
Abstract
This study aimed to explore the curative effect of curcumin on liver fibrosis and its correlation with the gut–liver axis in animal models. Histological staining was utilized to conduct histological analysis of the liver and intestine. An automatic biochemical analyzer or enzyme-linked immunosorbent assay system was utilized for analyzing the biochemical indexes in mice. Western blotting was employed to examine the level of relevant proteins. Furthermore, 16S rRNA high-throughput sequencing was performed to explore the impact of curcumin on intestinal microorganisms in rats with liver fibrosis. Ultrahigh-performance liquid chromatography with quadrupole-orbitrap mass spectrometry was utilized to analyze the effect of curcumin on rat feces metabolites. Our results showed that curcumin reduced the formation of collagen fibers caused by carbon tetrachloride in a dose-dependent manner. In addition, curcumin was able to restore intestinal permeability in rats with liver fibrosis. By adopting α diversity analysis (Chao 1 index, Shannon index, and Simpson index), we observed that both the diversity and the abundance of intestinal flora in rats with liver fibrosis were increased. The principal component analysis diagram demonstrated that curcumin could enhance the abundance and diversity of intestinal flora, and also restore the composition of model rat flora, which was similar to that in normal rats, thereby correcting the imbalance of flora in rats with liver fibrosis. In addition, curcumin regulated feces metabolites and their signaling pathways, including glycerophospholipid metabolism, pantothenate and CoA biosynthesis. Our findings suggest that curcumin exhibits antiliver fibrosis effects, and its antiliver fibrosis effects might correlate with gut–liver axis.
INTRODUCTION
Liver fibrosis is a common disease that is manifested as irregular deposition of fibrotic tissue in the liver. 1,2 Without effective treatment, the progression of the disease will lead to the formation of fibrous nodules of the liver and the destruction of the normal liver cells, eventually progressing into cirrhosis, liver failure, and even liver cancer. 3,4 Therefore, studying antifibrosis drugs is of great clinical significance. The liver and the gut are closely interconnected through the portal vein. 5 The intestinal barrier plays a crucial role as a physical defense within the intestinal tract. When this barrier is compromised, the liver becomes susceptible to poison originating from the gut, including lipopolysaccharide (LPS), bile acid (BA), peptidoglycan, and ethanol. 6 –8
There are >100 trillion microorganisms in the human intestinal tract, including one archaea (Euryarchaeota) and five bacterial phyla (Actinomycetes, Verrucomicrobia, Bacteroidetes, Proteobacteria, and Firmicutes). 9 These trillions of microorganisms constitute the intestinal microbiota, which varies among individuals and is crucial for well-being. In recent years, the intestinal microbiota was observed to not only influence normal functions of the intestinal tract but also closely correlate with many other nonintestinal diseases such as liver diseases. 10 –12
The gut–liver axis is a concept that interprets the complicated correlations between the intestinal tract and the liver; it is comprised of intestinal barrier, microbiota, portal circulation, Bas, and the gut immune system. The gut–liver axis is critical for overall well-being, while disruptions in this axis can contribute to various health conditions. 13,14 For instance, dysbiosis (imbalance in gut microbes) might lead to liver diseases such as fatty liver or inflammation. Conversely, liver diseases can influence gut health, impacting nutrient absorption and microbial balance. In recent studies, researchers discovered that modulating the gut–liver axis might facilitate the treatment of liver disease, implying the significance of the gut–liver axis. 15 –17
Curcumin, a polyphenol compound with low relative molecular weight, was first isolated from Curcuma longa L1 in 1870. Subsequently, its chemical structure was clarified in 1910, and since then significant progress has been made in the study of its physiological and pharmacological effects. Studies have demonstrated that curcumin displays a broad spectrum of pharmacological activities, encompassing anti-inflammatory, antioxidant, lipid-regulating, and anti-infective properties, characterized by low toxicity and minimal adverse reactions. While the use of curcumin in the Oriental medical system has a long history, western medicine has also increasingly recognized its therapeutic potential. Curcumin stands as one of the most widely sold natural edible pigments globally, having gained approval as a food additive from various government authorities.
Curcumin is attractive to researchers not just as a nonsteroidal anti-inflammatory drug, but also for its wide range of preventive properties against various diseases. For instance, studies discovered that curcumin displayed a good antibacterial effect, with turmeric extract being shown to inhibit the growth of Escherichia coli in vitro. Furthermore, curcumin has been found to induce apoptosis-like responses in E. coli, whereas other studies have demonstrated that curcumin improves the curative effect of triple therapy on dyspepsia.
In the last few years, there have been reports indicating that curcumin possesses hepatoprotective effects, positioning it as a promising candidate for the treatment of liver fibrosis. However, the detailed mechanism of antiliver fibrosis of curcumin remains largely unknown. Thus, we performed this study to investigate the antiliver fibrosis effect of curcumin, and more importantly, to explore the potential mechanism and correlation with the gut–liver axis based on our previous findings and speculations. The innovation of this study lies in the interpretation that curcumin contributes to the beneficial bacteria production, and the alleviation of liver fibrosis is correlated with the recovery of intestinal flora and regulation of feces metabolites.
MATERIALS AND METHODS
Study design
To elucidate the potential mechanism by which curcumin treats liver fibrosis within the gut–liver axis, we initiated this animal study. Initially, we induced a liver fibrosis animal model using carbon tetrachloride (CCl4) and subsequently administered curcumin to examine its antifibrotic effects. Next, 16S rRNA sequencing technology was used to detect the impacts of curcumin on intestinal microbiota in rats with liver fibrosis, along with the intestinal barrier integrity in each group of rats. Finally, metabolomics technology was used to investigate the metabolic intermediates and metabolic routes through analyzing rat feces.
Animals and drug administration
All animal experiments adhered to the guidelines for Animal Experimentation at Nanjing University of Chinese Medicine, and the experimental protocol received approval from The Ethics Committee of Animal Experiments at Nanjing University of Chinese Medicine (Protocol No.: 201813079). Male Sprague-Dawley rats (SLAC, Shanghai, China) were procured and housed in a specific pathogen-free facility. Before the study, the rats were fed ad libitum for 1 week. They were then randomly assigned to one of five groups, each comprising eight rats: The control group was subjected to intraperitoneal injections of 1 mL/kg of olive oil twice a week for a duration of 8 weeks.
