0; SPSS, Inc., Chicago, IL, USA). Differences were considered statistically significant at P < 0.05.
RESULTS
Effects of IB complex on skin hydration, TWEL, HA, and wrinkle formation in UVB-exposed SKH-I hairless mice
We measured the effect of IB complex on BW gain, food intake, food efficiency rate, and organ (liver, kidney, spleen) weights of UVB-irradiated SHK-I hairless mice. There were no significant differences among groups, which means that IB complex supplementation did not affect the experimental results (Table 2).
Body Weight Gain, Food Consumption, Food Efficiency Rate, and Organ Weight
Values are presented as means ± SD (n = 8). Different letters show a significant difference by ANOVA with Duncan's multiple range test at p < 0.05.
NC: AIN93G, C: ultraviolet B (UVB) irradiation+AIN93G, PC1: UVB irradiation+AIN93G+L-ascorbic acid 100 mg/kg b.w., PC2: UVB irradiation+AIN93G+Arbutin 100 mg/kg b.w., IB complex 100: UVB irradiation+AIN93G+IB complex 100 mg/kg b.w., IB complex 200: UVB irradiation+AIN93G+IB complex 200 mg/kg b.w., IB complex 400: UVB irradiation+AIN93G+IB complex 400 mg/kg b.w.
Weight gain (g/8 weeks) = final BW (g) − initial BW (g).
FER = {weight (g)/food intake (g)} * 100.
ANOVA, analysis of variance; BW, body weight; FER, food efficiency rate; L-AA, L-ascorbic acid; NC, normal control; SD, standard deviation.
To investigate the effect of IB complex on the morphology and histopathology of the dorsal skin of UVB-irradiated SHK-I hairless mice, we measured the skin hydration, TWEL, HA, wrinkle, and thickness of the dorsal skin. The skin hydration was significantly lower in the control group compared to that in the NC group, and significantly enhanced in the L-AA, arbutin, and IB complex 400 (40.7%, 23.8%, and 19.9%, respectively) (P < .05) (Fig. 1A). TEWL was significantly augmented in the control group compared to that in the NC group, and significantly diminished in the L-AA, arbutin, and IB complex 400 (52.9%, 28.2%, and 23.5%, respectively (P < .05) (Fig. 1B).

Effects of IB complex on skin hydration
HA was markedly declined in the control group compared to that in the NC group, and meaningfully increased in the L-AA, arbutin, IB complex 200, and IB complex 400 (170.7%, 89.3%, 43.8%, and 99.3%, respectively) (P < .05) (Fig. 1C). Wrinkle area, epidermal thickness, and dermis thickness were significantly enlarged in the control group compared to that in the NC group and drastically decreased in the L-AA and arbutin groups compared to that in the control group (P < .05) (Fig. 1E–G). In addition, the wrinkle area, epidermal thickness, and dermal thickness were significantly decreased in the IB complex compared to that in the control group (11.1%–45.4%, 20.2%–38.8%, and 17.7%–26.6%, respectively) (P < .05) (Fig. 1E–G). L-AA, arbutin, and IB complex supplement groups improved the morphological and histopathological changes induced by UVB irradiation, including skin hydration, wrinkle formation, and epidermal and dermis thickness.
Effects of IB complex on antioxidant activities and mRNA expression of proinflammatory cytokines (TNF-α, IL-1β, IL-6) in UVB-exposed SKH-I hairless mice
To investigate the effect of IB complex on the antioxidant activities and proinflammatory cytokines of the dorsal skin of UVB-irradiated SHK-I hairless mice, we analyzed the dorsal skin from UVB-irradiated SHK-I hairless mice using ELISA kits and real-time PCR. The antioxidant enzyme activities (SOD, CAT, and GPx) were significantly decreased in the control group compared to the NC group, and substantially enhanced in the L-AA and arbutin groups compared to that in the control group (P < .05) (Fig. 2A–C). The SOD and CAT activities were significantly increased in the IB complex 200 and IB complex 400 compared to that in the control group (16.7% and 45.7%, 51.8%, and 127.6%, respectively) (P < .05) (Fig. 2A, B) and the GPx activities were notably amplified in the IB complex compared to the control group (20.9%–32.1%) (P < .05) (Fig. 2C).

Effects of IB complex on antioxidant activities of SOD
Besides, the mRNA expression of TNF-α, IL-1β, and IL-6 was significantly increased in the control group compared to that in the NC group, and significantly decreased in the L-AA and arbutin groups compared to that in the control group (P < .05) (Fig. 2D–F). The mRNA expression of TNF-α was significantly reduced in the IB complex compared to that in the control group (12.3%–37.2%) (P < .05) (Fig. 2D). The mRNA expression of IL-1β was markedly declined in the IB complex 200 and IB complex 400 compared to that in the control group (26.5% and 36.3%, respectively) (P < .05) (Fig. 2E). The mRNA expression of IL-6 was significantly diminished in the IB complex-treated groups compared to that in the control group (12.7%–41.7%) (P < .05) (Fig. 2F).
