
Abstract
Gastric cancer is the fifth most common cancer globally and the third leading cause of cancer-related mortality. Existing treatment strategies for gastric cancer often present numerous side effects. Consequently, recent studies have shifted toward devising new treatments grounded in safer natural substances. α-Pinene, a natural terpene found in the essential oils of various plants, such as Lavender angustifolia and Satureja myrtifolia, displays antioxidant, antibiotic, and anticancer properties. Yet, its impact on gastric cancer remains unexplored. This research assessed the effects of α-pinene in vitro using a human gastric adenocarcinoma cell-line (AGS) human gastric cancer cells and in vivo via a xenograft mouse model. The survival rate of AGS cells treated with α-pinene was notably lower than that of the control group, as revealed by the 3-(4,5-dimethylthiazol-2-yl)-2,5 diphenyltetrazolium bromide assay. This decline in cell viability was linked to apoptosis, as verified by 4′,6-diamidino-2-phenylindole and annexin V/propidium iodide staining. The α-pinene–treated group exhibited elevated cleaved-poly (ADP-ribose) polymerase and B cell lymphoma 2 (Bcl-2)–associated X (Bax) levels and reduced Bcl-2 levels compared with the control levels. Moreover, α-pinene triggered the activation of extracellular signal-regulated kinase, c-Jun N-terminal kinase, and p38 within the mitogen-activated protein kinase (MAPK) pathway. In the xenograft mouse model, α-pinene induced apoptosis through the MAPK pathway, devoid of toxicity. These findings position α-pinene as a promising natural therapeutic for gastric cancer.
INTRODUCTION
Gastric cancer, originating from the glandular epithelium of the gastric mucous membrane, ranks as the fifth most common cancer worldwide and the third leading cause of cancer-related mortality. 1,2 Often, gastric cancer remains asymptomatic in its early stages, leading to diagnoses typically made during advanced or metastatic stages. 3,4 Current treatment modalities include surgery, radiation therapy, and chemotherapy. Yet, there is a pressing need for advancements in treatment approaches to mitigate the progression, metastasis, and recurrence of gastric cancer and address the side effects of established anticancer drugs. 5,6 Consequently, research focusing on relatively safe natural materials is gaining momentum. 7
α-Pinene is the most prevalent natural terpene in the plant kingdom. Characterized by its alkene properties and a 4-membered ring structure, α-pinene is present in the essential oils of various plants, including Lavender angustifolia, Satureja myrtifolia, Canarium tramdenanum, and Rosmarinus officinalis. 8,9 Furthermore, α-pinene has proven effective as a bronchodilator in asthma treatment and has antioxidant and antibiotic properties. 10,11 Although recent findings have highlighted the anticancer activity of α-pinene, unveiling its roles in regulating cell cycle progression, inhibiting inflammation, and promoting apoptosis, 12,13 research investigating its signaling mechanism and anticancer efficacy in gastric cancer remains scarce.
Apoptosis, a type of programmed cell death, 14 is primarily governed by B cell lymphoma 2 (Bcl-2) family of proteins, which segregate into pro-apoptotic and anti-apoptotic proteins. 15 The pro-apoptotic proteins, encompassing Bcl‑2–associated X (Bax), BH3-interacting domain death agonist (Bid), and Bcl-2–associated agonist of cell death (Bad), disrupt the outer mitochondrial membrane, inducing apoptosis. Conversely, anti-apoptotic proteins, namely Bcl-2 and B cell lymphoma—extra large (Bcl-xL), maintain the mitochondrial membrane's integrity, thus impeding apoptosis. 16 –18 In DNA damage or cellular stress circumstances, pro-apoptotic protein expression escalates, augmenting membrane permeability. This results in the release of cytochrome c, which subsequently forms a caspase complex, deactivating anti-apoptotic proteins and initiating apoptosis through a caspase-mediated reaction that fragments poly (ADP-ribose) polymerase (PARP). 19 –21
The mitogen-activated protein kinase (MAPK) cascade is pivotal in apoptosis and cellular stress responses, differentiation, and growth. 22 Proteins within this pathway, such as p38, c-Jun N-terminal kinase (JNK), and extracellular signal-regulated kinase (ERK), orchestrate various biological functions, including intracellular communication. 23 Both JNK and p38 respond to stressors like DNA damage and osmotic pressure fluctuations, enhancing pro-apoptotic protein expression, suppressing anti-apoptotic protein expression, and modulating the MAPK pathway to trigger apoptosis. 24 –26 Although ERK expression is traditionally linked to cellular survival, growth, and differentiation, emerging studies indicate its association with apoptosis and autophagy. 27 However, such an association has not been investigated so far.
Hence, this study aimed to assess the impact of α-pinene on human gastric cancer a human gastric adenocarcinoma cell-line (AGS) cells and to explore its potential interactions with the MAPK pathway.