In contrast, both the model group and the curcumin treatment group (Cur) were administered intraperitoneal injections of 1 mL/kg of a 50% CCl4 and olive oil solution twice a week over the same 8-week period. The curcumin treatment groups received oral gavage simultaneously with curcumin infusion at 100, 200, and 400 mg/kg six times a week, respectively (Cur100, Cur200, Cur400), from the third week, while both the control group and the model group received oral gavage of saline. 18 The interventions lasted for 6 weeks. The daily administration and curcumin dosage were based on previously reported studies.
In a parallel experiment, 56 additional rats were randomly allocated to 7 groups, each consisting of 8 rats. The control group was subjected to intraperitoneal injections of 1 mL/kg of olive oil twice a week for a duration of 8 weeks. In contrast, the remaining six groups were subjected to intraperitoneal injections of 1 mL/kg of a 50% CCl4 and olive oil solution twice a week for the same 8-week duration.
The effective microorganism (EM) treatment group received oral gavage simultaneously with 2 mL clostridium caseinate live bacteria lyophilized powder (2 × 1010 CFU/mL) six times a week from the third week; dextran sulfate sodium (DSS) treatment group received oral gavage simultaneously with 4% DSS solution (1 mL/kg) six times a week from the third week; Penicillin G treatment group received oral gavage simultaneously with 2 mL Penicillin G solutions six times a week from the third week; DSS + Cur treatment group received oral gavage simultaneously with 4% DSS solution (1 mL/kg), and curcumin infusion (200 mg/kg) six times a week from the third week; G + Cur treatment group received oral gavage simultaneously with 2 mL Penicillin G solutions and curcumin infusion (200 mg/kg) six times a week from the third week.
Toward the conclusion of the experiments, rats underwent an overnight fasting period before being carbon dioxide euthanized. Serum samples were obtained by collecting and centrifuging blood samples. Feces and intestinal/liver tissues were carefully gathered and promptly stored at −80°C for further analysis. All animals were kept in environments free from pathogens, with unlimited access to both food and water. Ethical approval for the animal experiments conducted in this study was obtained from the Ethics Committee, and strict adherence to all relevant regulations concerning the ethical treatment of animals was ensured.
To ensure the ethical treatment of animals in our study, Sprague-Dawley rats were euthanized using the intraperitoneal (i.p.) or intravenous (i.v.) injection of pentobarbital sodium at a dose of 100–150 mg/kg. This method is widely accepted as the preferred method for animal euthanasia as it causes the animal to lose consciousness and enter anesthesia, resulting in a humane and painless death.
Histological analyses of liver and intestine tissues
The liver and intestine tissues were subjected to histological analyses by various staining techniques, including hematoxylin and eosin (H&E) staining, Masson's trichrome staining, and Sirius Red staining. The sections were then examined using an optical microscope (Nikon Eclipse Ci, Japan) to observe tissue morphology and identify any histological abnormalities (200 × ).
Biochemical indicators analysis
The rat blood was collected and centrifuged at low temperature (4°C, 1,200 g, 10 min), then the supernatant was extracted for biochemical analysis. An automated biochemical analyzer (Hitachi 7600, Japan) was applied for the measurement of enzymes and compound expressions, including hydroxyproline (HYP), hyaluronic acid (HA), procollagen-III (PC-III), and collagen-IV according to the manufacturer's instruction. Enzyme-linked immunosorbent assay (ELISA) kits were utilized to evaluate the expressions of LPS (YFXER00772; Yifeixue Biotech Co., Ltd., China), diamine oxidase (YFXER00030; Yifeixue Biotech Co., Ltd.), and D-lactate (D-LA) (YFXER00029; Yifeixue Biotech Co., Ltd.) according to the manufacturer's instruction.
Western blot analysis
Levels of Zonula Occludens-1 (ZO-1) and Occludin in intestinal tissues were determined by Western blot. To begin with, total protein was extracted, and its concentration was quantified using the bicinchoninic acid protein quantification method. Monoclonal antibodies against MOUSE Zonula (1:200; Sigma Company, USA) and MOUSE Occludin-1 (1:200; Sigma Company) were used. The protein samples underwent boiling and denaturation, followed by electrophoresis. After electrophoresis, they were transferred to a polyvinylidene fluoride membrane.
Subsequently, the membrane was blocked with a blocking solution at room temperature. The samples were then incubated overnight at 4°C, and the membranes were visualized using chemiluminescent agent enhanced chemiluminescence (Millipore, USA) followed by a gel documentation system (Bio-Rad, USA). ImageJ was utilized for the quantification of gray values of the bands.
16S rRNA gene sequencing
To analyze the fecal microbiota of five randomly selected subjects from each group, 16S rRNA gene sequencing was carried out. E.Z.N.A.® Stool DNA Kit was utilized for the extraction of DNA, and its purity was determined. Polymerase chain reaction (PCR) amplification was carried out using primers 515F 5′-barcode-GTGCCAGCMGCCGCGG-3′ and 907R 5′-CCGTCAATTCMTTTRAGTTT-3′.
Amplicons were obtained and subjected to purification using the AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, USA) per the manufacturer's specifications. After purification, Qubit®3.0 (Life Invitrogen) was utilized for quantifying the PCR products. To enhance diversity, sets of 24 amplicons with distinct barcodes were amalgamated in equal proportions. This pooled DNA served as the basis for constructing a Pair-End library, and the resultant amplicon library underwent paired-end sequencing (2 × 250), with raw reads deposited into the NCBI Sequence Read Archive database.
For the processing of raw fastq files, demultiplexing was performed through barcode sequence information of the sample. The demultiplexing criteria included the following: (i) truncation of 250 bp reads at sites with a mean quality score <20 over a 10 bp sliding window, reads that were <50 bp were removed; (ii) precise matching, and exclusion of reads having unclear characters; (iii) assembly of only sequences with an overlap >10 bp based on overlap sequence, while unassembled reads were abandoned. Operational taxonomic units (OTUs) at a 97% similarity cutoff were clustered through UPARSE 7.1, with the identification and removal of chimeric sequences through UCHIME. The taxonomic affiliation was then determined.
Microbiome analyses
Fecal samples (0.2 g each) were collected and used to analyze the intestinal microbiome. QIAamp DNA Fecal Mini Kit (No. 51504; QIAGEN) was exploited for extracting the DNA of fecal microbial communities, then the sample quality was verified through agarose gel electrophoresis. Primers were designed and used to perform PCR amplification. Sequencing in paired-end mode was performed using the Illumina NovaSeq platform.