These results suggest that L-AA, arbutin, and IB complex supplement groups ameliorated the antioxidant enzyme activities and mRNA expression of proinflammatory cytokines induced by UVB irradiation.
Effects of IB complex on mRNA expression of skin moisturizing-related factors and collagen synthesis-related factors in UVB-exposed SKH-I hairless mice
To investigate the changes in mRNA expression of skin moisturizing-related factors and collagen synthesis-related factors by IB complex supplementation, we analyzed the dorsal skin from UVB-irradiated SKH-I hairless mice using real-time PCR. The mRNA expression of skin moisturizing-related factors such as HAS1-3, fibrillin-1, LCB1 (SPT), and DEGS1 was significantly decreased in the control group compared to that in the NC group, and significantly increased in the L-AA and arbutin groups compared to that in the control group (P < .05) (Fig. 3A–F). The mRNA expression of HAS1, HAS2, and HAS3 was markedly improved in the IB complex 200 and IB complex 400 compared to that in the control group (56.9%, and 117.8%, 31.2% and 85.7%, 39.7% and 93.6%, respectively) (P < .05) (Fig. 3A–C), and the mRNA expression of fibrillin-1 was significantly notably increased in the IB complex-treated groups compared to that in the control group (64.6%–173.4%) (P < .05) (Fig. 3D).
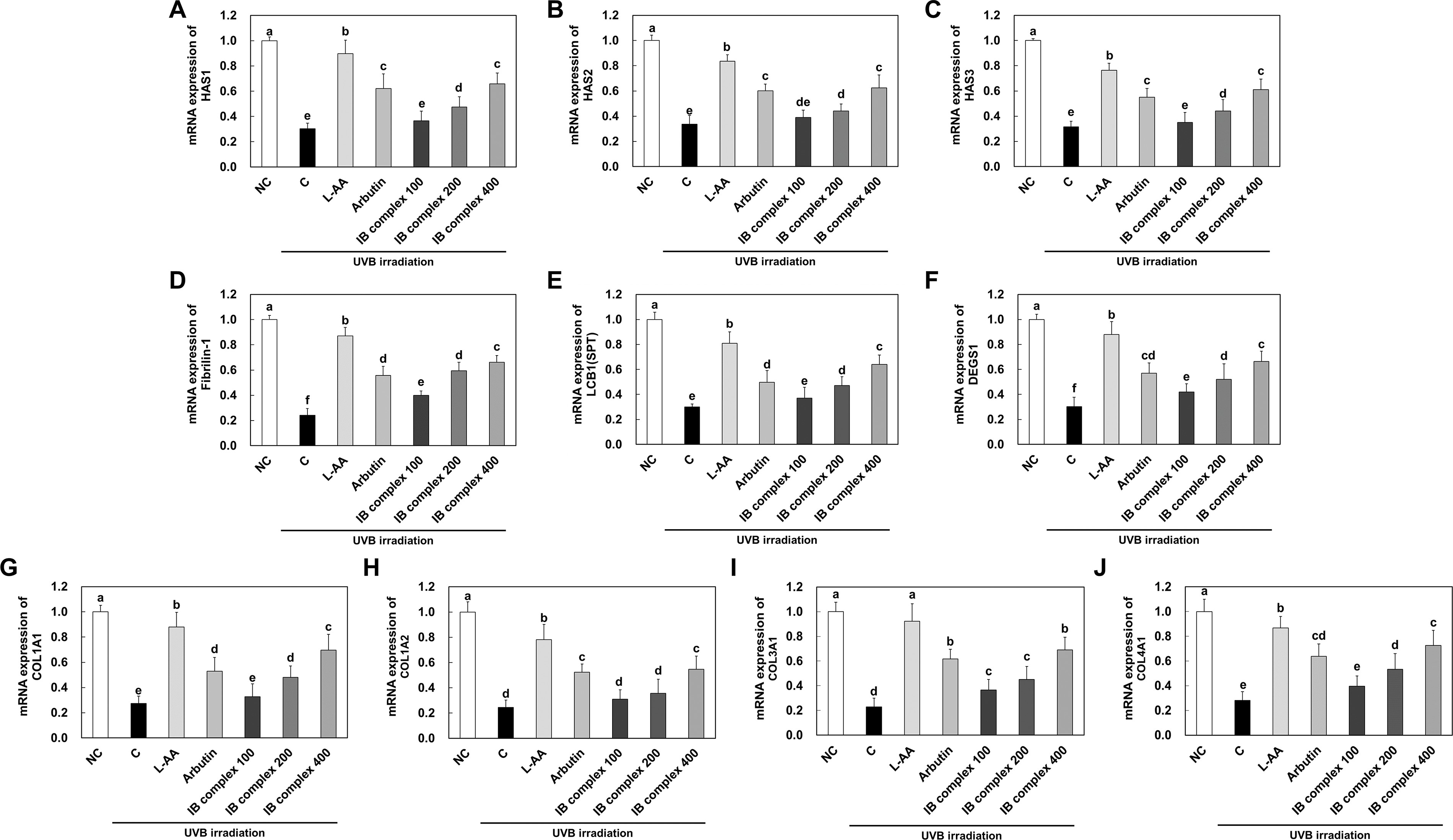
Effects of IB complex on mRNA expressions of HAS1
The mRNA expression of LCB1 (SPT) was drastically increased in the IB complex 200 and IB complex 400 compared to that in the control group (57.3% and 114.5%, respectively) (P < .05) (Fig. 3E). In addition, the mRNA expression of DEGS1 was significantly enhanced in the IB complex compared to that in the control group (39.1%–120.4%) (P < .05) (Fig. 3F). The mRNA expression of collagen synthesis related factors such as COL1A1, COL1A2, COL3A1, and COL4A1 was appreciably declined in the control group compared to that in the NC group, and significantly increased in the L-AA and arbutin groups compared to that in the control group (P < .05) (Fig. 3G–J). The mRNA expression of COL1A1 was significantly increased in the IB complex 200 and IB complex 400 compared to that in the control group (74.8% and 153.3%, respectively) (P < .05) (Fig. 3G).