MATERIALS AND METHODS
Materials and reagents
(+)-α-Pinene ≥99% (Fig. 1A), dimethyl sulfoxide (DMSO), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), and 4′,6-diamidino-2-phenylindole (DAPI) were purchased from Sigma-Aldrich (St. Louis, MO, USA). DMSO served as a solvent for α-pinene and as the treatment for the control group. The fluorescein isothiocyanate (FITC)-Annexin-V Detection Kit was obtained from BD Pharmingen™ (San Diego, CA, USA). Various antibodies and their sources include anti-β-actin (mouse, sc-47778, 1:2000) and secondary antibody mouse IgG (mouse, sc-516102 1:3000) from Santa Cruz Biotechnology, Inc., (Dallas, TX, USA); anti-Bcl-2 (rabbit, NB100-56098, 1:1000) from Novus (Littleton, CO, USA); and anti-PARP (rabbit, # 9542, 1:1000), anti-Bax (rabbit, # 2772, 1:700), anti-ERK (rabbit, # 9102, 1:1000), anti-p-ERK (rabbit, # 9101, 1:1000), anti-JNK (rabbit, # 9252, 1:700), anti-p-JNK (rabbit, # 4668, 1:500), anti-p38 (rabbit, # 9212, 1:1000), anti-phosphorylated p38 (p-p38; rabbit, # 9211, 1:1000), and secondary antibody rabbit IgG (rabbit, # 7074, 1:2000) from Cell Signaling Technology (Beverly, MA, USA).
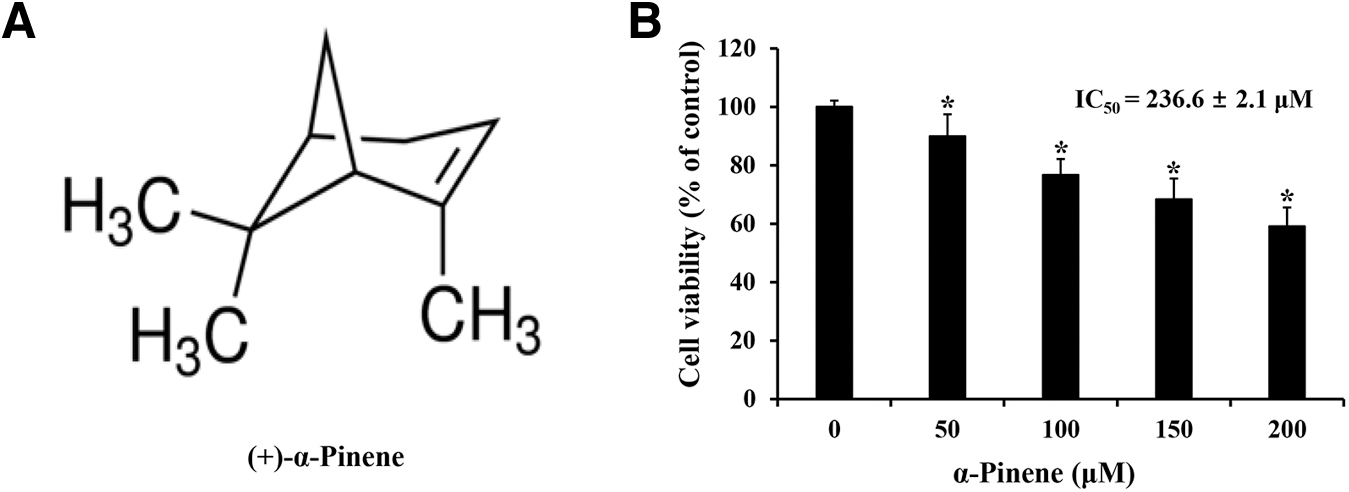
Effects of α-pinene on AGS cell viability.
Cell culture conditions
AGS cells, sourced from the Korean Cell Line Bank (Seoul, Republic of Korea), were maintained in RPMI-1640 (Welgen, Gyeongsan, Republic of Korea) medium supplemented with 5% fetal bovine serum (FBS; Welgen) and 1% streptomycin/penicillin (Gibco BRL, Grand Island, NY, USA), incubated at 37°C in 5% CO2. Upon reaching 80–90% confluence, subculturing was performed, and the medium was replaced every 2–3 days.
MTT assay
AGS cells were seeded at 2 × 104 cells/mL on a 96-well plate and incubated for 24 h at 37°C. Then, the medium was removed and treated with α-pinene (0, 50, 100, 150, and 200 μM) for 24 h at 37°C. The medium was removed, and 40 μL/well MTT solution was applied for 90 min. The MTT solution was removed, and 100 μL/well DMSO was added to dissolve formazan. Then, absorbance was measured at 595 nm using an ELISA microplate reader to determine cell viability (Bio‑Rad Laboratories, Inc., Hercules, CA, USA).
DAPI staining
AGS cells were seeded at 2 × 105 cells/mL in 60-mm dishes and cultured for 24 h. The medium was removed, and α-pinene was added at 0, 100, or 200 μM, followed by incubation at 37°C for 24 h. After removing the medium, the cells were fixed with 4% paraformaldehyde for 15 min. Then, staining was performed for 1 min at 20°C with 2 mL/dish of DAPI. Afterward, the DAPI solution was removed, and a fluorescence microscope (Zeiss Fluorescence Microscope, Thornwood, NY) was used at 200 × magnification to observe morphological changes (apoptotic bodies) nuclear penetrability of the DAPI stain.