Reads obtained were subjected to denoising, merging, and nonchimeric detection using the DADA2 method within QIIME 2 software. The resulting OTU abundance was employed for species-level taxonomy classification, comparing against the Greengenes database. Visual representation of the taxonomic composition for each sample was generated. In addition, Venn diagrams were constructed to illustrate shared or unique OTUs among groups. Clustering results and taxonomic information were further computed and displayed using an interactive heatmap, implemented with the PheatMap package in R.
Metabolomics of rat feces by ultrahigh-performance liquid chromatography with quadrupole-orbitrap mass spectrometry
UPLC-QE-MS is a powerful analytical technique that combines ultrahigh-performance liquid chromatography (UPLC) with quadrupole-orbitrap mass spectrometry (QE-MS). This technology can be used for high-resolution and sensitive analysis of complex samples, including rapid identification and analysis of chemical components. In this study, UPLC-QE-MS (Thermo Fisher Scientific, USA) was utilized for metabolomics of rat feces.
Fecal samples (50 mg each) from the control group, model group, and Cur400 group were combined with 1000 μL of extraction liquid (water: acetonitrile: methanol = 1:2:2). Subsequently, each sample was vortexed for 30 s, ground for 4 min using a grinder, and sonicated for 5 min. These procedures repeated three times. After 1 h of standing at 4°C, the sample was centrifuged (4°C, 1,200 g) for 15 min, the following supernatant was collected into the vial.
Equal amounts of the supernatant from each group were mixed to make the quality control sample. The liquid chromatographic parameters were set as follows: phase A comprised of 25 mmol/L ammonium acetate and 25 mmol/L ammonia, whereas phase B consisted of acetonitrile. A gradient elution scheme was employed with the following time intervals: 0–0.5 min at 95% B, 0.5–7 min transitioning from 95% to 65% B, 7–8 min from 65% to 40% B, 8–9 min at 40% B, 9–9.1 min shifting from 40% to 95% B, and 9.1–12 min at 95% B. Other conditions remained at their default settings.
For mass spectrometry, the sheath gas flow rate was set at 50 Arb, auxiliary gas flow rate at 10 Arb, capillary temperature at 320°C, and scan mode as Full-MS, MS-MS. The spray voltage was adjusted to 3.5 kV (ESI+) or −3.2 kV (ESI–). Raw data were converted to mzXML format, and subsequent processes were conducted through R packages. The substances were then annotated by matching with the secondary mass spectrometry database. The data were standardized by SIMCA software, and metabolites with significant differences were selected based on variable importance in the project >1 and P < .05. Metabolic pathway analysis and correlation analysis of biased metabolites were performed.
Statistical analyses
Statistical analyses were performed on the biological assay results, and the data were expressed as means ± standard deviations. Differences among groups were analyzed using one-way analysis of variance in GraphPad Prism 8.0 (GraphPad Software, USA). Significance was accepted at P < .05.
RESULTS
Curcumin alleviated CCl4-induced hepatic fibrosis
As presumed, CCl4 caused hepatic lipid accumulation, immune cell invasion, and fibrosis, whereas curcumin alleviated liver fibrosis progression in a dose-dependent manner (Fig. 1A). Furthermore, the model group exhibited increased HYP, HA, PC-III, and Collagen IV levels (P < .05; Fig. 1B) compared with the control group; and the curcumin group had lower expressions compared with the model group, which showed antiliver fibrosis effect.

Morphology, histological analysis
Similar results were observed in our supplementary experiment (P < .05; Supplementary Fig. S1). Overall, these findings suggested that curcumin ameliorated hepatic fibrosis in an animal model. The liver fibrosis indicators were also decreased after the administration of EM and Penicillin G (all P < .05; Supplementary Fig. S1).
Curcumin restored the intestinal permeability in rats with liver fibrosis
After the administration of curcumin, there was a notable reduction in epithelial disruption and abscess formation observed in the model group. In addition, curcumin treatment significantly mitigated the heightened edema and cellular infiltration into the submucosal layer (Fig. 2A). Compared with the control group, the model group exhibited higher LPS, DAO as well as D-LA levels, whereas curcumin groups had lower LPS, DAO, and D-LA levels compared with the model group, supporting that curcumin might regulate the intestinal permeability, which was damaged by CCl4 (P < .001; Fig. 2B).

H&E staining of the intestinal
Furthermore, Western blot analyses for tight junction proteins were performed in the intestine of each group. In liver fibrosis rats, the ZO-1 (P < .001) and Occludin levels (P < .001) were decreased significantly, whereas curcumin treatment increased ZO-1 (P < .05) and Occludin levels (P < .05), supporting that curcumin potentially regulated the intestinal permeability in rats with liver fibrosis (Fig. 3). We also discovered that after the administration of EM and Penicillin G, the LPS, DAO, and D-LA levels in liver fibrotic rats were decreased as well (all P < .05; Supplementary Fig. S2).