The mRNA expression of COL1A2 was significantly augmented in the IB complex 400 compared to that in the control group (122.4%), whereas the IB complex 100 and IB complex 200 were not significantly different (26.4% and 44.8%, respectively) (P < .05) (Fig. 3H). The mRNA expression of COL3A1 was significantly increased in the IB complex compared to that in the control group (61.0%–203.7%) (P < .05) (Fig. 3I). In addition, the mRNA expression of COL4A1 was significantly increased in the IB complex 200 and IB complex 400 compared to that in the control group (89.7% and 157.9%, respectively) (P < .05) (Fig. 3J). These results demonstrated that L-AA, arbutin, and IB complex supplement groups improved the mRNA expression of skin moisturizing-related factors and collagen synthesis-related factors induced by UVB irradiation.
Effects of IB complex on protein expression of skin moisturizing-related factors in UVB-exposed SKH-I hairless mice
To investigate the changes in protein expression of skin moisturizing-related factors by IB complex supplementation, we analyzed the dorsal skin from UVB-irradiated SKH-I hairless mice using western blot analysis. The protein expression of CerS4 was significantly decreased in the control group compared to that in the NC group, whereas L-AA, arbutin, and IB complex supplement groups were significantly greater compared to that in the control group (123.0%, 86.5%, and 66.4% to 78.9%, respectively) (P < .05) (Fig. 4B). The protein expression of p-IκB/IκB, p-p65/p65, and COX-2 was significantly heightened in the control group compared to that in the NC group, and markedly decreased in the L-AA and arbutin groups compared to that in the control group (P < .05) (Fig. 4C–E).

Effects of IB complex on protein bands
The protein expression of p-IκB/IκB was significantly diminished in the IB complex compared to that in the control group (40.7%–61.0%) (P < .05) (Fig. 4C), and the protein expression of p-p65/p65 was significantly decreased in the IB complex compared to that in the control group (50.3%–56.8%) (P < .05) (Fig. 4D). In addition, the protein expression of COX-2 was significantly decreased in the IB complex compared to that in the control group (25.9%–63.9%) (P < .05) (Fig. 4E). These results suggest that L-AA, arbutin, and IB complex supplement groups improved the protein expression of skin moisturizing-related factors induced by UVB irradiation.
Effects of IB complex on mRNA and protein expression of skin wrinkle formation-related factors in UVB-exposed SKH-I hairless mice
To investigate the changes in mRNA and protein expression of skin wrinkle formation-related factors by IB complex supplementation, we analyzed the dorsal skin from UVB-irradiated SKH-I hairless mice using real-time PCR and western blot analysis. The mRNA expression of TGFβR1, PCOLCE, and pro-COL1 was meaningfully decreased in the control group compared to that in the NC group, and significantly enhanced in the L-AA and arbutin groups compared to that in the control group (P < .05) (Fig. 5A–C). The mRNA expression of TGFβR1 was notably increased in IB complex 200 and IB complex 400 compared to that in the control group (99.5% and 126.4%, respectively) (P < .05) (Fig. 5A), and mRNA expression of PCOLCE was significantly increased in IB complex compared to that in the control group (44.3%–138.0%) (P < .05) (Fig. 5B).