Annexin-V/propidium iodide staining
AGS cells were seeded into 75 cm2 flasks and cultivated for 24 h in an incubator. The cells were treated with 0, 100, or 200 μM α-pinene at 37°C for 24 h. Trypsin-ethylenediaminetetraacetic acid (EDTA) was used to harvest the cells, and the cell pellet was centrifuged at 260 g for 5 min at 4°C and suspended in 1 × binding buffer. The cell samples were stained with annexin-V and propidium iodide (PI) for 15 min at 20°C, and the degree of apoptosis was quantified using a FACSCalibur™ flow cytometer (BD Biosciences, Franklin Lakes, NJ, USA).
Western blot analysis
A constant number of AGS cells was cultivated for 24 h in a 75 cm2 flask, and medium supplemented with 0, 100, and 200 μM α-pinene was added at 37°C for 24 h. The cells were treated with trypsin-EDTA and centrifuged at 260 g and 4°C for 5 min, and the cell pellet was treated with cell lysis buffer (Invitrogen, Carlsbad, CA, USA) for 20 min at 4°C. The supernatant obtained after centrifugation for 5 min at 5920 g and 4°C was the cell lysate. The proteins in the cell lysate were quantified using a Bradford protein assay. The proteins were resolved using 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred to a nitrocellulose membrane. The membrane was blocked for 2 h at 20°C using 5% skim milk or bovine serum albumin and incubated overnight with the primary antibody at 4°C.
Then, the membrane was incubated with anti-mouse IgG and anti-rabbit IgG as secondary antibodies at 20°C for 2 h. Next, enhanced chemiluminescence detection reagents (Pierce, Rockford, IL, USA) were used for protein detection, and results were quantified using ImageJ Launcher (NCBI).
Establishment of a xenograft model
BALB/c nude female mice (4 weeks old, 18–22 g) were purchased from Nara Biotec (Seoul). The animal experiment was approved by Kongju National University's Institutional Animal Care and Use Committee (No. KNU_2021-03). All animal experiments were performed at the SPF Animal Laboratory of Kongju National University. The mice were maintained under a 12 h day/night cycle at 23°C ± 3°C and 40% ± 10% humidity. Since AGS cells were identified as a suitable gastric adenocarcinoma model for gastric cancer xenograft modeling, we established a xenograft model using AGS cells to confirm the in vivo anticancer efficacy of α-pinene in human gastric cancer. To establish the xenograft model, 1 × 107 AGS cells/mL in phosphate-buffered saline (PBS) with 20% FBS was administered subcutaneously into both shoulders of the mice.
After tumor formation on the shoulders of mice, the solid tumors were excised, and 3-mm tumor sections were implanted subcutaneously into the experimental group. After 10 days of tumor growth, the mice were divided randomly into PBS group (control group, n = 5) and α-pinene group (200 mg/kg, n = 5). The mice were orally administered PBS or α-pinene (200 mg/kg) every other day for 12 days. 28 The tumor was measured every second day using Vernier calipers (Mitutoyo, Kawasaki, Japan). Tumor volume was calculated according to the following formula: Volume (mm3) = 0.5 × (wide) 2 × length. 28,29 At the end of the experiment, all mice were killed using CO2 gas (30% per min, 3 min), and tumors were excised and weighed.
Hematoxylin and eosin staining
The liver and kidney tissues were fixed with 10% formaldehyde, embedded into paraffin, and cut into 4-μm sections. The sections were stained with hematoxylin and eosin (H&E) and observed under an optical microscope (BX41; Olympus Co., Tokyo, Japan) at 400 × magnification to identify the toxic effects of α-pinene in the liver and kidneys.
Terminal deoxynucleotidyl transferase dUTP nick-end labeling staining
The tumor tissue was fixed with 10% formaldehyde, embedded into paraffin, cut into 5-μm sections, and stained using the DeadEnd™ Colorimetric transferase dUTP nick-end labeling (TUNEL) system (Promega, Madison, WI, USA). The stained tumor tissues were observed using an optical microscope at 400 × magnification.
Statistical analysis
Data are expressed as mean ± standard deviation. Group comparisons were performed using Dunnett's t-test following a one-way analysis of variance. Differences were considered statistically significant at P < .05.
RESULTS
α-Pinene reduced the viability of AGS cells
The MTT assay assessed the viability of AGS cells after α-pinene treatment for 24 h at 0, 50, 100, 150, and 200 μM. The mean survival rates of AGS cells were 89.9%, 76.7%, 68.3%, and 59.1% after treatments with α-pinene at 50, 100, 150, and 200 μM, respectively. The IC50 value (concentration required for 50% growth inhibition) of α-pinene for AGS cell viability was determined to be 236.6 ± 2.1 μM (Fig. 1B). Therefore, α-pinene decreased AGS cell viability in a concentration-dependent manner.
α-Pinene induced apoptosis in AGS cells
To ascertain if the reduction in AGS cell viability from α-pinene treatment was owing to apoptosis, DAPI staining was performed. Compared with that in the control group, the mean apoptotic body rates increased by 11.8% and 38.3% in groups treated with 100 and 200 μM α-pinene, respectively (Fig. 2A). Moreover, to quantify apoptosis, cells were stained with annexin-V and PI for fluorescence-activated cell sorting analysis. Apoptosis rates in the 100 and 200 μM α-pinene-treated groups were 24.8% and 34.0%, respectively (Fig. 2B). When AGS cells were treated with 0, 100, and 200 μM α-pinene for 24 h, cleaved-PARP and Bax levels increased, whereas Bcl-2 levels decreased compared with the control levels (Fig. 3).