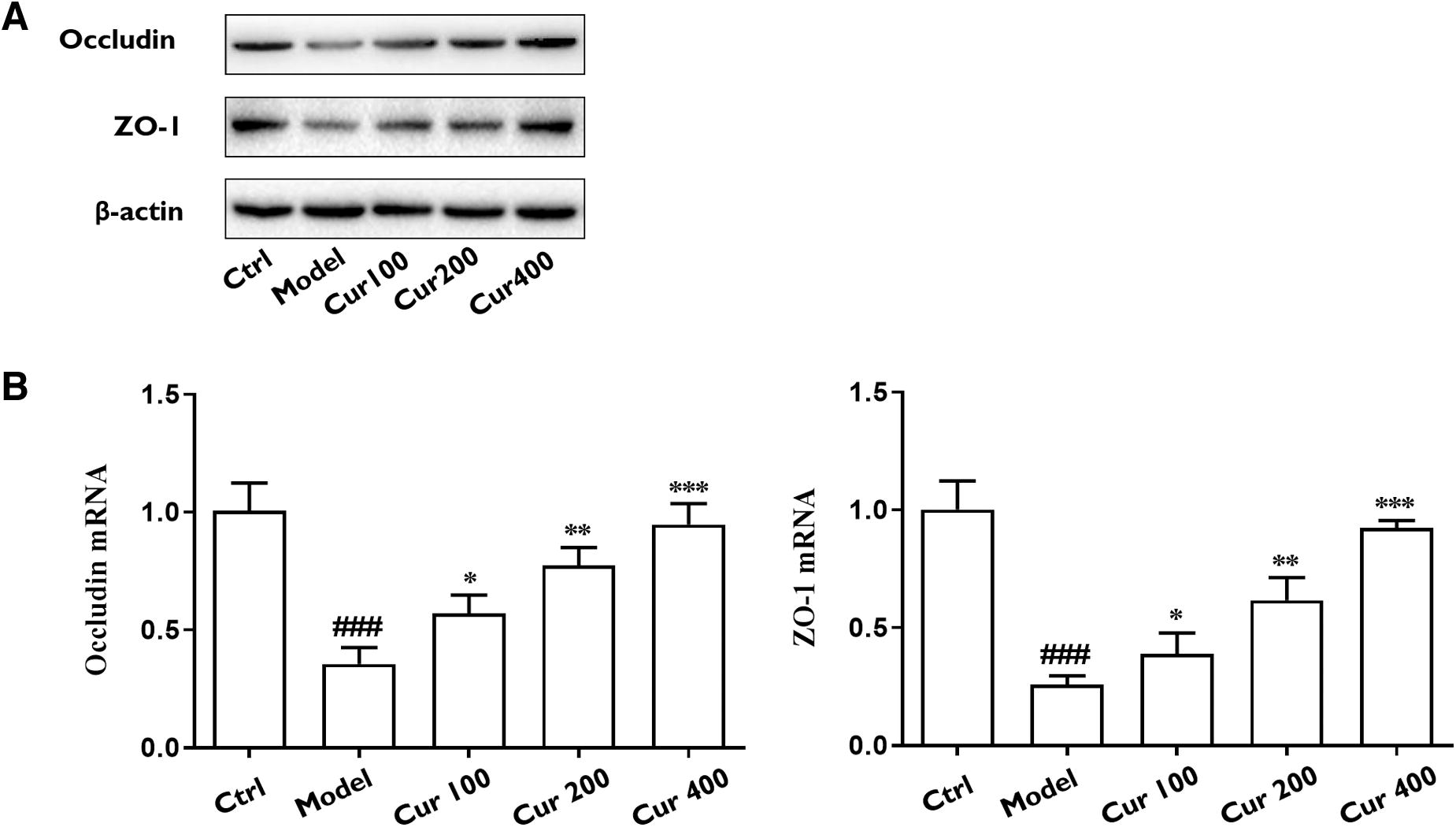
Western blot results of ZO-1 and Occludin
Curcumin alleviated CCl4-induced gut dysbiosis
The higher the Chao 1 index and OTUs number, the higher the flora richness, also, greater Shannon index values and smaller Simpson index values indicated increased flora diversity; see Table 1 and Supplementary Table S1.
Comparison of Diversity Parameters Among Normal Group, Liver Fibrosis Group, and Curcumin Administration Group (
P < .05, compared with control group.
P < .05, compared with model group.
P < .05, compared with Cur100 group.
P < .05, compared with Cur200 group.
Cur, curcumin; OTUs, operational taxonomic units.
The Chao 1 index (P < .05) and Shannon index (P < .05) were reduced in the model group compared with the control group, whereas the Simpson index (P < .05) exhibited an increase relative to the control group, suggesting that the diversity and richness of intestinal flora were compromised in rats with liver fibrosis. Whereas in the curcumin treatment group, the Chao 1 index (P < .05) and Shannon index (P < .05) were increased, and the Simpson index (P < .05) was decreased compared with those of the model group, indicating that curcumin was able to restore the intestinal flora diversity and richness in rats with liver fibrosis.
As shown in Figure 4, there were 746 OTU sequences in the liver fibrosis model group, control group, and curcumin administration groups, 12 unique sequences in the normal control group, 10 in the model group, and 404, 32, and 18 in low-, middle-, and high-dose curcumin administration groups, respectively. Principal component analysis results are shown in Figure 5; the samples of the model group and normal control group were clustered without crossing, which was well differentiated, and different individuals in the model group had a large degree of dispersion, which indicated that there existed a huge difference in the flora structure, while there was a small individual difference in the flora structure in the control group. In addition, the high, middle, and low doses of curcumin could adjust for the composition of the flora to different degrees, among which the low dose of curcumin had the most significant effect.
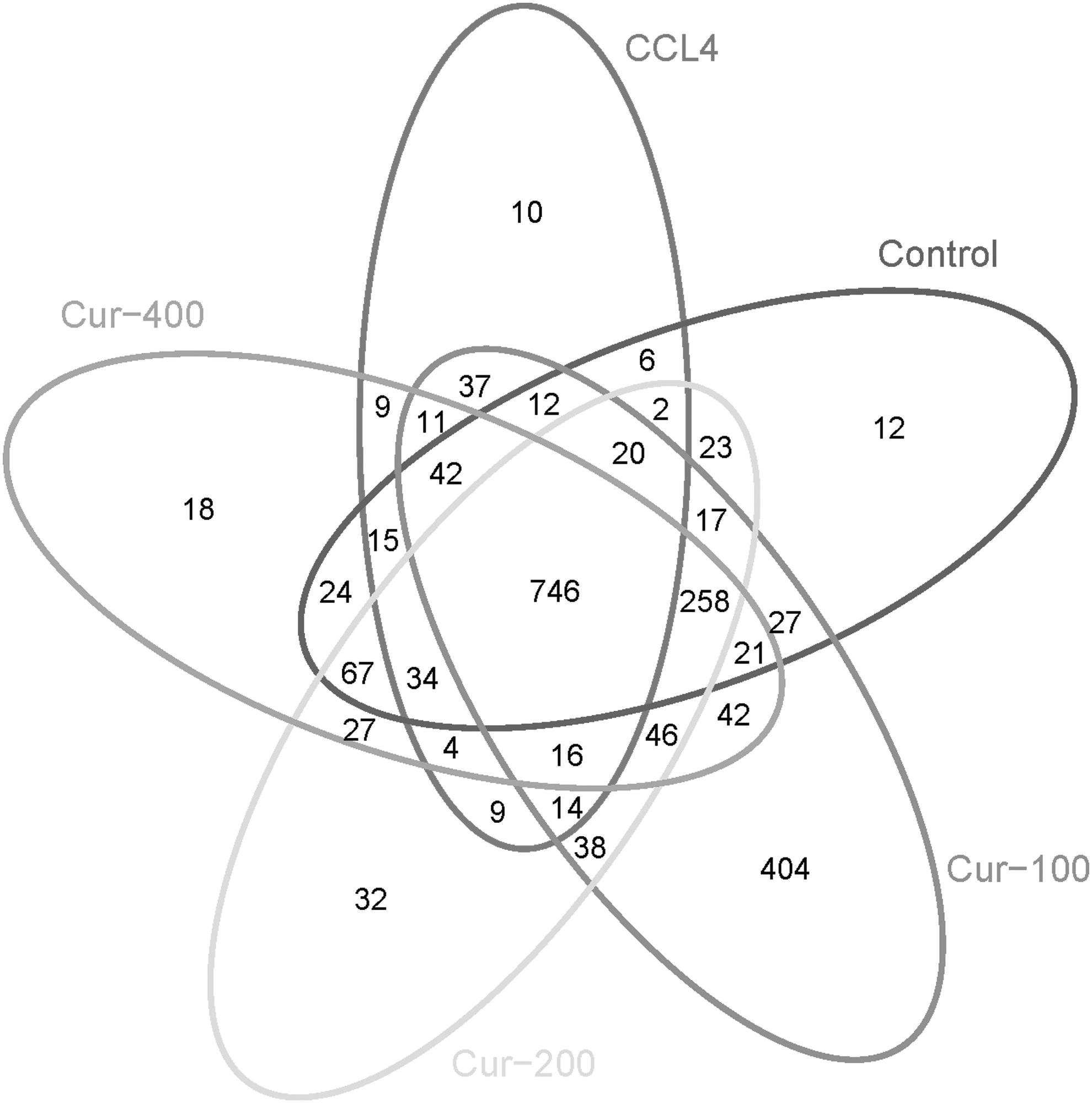
Venn diagram of the OUT distribution of each group. CCL4, carbon tetrachloride; OUT, operational taxonomic units.