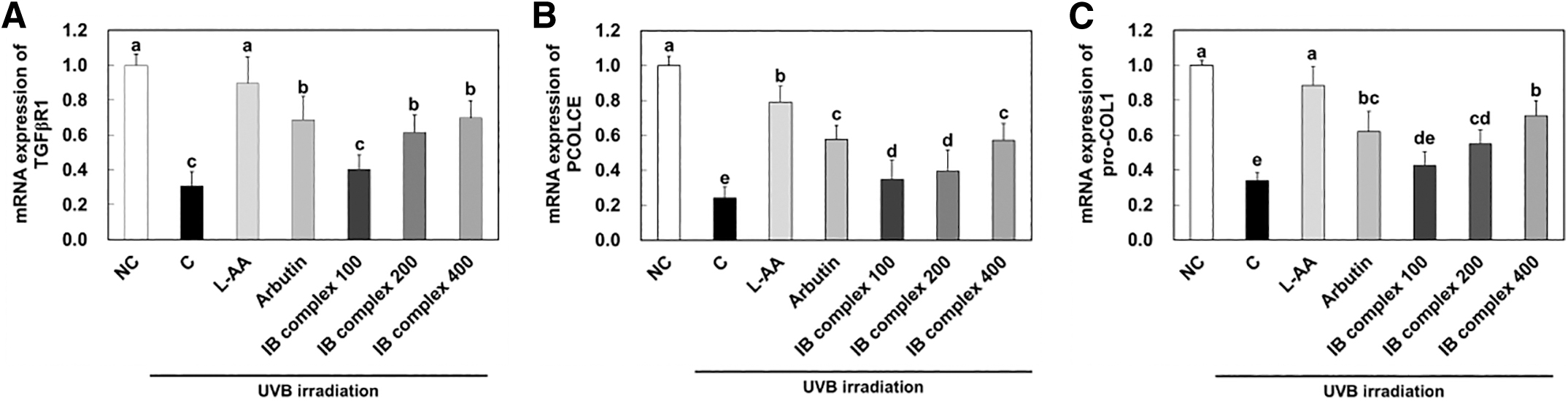
Effects of IB complex on mRNA expression of TGFβR1
Also, the mRNA expression of pro-COL1 was significantly amplified in IB complex 200 and IB complex 400 compared to that in the control group (62.5% and 110.0%, respectively) (P < .05) (Fig. 5C). The protein expression of COL1 was significantly decreased in the control group compared to that in the NC group, and significantly increased in the L-AA and arbutin groups compared to that of the control group (P < .05) (Fig. 6B). The protein expression of p-JNK/JNK, p-c-Fos/c-Fos, p-c-Jun/c-Jun, MMP-1, MMP-2, MMP-3, and MMP-9 was extensively increased in the control group compared to the NC group, whereas L-AA and arbutin groups were significantly decreased compared to the control group (P < .05) (Fig. 6C–I).
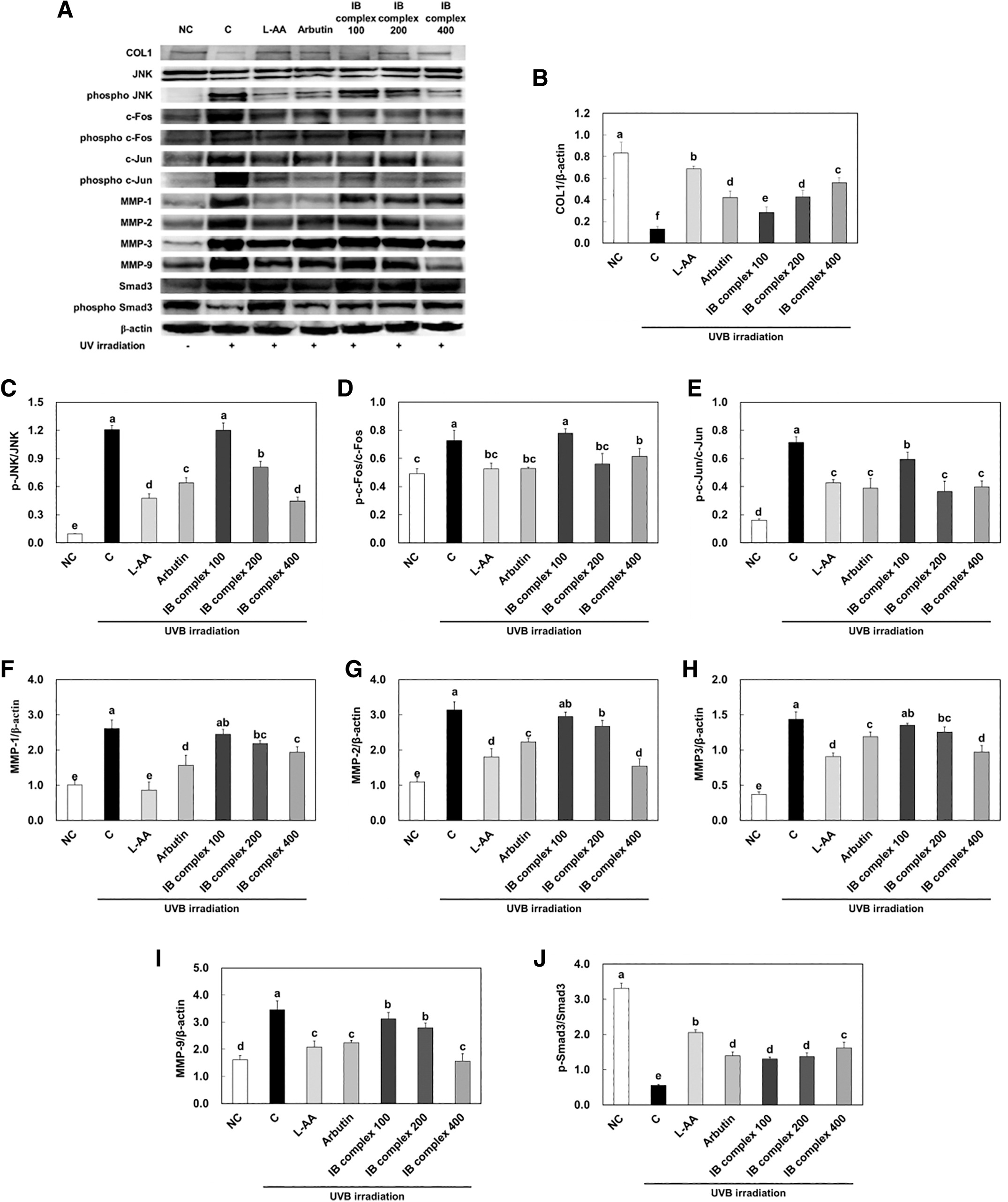
Effects of IB complex on protein bands
Also, the protein expression of p-Smad3/Smad3 was significantly declined in the control group compared to that in the NC group, and significantly enhanced in the L-AA and arbutin groups compared to that of the control group (P < .05) (Fig. 6J). The protein expression of COL1 was significantly increased in IB complex compared to that in the control group (117.2%, 228.0%, and 329.3%) (P < .05) (Fig. 6B). However, the protein expression of p-JNK/JNK and p-c-Fos/c-Fos were significantly decreased in IB complex 200 and IB complex 400 compared to that in the control group (33.2% and 62.9%, 22.9%, and 15.4%, respectively), and protein expression of p-c-Jun/c-Jun was significantly decreased in IB complex compared to that in the control group (17.0%–49.0%) (P < .05) (Fig. 6C–E).