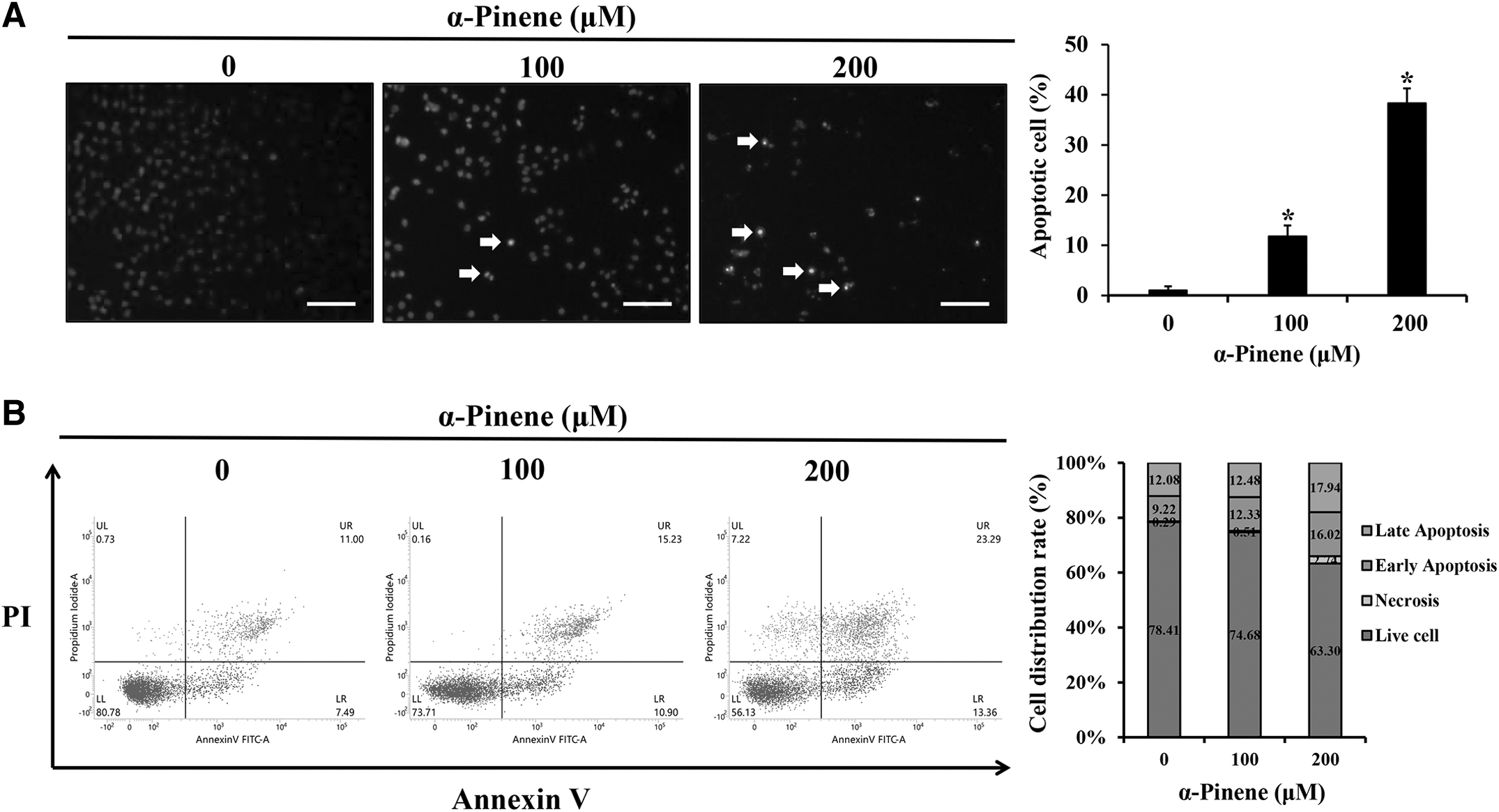
Effects of α-pinene on apoptosis in AGS cells.