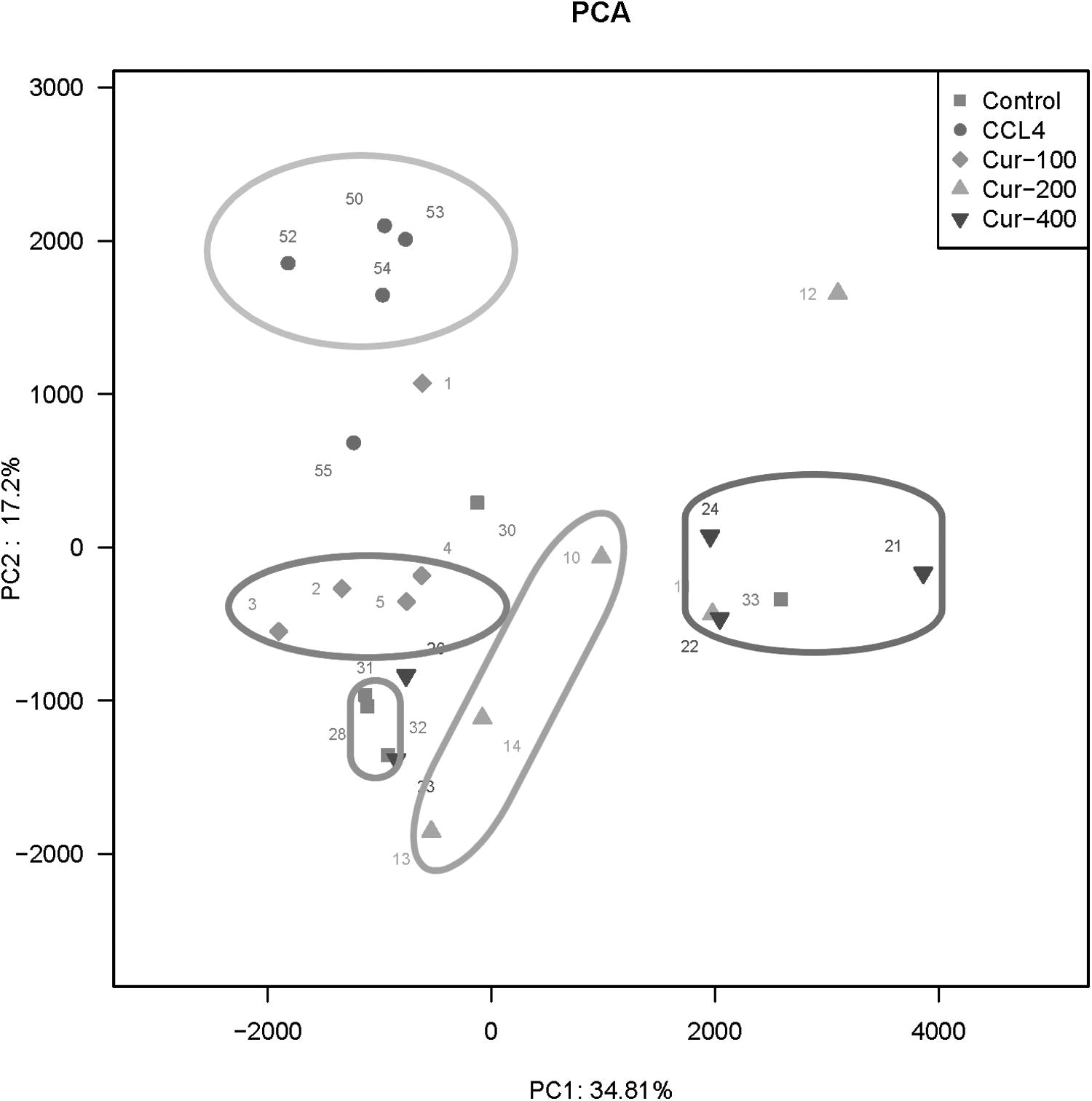
PCA scatter diagram of each group of samples. PCA, principal component analysis.