The protein expression of MMP-1 was significantly decreased in the IB complex 200 and IB complex 400 compared to that in control group (16.1% and 25.9%, respectively) (P < .05) (Fig. 6F), and the protein expression of MMP-2 was significantly decreased in the IB complex 200 and IB complex 400 compared to that in control group (14.7% and 50.9%, respectively) (P < .05) (Fig. 6G). Also, the protein expression of MMP-3 was significantly diminished in IB complex 200 and IB complex 400 compared to that in the control group (12.7% and 32.2%, respectively) (P < .05) (Fig. 6H), and the protein expression of MMP-9 was significantly declined in the IB complex compared to that in the control group (9.9%–54.8%) (P < .05) (Fig. 6I). The protein expression of p-Smad3/Smad3 was significantly increased in IB complex compared to that in the control group (134.4%–191.3%) (Fig. 6J).
These results suggest that L-AA, arbutin, and IB complex supplement groups preserved the protein expression of skin wrinkle formation-related factors induced by UVB irradiation.
Effects of IB complex on melanogenesis-related factors in UVB-exposed SKH-I hairless mice
To investigate the changes in protein expression of melanogenesis-related factors by IB complex supplementation, we analyzed the dorsal skin from UVB-irradiated SKH-1 hairless mice using an ELISA kit and western blot analysis. The tyrosinase activity, nitric oxide (NO), and cAMP concentrations were significantly increased in the control group compared to that in the NC group, whereas the L-AA and arbutin groups were significantly declined compared to the control group (P < .05) (Fig. 7A–C). The tyrosinase activity was significantly decreased in IB complex 200 and IB complex 400 compared to that in the control group (21.7% and 30.4%, respectively) (P < .05) (Fig. 7A). The NO concentrations were significantly diminished in IB complex 400 compared to that in the control group (28.9%) (P < .05) (Fig. 7B).