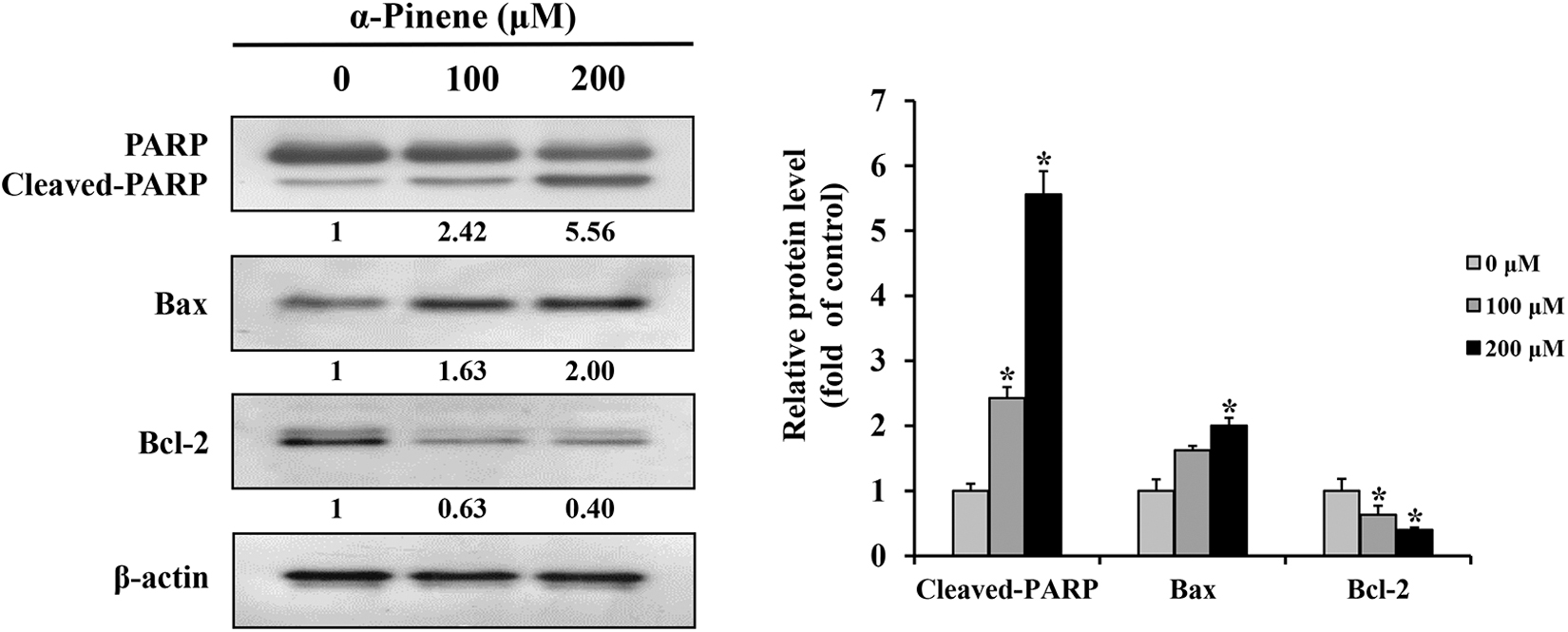
Effects of α-pinene on the regulation of apoptotic protein levels in AGS cells. AGS cells were subjected to treatment with α-pinene (0, 100, and 200 μM) for 24 h, and the protein expression levels of PARP, Bax, and Bcl-2 were measured by western blotting. β-actin was used as loading control, and the quantification was performed using ImageJ. Control cells were subjected to treatment with DMSO. Data are presented as mean and SD for three samples. *P < .05 compared with the control group. Bax, Bcl‑2–associated X; Bcl-2, B cell lymphoma 2; PARP, poly (ADP-ribose) polymerase.
α-Pinene activated the MAPK pathway in AGS cells
To investigate the influence of α-pinene on the AGS cell apoptotic pathway, ERK, JNK, and p38 levels related to the MAPK pathway were analyzed. Cells treated with 100 and 200 μM α-pinene exhibited decreased ERK, JNK, and p38 levels but increased p-ERK, p-JNK, and p-p38 levels compared with the control levels (Fig. 4).
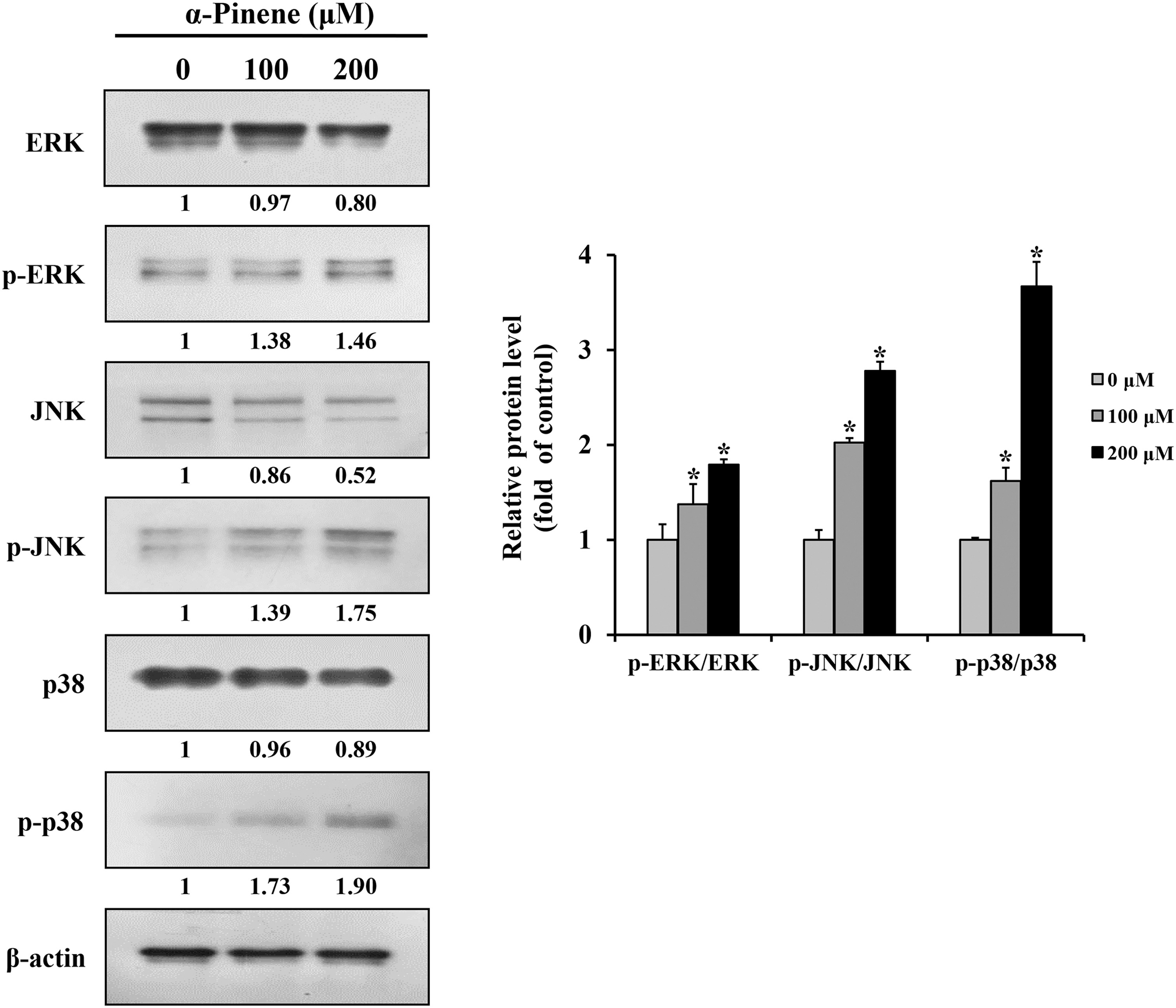
Effects of α-pinene on MAPK pathway in AGS cells. AGS cells were subjected to treatment with α-pinene (0, 100, and 200 μM) for 24 h, and the protein expression levels of ERK, p-ERK, JNK, p-JNK, p38, and p-p38 were measured by western blotting. β-actin was used as loading control, and the quantification was performed using ImageJ. Control cells were subjected to treatment with DMSO. Data are presented as mean and SD for three samples. *P < .05 compared with the control group. ERK, extracellular signal-regulated kinase; JNK, c-Jun N-terminal kinase; MAPK, mitogen-activated protein kinase.
α-Pinene inhibited tumor growth in the xenograft mouse model