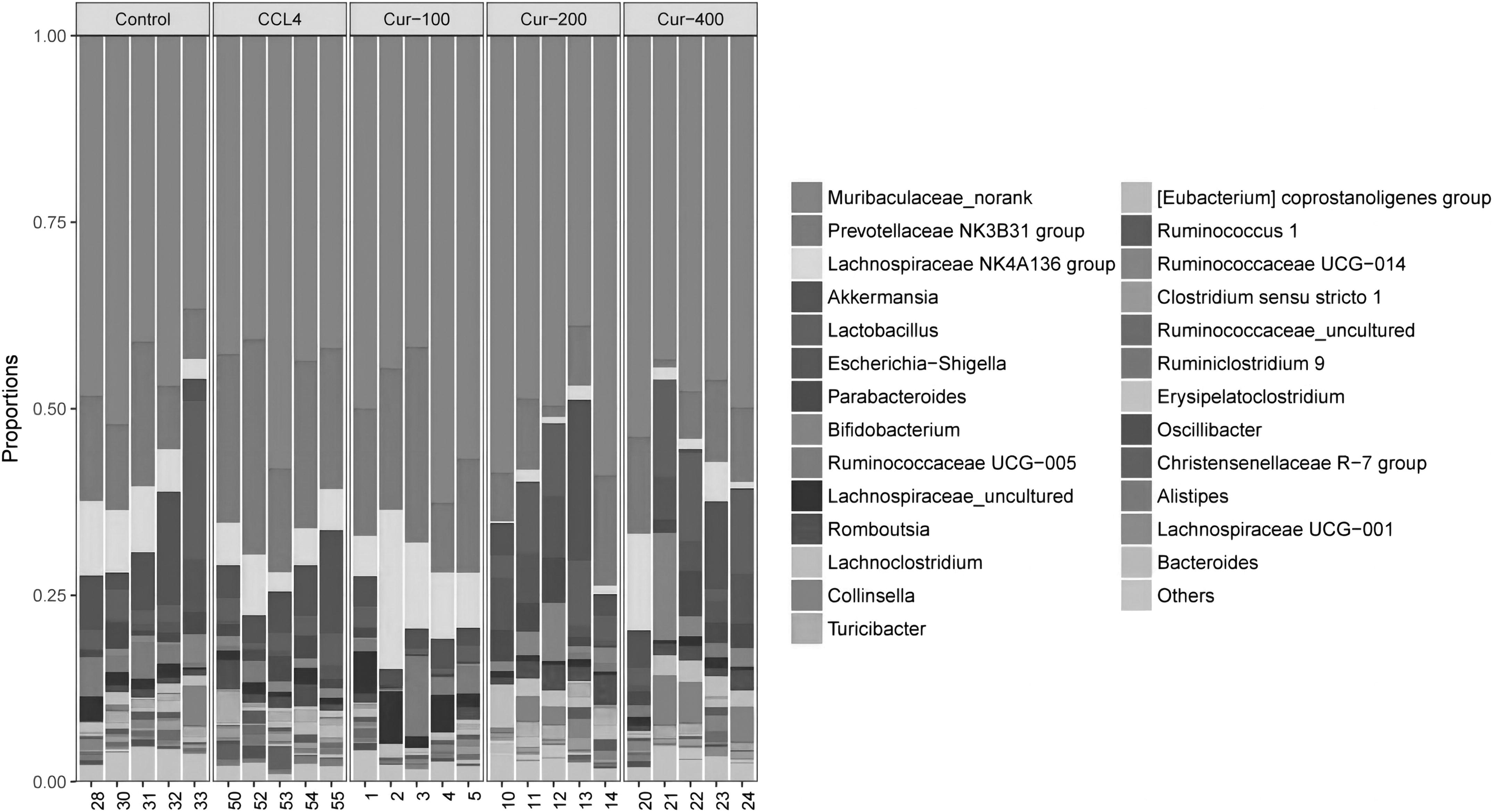
Figure 6A shows that at the phylum level, Proteobacteria accounted for 2.07% in the control group, 0.69% in the model group, and 5.23% and 4.29% in the middle- and high-dose groups of curcumin, respectively, which suggested that proteobacteria could be increased after the middle and high dose of curcumin. Figure 6B shows that lactobacillus represents 6.41% in the control group, 2.62% in the liver fibrosis model group, 6.85% in the curcumin middle-dose group, and 8.01% in the curcumin high-dose group. It suggested that curcumin could increase the proportion of lactobacillus in the intestinal tract of rats with hepatic fibrosis.
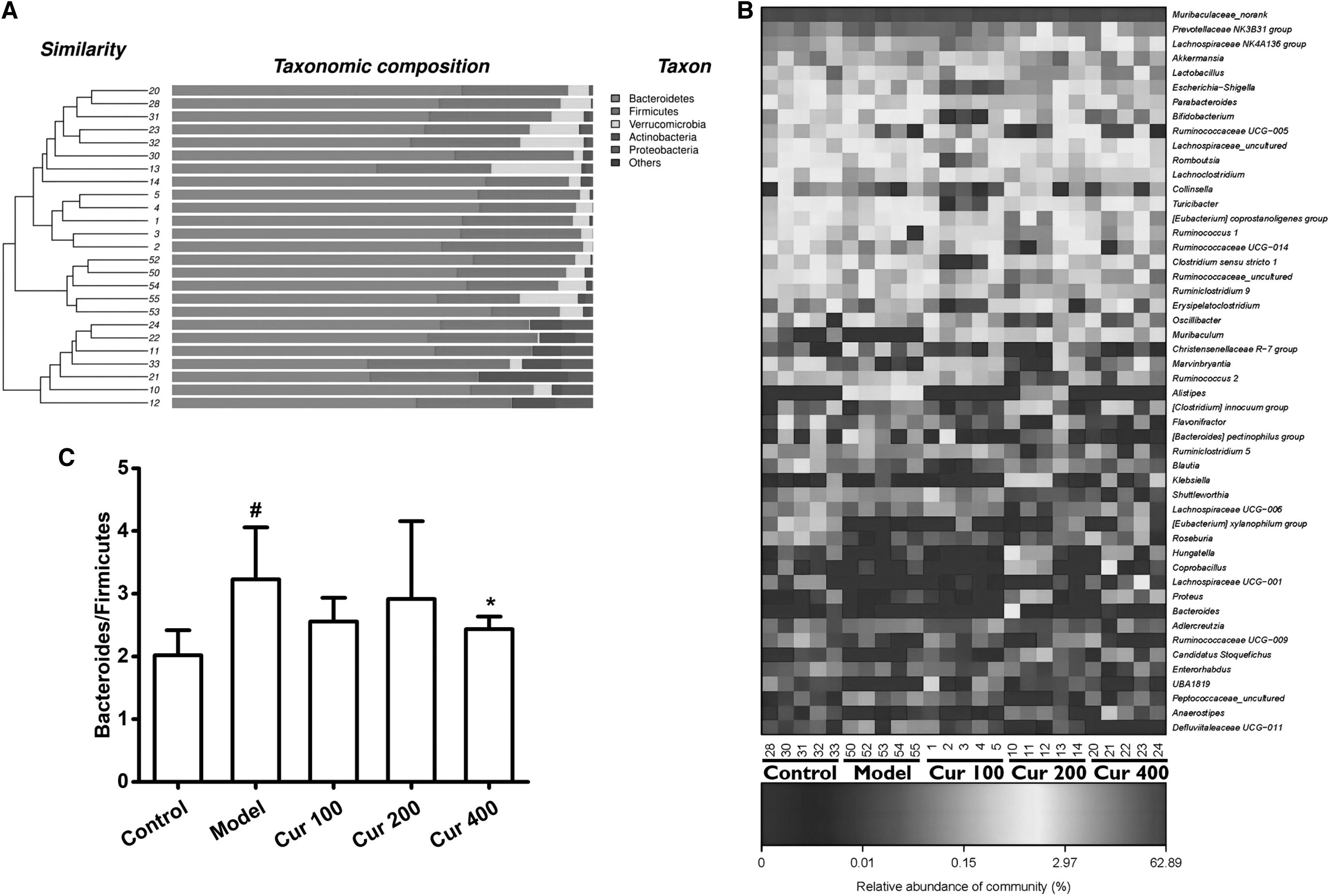
The cluster of intestinal microorganisms
Figure 6C shows that the model group had higher ratio of bacteroides/firmicutes compared with the control group (P < .05), and curcumin could reduce the increase of bacteroides/firmicutes ratio in different degrees when liver fibrosis occurs.