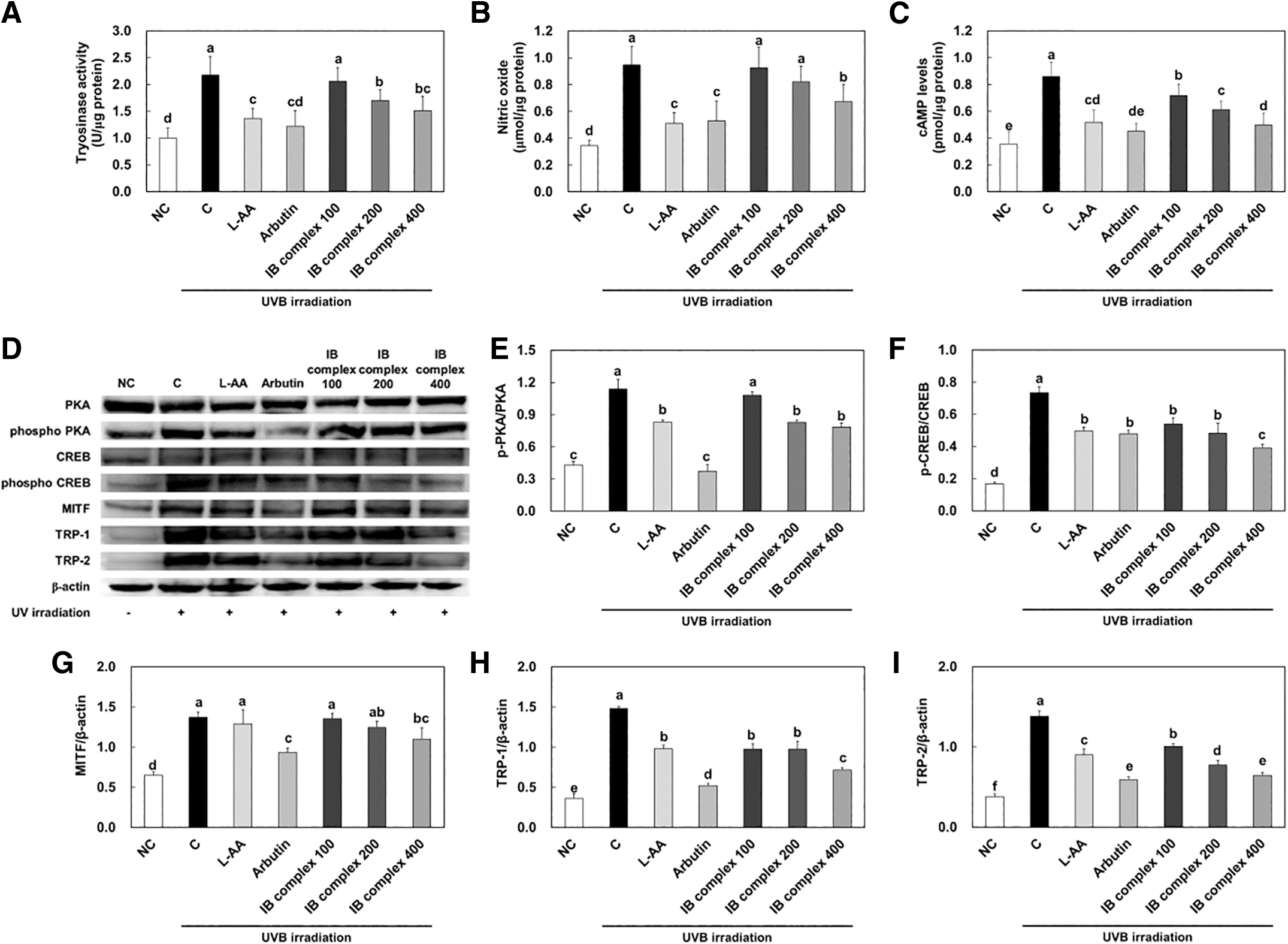
Effects of IB complex on tyrosinase activity
The cAMP levels were significantly decreased in IB complex 200 and IB complex 400 compared to that in the control group (28.7% and 42.2%, respectively) (P < .05) (Fig. 7C). The protein expression of p-PKA/PKA, p-CREB/CREB, MITF, TRP-1, and TRP-2 was significantly increased in the control group compared to that in the NC group, and significantly decreased in the L-AA and arbutin groups compared to that in the control group (P < .05) (Fig. 7E–I). The protein expression of p-PKA/PKA was significantly decreased in IB complex 200 and IB complex 400 compared to that in the control group (27.5% and 31.2%, respectively) (P < .05) (Fig. 7E). The protein expression of p-CREB/CREB, MITF, TRP-1, and TRP-2 were significantly reduced in IB complex compared to that in the control group (26.6%–46.8%, 1.2%–20.2%, 34.1%–51.7%, and 27.2%–53.3%) (P < .05) (Fig. 7F–I).
These results demonstrate that L-AA, arbutin, and IB complex supplement groups protected the protein expression of skin melanogenesis-related factors induced by UVB irradiation.
DISCUSSION
The skin is composed of three layers which consist of the epidermis, dermis, and the hypodermis. The skin has various functions such as maintaining body temperature, preventing water loss, and protecting the body from chemicals, pathogens, harmful substances, and UV radiation. UV radiation has been divided into different types in accordance with wavelength differences as follows: UVA (320–400 nm), UVB (280-320 nm), and UVC (200–280 nm). 22 –24 Antioxidant enzymes (SOD, CAT, and GPx) play a vital role in protecting the skin from photodamage caused by exposure to UV. Exposure to UV activates ROS and other free radicals, which leads to damage to the skin, including sunburn, premature aging, and skin cancer. 25 This study investigated whether IB complex could protect against the skin damage caused by UVB-irradiation in SHK-I mice via effects on skin hydration, wrinkle formation, and melanogenesis.