The in vitro experiments revealed that α-pinene induced apoptosis in the AGS gastric cancer cell line, inhibiting cell survival. To validate this effect in vivo, a xenograft model was used. Tumors in the α-pinene group showed a 28.2% reduced volume (Fig. 5A) and a 13.3% reduced weight (Fig. 5B) compared with those in the PBS group, although the difference was not statistically significant (P > .05). Furthermore, no significant weight differences were observed between the two groups (Fig. 5C).

Effects of α-pinene on xenograft tumors in vivo. BALB/c nude mice bearing the adenocarcinoma gastric cell line (AGS) as a xenograft were administered α-pinene (0, 200 mg/kg) every other day for 12 days, following which, the
α-Pinene did not result in histopathological changes
After administering α-pinene into the xenograft mouse model, the livers and kidneys of the control and treatment groups were evaluated using H&E staining. There were no significant differences between the two groups, indicating that no histopathological changes occurred in response to α-pinene treatment (Fig. 5D).
α-Pinene induced apoptosis in tumor cells via the MAPK pathway
Expression changes in MAPK pathway proteins in response to α-pinene in tumor tissue cells were confirmed through TUNEL staining and western blotting. Compared with that in the control group, the apoptosis rate significantly increased to 4.6% in the α-pinene group (Fig. 6A). Furthermore, Bax expression levels were significantly higher, whereas Bcl-2 expression levels were significantly lower in tumors treated with α-pinene compared with those in the control group tumors (Fig. 6B); levels of the MAPK-related proteins p-ERK, p-JNK, and p-p38 increased significantly after α-pinene treatment (Fig. 6C).