Curcumin regulated intestinal microorganisms
From Figure 7, it could be seen that there were different microorganisms with linear discriminant analysis (LDA) scores greater than the set value (set value = 3), in other words, biomarkers with statistical differences. In the Lefse analysis, Firmicutes (firmicutes) were enriched in the control group; Prevotellaceae (Prevost bacteriaceae), Defluviitaleaceae, Peptococcaceae (digestive coccidae) were enriched in the liver fibrosis model group; Clostridiales and Clostridia were enriched in curcumin low-dose group; Proteobacteria (Proteobacteria), Peptostreptococcaceae (Gastroenterobacteriaceae), Enterobacteriaceae (Enterobacteriaceae) were enriched in curcumin in the middle-dose group; Actinobacteria (Actinomycete), Bifidobacterium (Bifidobacterium), and Eubacteriaceae (Eubacteriaceae) were enriched in curcumin high-dose group.
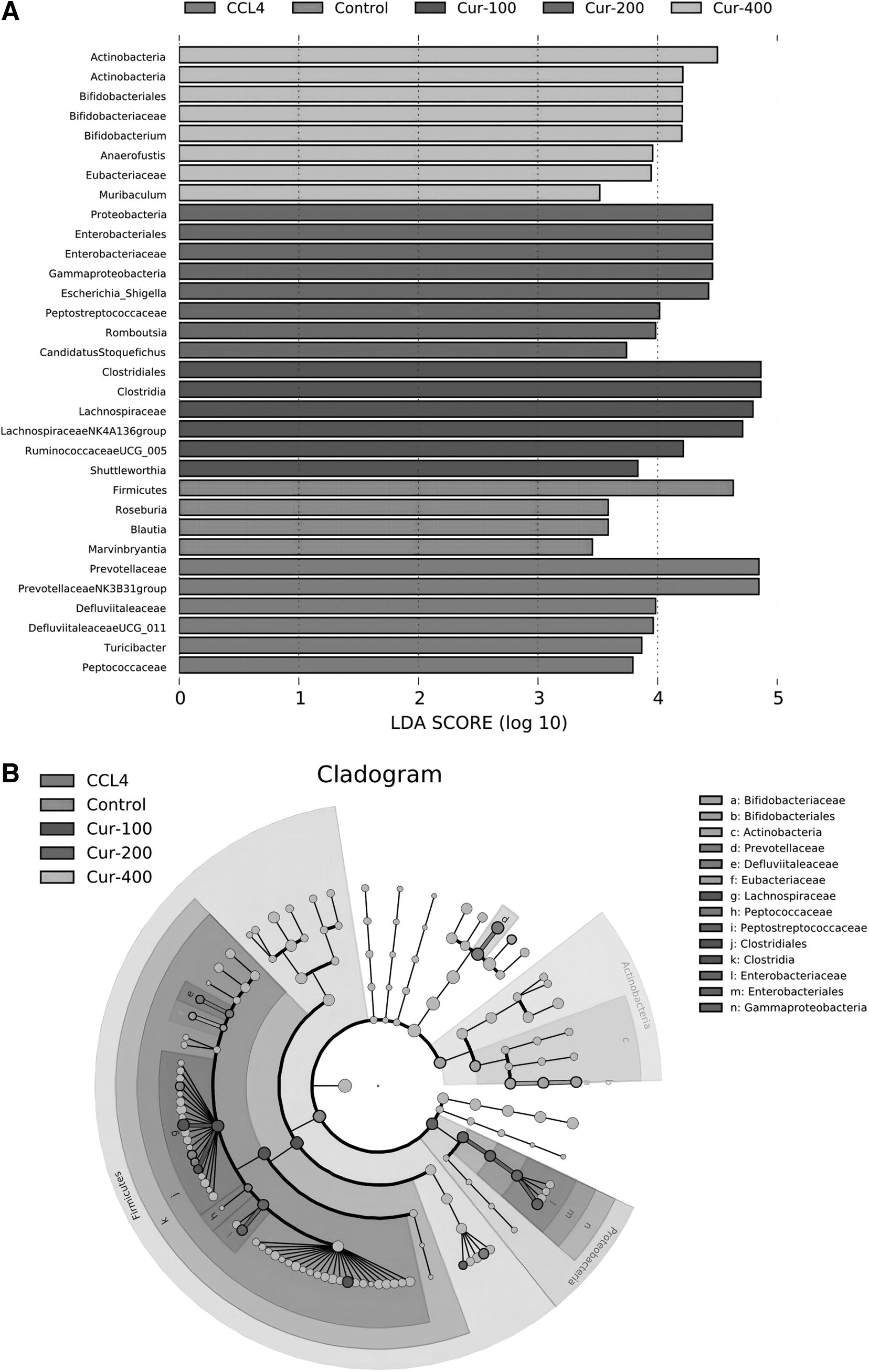
LEfSe analysis of intestinal flora data.
The left histogram illustrates the LDA score computation for the feature at the OTU level (Fig. 7A), whereas the right cladogram depicts the relative abundance of OTUs (Fig. 7B). In the LEFse tree, distinct colors signified different groups, with each color indicating a significant microbial marker in that group. The corresponding biomarker names are listed in the upper right corner.