Loss of skin hydration and wrinkle formation were reduced in IB complex supplemented groups, and the antioxidant activities were enhanced in IB complex supplemented groups. Also, L-AA and arbutin were used as positive controls. L-AA was reported that which could help protect the skin from the harmful effects of free radicals and is essential for the production of collagen. Arbutin is a tyrosinase inhibitor, meaning it could help reduce the melanin production. 26,27
Skin hydration is a critical process that involves the regulation of multiple molecular mechanisms, including the regulation of HAS, the formation of LCBs by LCB1 (SPT), the activity of DEGS1, the production of COL1As, and the regulation of nuclear factor-kappa B (NF-κB), MMPs, and COX-2. 28,29 HA is a key component of the skin's ECM, where it functions to bind and retain water molecules. The synthesis of HA is regulated by three isoforms of HAS, which catalyze the production of HA from precursor molecules. LCBs are a type of sphingolipid that is critical for the formation of the skin's barrier function. LCB1 is an enzyme that participates in the synthesis of LCBs, which are subsequently converted into ceramides. Ceramides play a critical role in maintaining the skin's barrier function by forming a protective layer that helps to prevent water loss. 30 –32
DEGS1 is an enzyme that is important in the synthesis of sphingolipids and catalyzes the conversion of dihydroceramides to ceramides. 33 The COL1A genes, particularly COL1A1, play a crucial role in the synthesis of type I collagen, the most abundant collagen found in the skin. Type I collagen is a key structural component that provides strength, elasticity, and support to the skin. 34,35 In addition, NF-κB and COX-2 activation can result in down-regulation of filaggrin, a protein important for skin barrier integrity, which may contribute to skin inflammation and barrier dysfunction. 36 –38 In this study, IB complex increased mRNA expression of HAS1-3, LCB1 (SPT), DEGS1, and COL1A1, COL1A2, COL3A1, COL4A1, and protein expression of CerS4. Besides, IB complex decreased protein expression of p-IκB/IκB and COX-2. These results suggest that IB complex may control skin hydration via the upregulation of HA and ceramide synthesis and downregulation of NF-κB pathways.
The formation of skin wrinkles is a complex process involving multiple molecular mechanisms. One of the key pathways involved in wrinkle formation is the activation of the JNK pathway. JNK is a stress-activated protein kinase that is activated in response to various stress stimuli, including UV radiation. JNK activation leads to the phosphorylation and activation of c-Jun and other transcription factors, including c-Fos. These transcription factors activate the expression of genes involved in ECM remodeling, including MMPs. MMPs are upregulated in response to various stimuli, including UV radiation, and contribute to wrinkle formation. In addition to the JNK pathway, another key pathway involved in wrinkle formation is the transforming growth factor beta (TGF-β) pathway. TGF-β signaling leads to the activation of Smad3, which plays a key role in ECM synthesis and degradation.
Smad3 downregulates the expression of MMPs, leading to inhibited ECM degradation and wrinkle formation. 39,40 In this study, IB complex decreased protein expression of p-JNK/JNK, p-c-Fos/c-Fos, p-c-Jun/c-Jun, and MMPs, whereas IB complex increased protein expression of p-Smad3/Smad3. These results suggest that IB complex could protect wrinkle formation via the downregulation of JNK, c-Fos, c-Jun, and MMPs pathways and the upregulation of Smad3.
Melanin is a skin pigment, which provides a first line of defense against UV radiation. Melanogenesis is regulated by multiple signaling pathways, including the PKA and the CREB pathways. PKA is a protein kinase that plays a key role in melanogenesis. When activated, PKA phosphorylates and activates the transcription factor CREB. Activated CREB then binds to the promoter region of the MITF gene and increases its expression. MITF is a transcription factor that regulates the expression of several genes involved in melanin synthesis, including tyrosinase, TRP-1, and TRP-2. Tyrosinase catalyzes the conversion of tyrosine to DOPA, which is subsequently converted to melanin. NO is a signaling molecule that modulates tyrosinase activity in melanocytes, the cells that produce melanin. Nitric oxide plays a significant role in melanogenesis by up-regulating the tyrosinase gene. Research indicates that nitric oxide induces tyrosinase gene expression, leading to increased tyrosinase activity and protein levels. This up-regulation is mediated through the cGMP pathway, suggesting that enhancing tyrosinase gene expression via this pathway is a primary mechanism for melanogenesis. Additionally, nitric oxide affects the oxidations of l-dopa and dopamine, influencing the activity of enzymes like tyrosinase and peroxidase, which are involved in melanin synthesis.. 41 –43
In this study, IB complex decreased tyrosinase activity, NO contents, cAMP levels, and protein expression of p-PKA/PKA, p-CREB/CREB, MITF, TRP-1, and TRP-2. These results propose that IB complex could protect melanogenesis via downregulation of PKA, CREB, MITF, and TRPs pathways.
In conclusion, Indian gooseberry and barley sprout complex (IB complex) supplementation prevented skin damage such as dryness, wrinkle formation, and oxidative stress induced by UVB irradiation. IB complex ameliorated skin moisturization via the activation of HA and ceramide synthesis, and suppressed wrinkle formation via the repression of JNK, c-Fos, c-Jun, MMPs, and COX-2, and the promotion of collagen synthesis pathways. Moreover, IB complex inhibited melanogenesis via the reduction of cAMP, CREB, and MITF (Fig. 8). In accordance with these findings, we suggest that the supplementation with IB complex might be beneficial foods for protecting against skin photoaging.