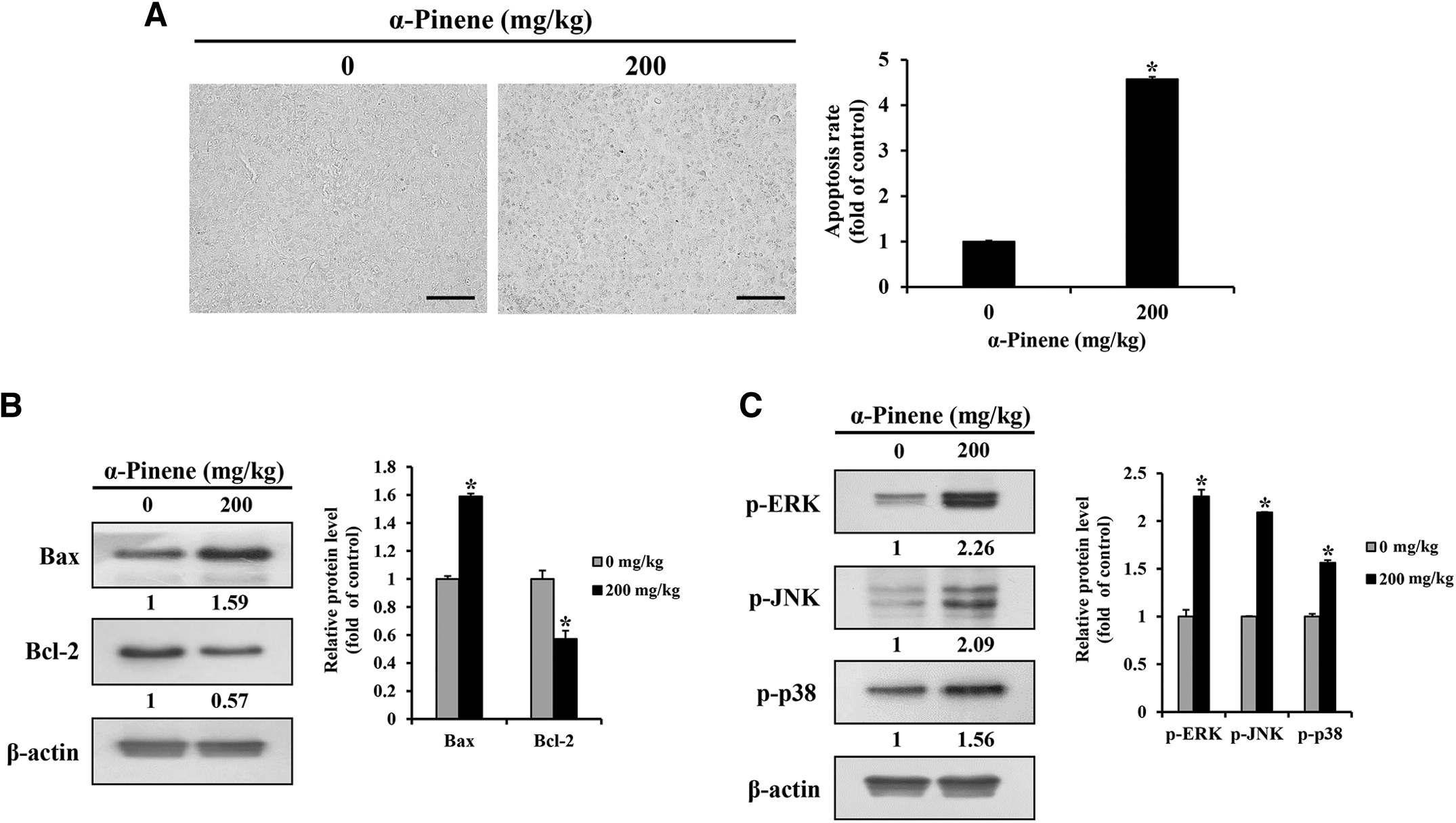
Effects of α-pinene on apoptosis in vivo.
DISCUSSION
Although the anticancer and tumor inhibitory effects of α-pinene through apoptosis induction have been identified in liver 30 and ovarian cancers, 31 studies on its effects on human gastric cancer cells remain scant. This study demonstrates that α-pinene decreases the survival rate of the human gastric cancer cell line AGS and that this anticancer effect is primarily owing to the induction of apoptosis via the MAPK pathway. This anticancer effect was further corroborated through in vivo assays.
The principle behind the MTT assay is that only viable cells convert MTT into purple formazan through active metabolism. 32 The MTT assay showed a marked decrease in the mean cell survival rate as the α-pinene concentration increased from 50 to 200 μM. In HeLa cells (a human cervical cancer cell line) treated with 10, 20, 40, 80, 160, 320, and 640 μmol/L α-pinene for 24 h, the IC50 value stood at 46.87 ± 0.78 μmol/L, indicating a reduction in HeLa cell survival owing to α-pinene. 33 In addition, when α-pinene was applied at 0.125, 0.5, 2, and 8 mg/L to the human liver cancer cell line BEL-7402 for 24, 48, and 72 h, 79.3% cell growth inhibition occurred in the group treated with 8 mg/L α-pinene for 72 h. 34 These findings cumulatively suggest a concentration-dependent decrease in AGS cell viability owing to α-pinene.
During apoptosis, DNA fragmentation occurs alongside an increase in cell membrane permeability. Consequently, apoptotic bodies are discernible through intensified DAPI staining. 35 This study found an increase in the apoptosis rate with higher concentrations of α-pinene, as evidenced by DAPI staining. Furthermore, annexin-V/PI staining was used to quantify apoptosis in α-pinene–treated AGS cells. In general, phosphatidylserine is located within the cell membrane but shifts to the exterior during apoptosis, facilitating apoptosis detection via annexin-V. 36 In addition, the cell membrane deteriorates in the latter stages of apoptosis, allowing PI to differentiate between early and late apoptosis. 37 This study noted rising early and late apoptosis rates with increasing α-pinene concentrations from 0 and 100 to 200 μM.
In previous research, 2.5 μM α-pinene increased apoptosis when applied to the prostate cancer cell lines PC-3 and DU145 for 24 h. 38 Based on this previous research and the current results, the α-pinene–induced decrease in the AGS cell survival rate can likely be attributed to apoptosis.
The space between mitochondrial inner and outer membranes houses pro- and anti-apoptotic proteins. When the permeability of this mitochondrial membrane is altered—due to an increase in pro-apoptotic proteins or a decrease in anti-apoptotic ones—cytochrome c is released, triggering caspase activation and apoptosis. 39 –42 Moreover, as apoptosis progresses, there is a notable disruption in PARP, a significant entity in DNA repair and transcription regulation. 43 In this study, higher levels of cleaved-PARP and Bax were observed in α-pinene–treated AGS cells than in the control group, whereas Bcl-2 levels diminished. Earlier studies on cervical cancer cells treated with α-pinene reported similar observations, 33 implying that α-pinene enhances mitochondrial membrane permeability in AGS cells, triggering apoptosis.
Prior research indicates that α-pinene can exert anticancer effects by activating natural killer cells via the ERK/AKT 44 and PI3K/AKT/NF-κB pathways in human breast cancer cells. 45 Nevertheless, the anticancer influence of the MAPK pathway in human gastric cancer remains elusive. The MAPK pathway, instrumental in various cellular processes, including infections, cell cycle progression, and aging, 22 plays a pivotal role in cancer progression. 46 Whereas activated JNK functions as a pro-apoptotic kinase, p38 activation significantly affects cell growth inhibition and apoptosis. 47,48 JNK and p38 activation also induces the expression of apoptosis-related proteins. 49 Conversely, although ERK activation typically supports cell survival, apoptosis could be triggered by obstructing ERK activation. 50
Of note, in most cancers, p53 activation is diminished, 51 and ERK activation promotes p53 activity, leading to reduced cell cycle progression and apoptosis. 52,53 This study found decreased ERK, JNK, and p38 expression levels in AGS cells treated with α-pinene while levels of their phosphorylated counterparts increased. These results suggest that in AGS, α-pinene induces transcriptional activators associated with apoptosis induction through the activation of JNK and p38, and activation of ERK induces apoptosis through its role in supporting these transcriptional activities.
A xenograft model was used to delve deeper into the in vivo anticancer effects of α-pinene. In addition, the safety of α-pinene was evaluated by confirming acute inflammation (neutrophils) and chronic inflammation (fibrosis) in the liver and observing abnormalities in the glomeruli and tubules, which are most vulnerable to toxicity, in the kidney. Compared with those in the control group, 200 mg/kg α-pinene administration resulted in reduced tumor volume and weight, devoid of noticeable toxicity. Previous studies echo these results, showing notable tumor volume reduction after α-pinene administration in prostate cancer. 38 Moreover, in liver cancer, α-pinene compounds inhibited tumor growth by up to 52.7% at specific dosages. 28 Considering these past observations and those of this study, it is evident that α-pinene exhibits anticancer properties against gastric cancer tumors.
Apoptosis, as indicated by the elevated number of TUNEL-positive cells, correlated with decreased tumor volume and weight in α-pinene–administered mice. Furthermore, the tumors from the α-pinene group manifested an increased Bax expression, decreased Bcl-2 expression, and heightened levels of MAPK pathway-associated proteins. Previous investigations on human cervical cancer HeLa cell xenograft tumors corroborate these findings, underscoring in vivo apoptosis induction through α-pinene. 33 Collectively, these results advocate that the observed tumor volume and weight reductions after α-pinene administration are likely owing to apoptosis induction via the MAPK pathway. However, future experiments focusing on the mechanism of apoptosis induction by α-pinene in other human gastric cancer cells are required. Also, additional studies are crucial to validate its enduring antitumor efficacy against human gastric cancer and its in vivo effects at different concentrations.
In conclusion, α-pinene diminishes the survival rate of human gastric cancer AGS cells in a concentration-dependent manner by inducing apoptosis. This is evidenced by an increase in cleaved-PARP and Bax expression and a decrease in Bcl-2 expression. Both in vitro and in vivo experiments indicate that the α-pinene–mediated apoptosis is channeled through MAPK pathway-related proteins. These results suggest that α-pinene can induce apoptosis in human gastric cancer cells AGS by activating transcriptional activators involved in apoptosis progression by regulating the expression of proteins related to the MAPK pathway. Thus, these findings indicate a promising therapeutic potential for α-pinene in treating human gastric cancer.
Footnotes
AUTHORs' CONTRIBUTIONS
E-J.H.: Visualization, methodology, data curation, software, writing—original draft. E-Y.C.: Data curation, investigation, conceptualization, writing—review and editing. S-J.J.: Validation, visualization, methodology, writing—review and editing. S-W.L.: Visualization, investigation, software. J-M.M.: Investigation, methodology, software. G-H.J.: Validation, software, supervision, data curation. J-H.L.: Methodology, conceptualization, validation, supervision. S-H.H.: Investigation, conceptualization, validation, software. S-H.J.: Software, data curation, validation. M-S.Y.: Writing—review and editing, supervision, conceptualization. J-Y.J.: Resources, conceptualization, validation, supervision, project administration, writing—review and editing.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This study was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2021R1A2C1010912).