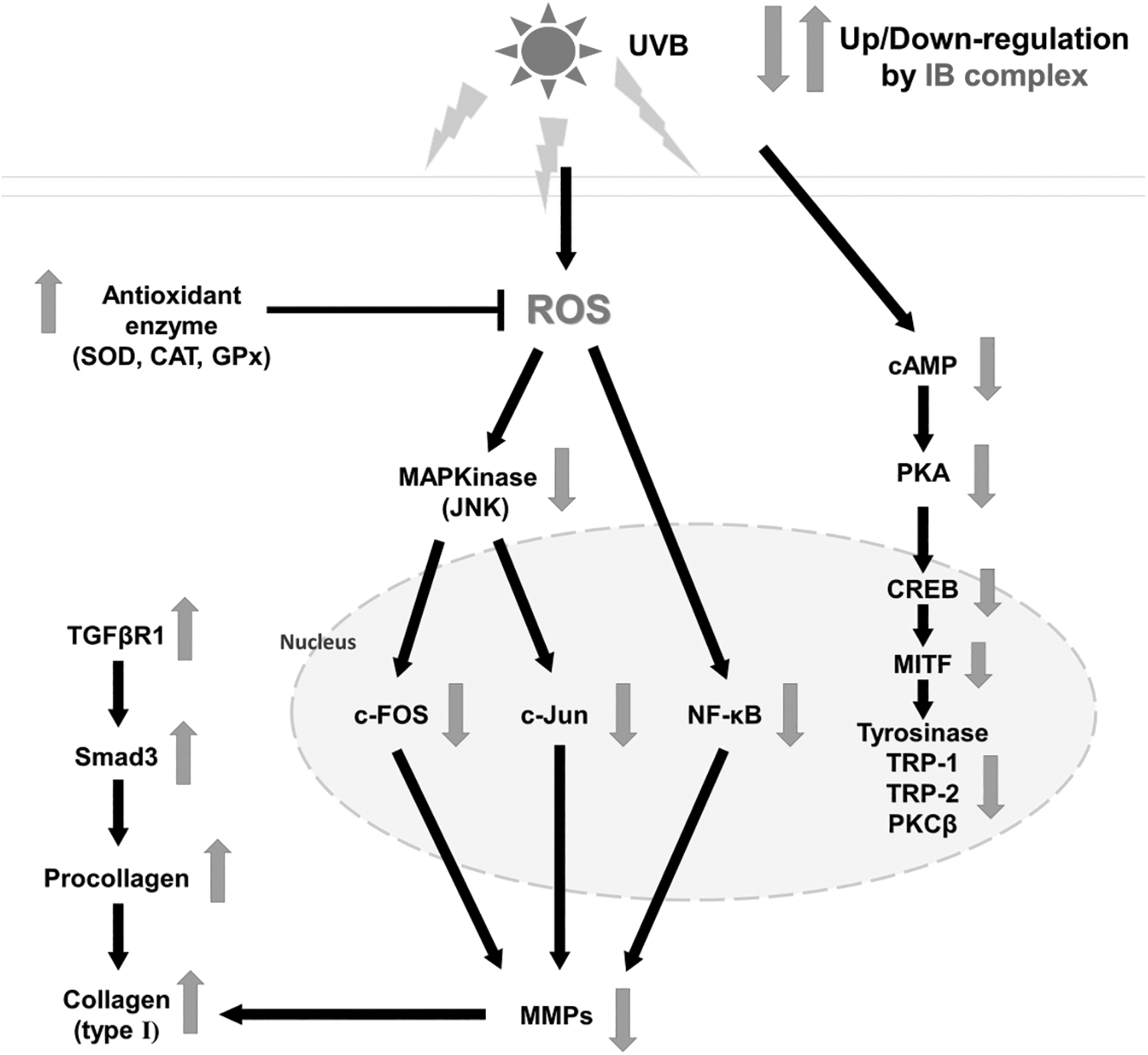
A schematic diagram showing the possible mechanism of IB complex in UVB-induced oxidative stress and photoaging in the skin.
Footnotes
AUTHORs' CONTRIBUTIONS
M.L.: conceptualization; investigation; data curation; visualization; writing—original draft; and writing—review and editing. D.K.: investigation; data curation; and visualization. M-R.P.: writing—review and editing. S.K.: writing—review and editing. J-L.K.: writing—review and editing. J.W.L.: writing—review and editing. O-K.K.: conceptualization; data curation; writing—original draft; and writing—review and editing. J.L.: conceptualization; data curation; writing—original draft; and writing—review and editing.
DISCLOSURE STATEMENT
M-.R.P., S.K., J-L.K., and J.W.L. are the current employees of a commercial company which holds a patent for the IB complex. M-.R.P., S.K., J-L.K., J.W.L., and J.Y. are listed as inventors.
FUNDING INFORMATION
No funding was received for this article.