The effect of curcumin on intestinal flora functional genes
The function of intestinal flora was predicted by PICRUSt analysis. According to the 16S sequencing data, the function prediction based on the KEGG database was carried out. The functional composition and relative abundance information of the sample could be visually displayed by the histogram of Figure 8. In the functional gene module, LEfSe was used to compare the functional genes with significant differences among different groups.
The functional composition and relative abundance information of the sample displayed by the histogram.
As shown in Figure 9, the functional genes of replication, recombination, and repair proteins were enriched in the control group; functional genes such as those of amino acid metabolism, polysaccharide biosynthesis and metabolism, energy metabolism, cofactor and vitamin metabolism, cell process and signal, folding sorting and degradation, oxidative phosphorylation, metabolism, gene information process, membrane and intracellular structural molecules, carbon fixation pathway in prokaryotes, terpenoids and polyketides metabolism, transcription, energy metabolism, secondary metabolites biosynthesis, citric acid cycle, etc. were enriched in the model group.
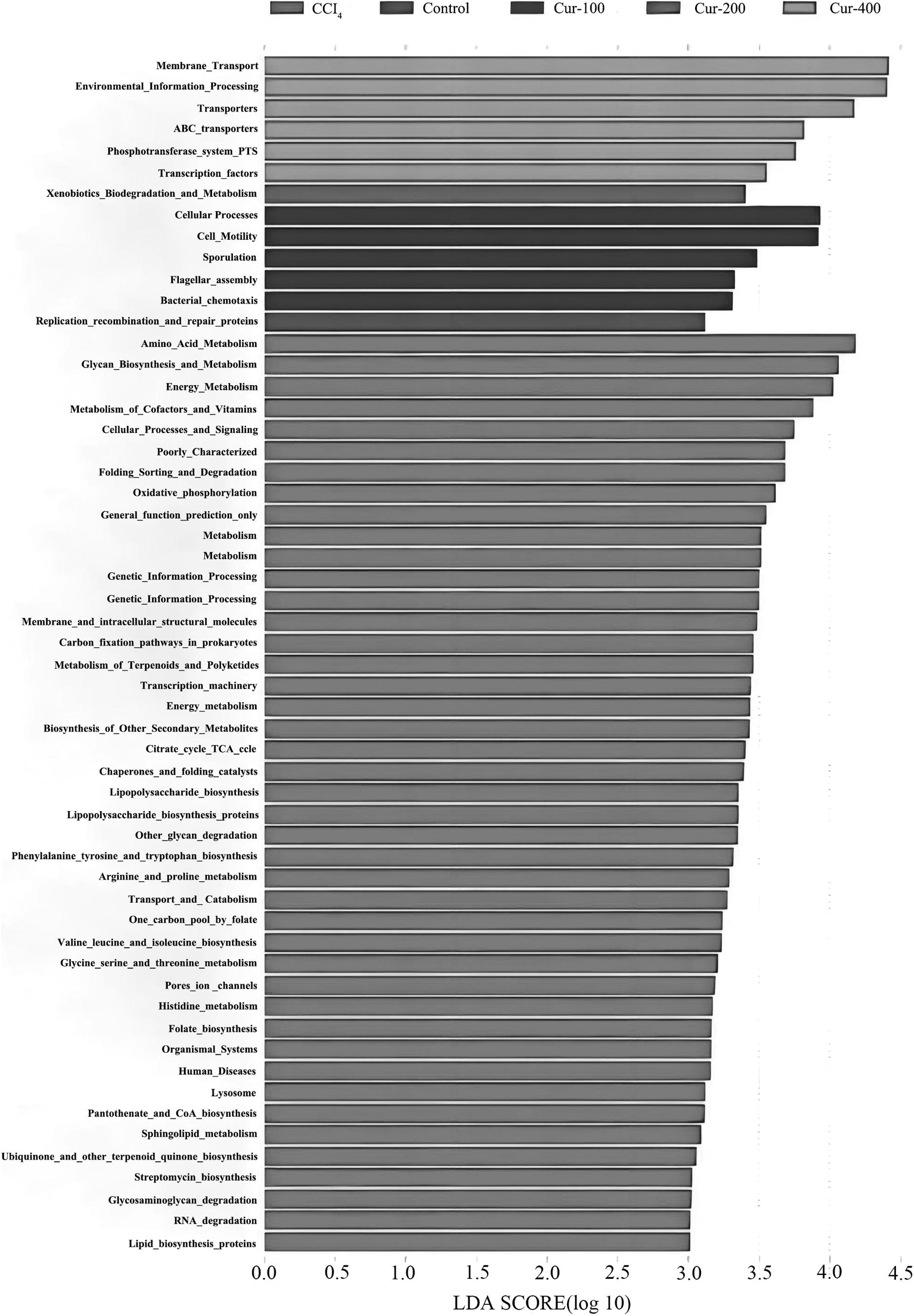
Comparison of the functional genes with significant differences among different groups.
The functional genes involved in cell processes, cell movement, spore formation, flagella combination, and cell chemotaxis were enriched in the curcumin low-dose group. Exogenous biodegradation and metabolic function genes were enriched in the curcumin middle-dose group; membrane transport, environmental information processing, transport, phosphotransferase system, and transcription factor functional genes were enriched in the curcumin high-dose group.
The effect of curcumin on metabolomics
Utilizing the UPLC-QE-MS metabolomics approach, we conducted a comprehensive analysis of the metabolites present in rat feces across the control group, model group, and Cur400 group. One thousand eighty-nine metabolites were annotated in positive mode, and 380 metabolites were annotated in negative mode, among which 16 positive metabolites and 8 negative metabolites were differentially expressed among the 3 groups. The detailed information of these 24 differential metabolites is listed in Tables 2 and 3.
Identification of Differential Metabolites Among Control Group, Model Group, and Cur400 Group in Positive Mode
Identification of Differential Metabolites Among Control Group, Model Group, and Cur400 Group in Negative Mode
Metaboanalyst was further utilized to investigate the signaling pathways of the 24 differential metabolites, which revealed that the biased metabolites were enriched in glycerophospholipid metabolism, pantothenate and CoA biosynthesis, sphingolipid metabolism, glycosylphosphatidylinositol anchor biosynthesis, glycerolipid metabolism, beta-alanine metabolism, propanoate metabolism, phosphatidylinositol signaling system, pyrimidine metabolism, and tryptophan metabolism (Table 4). The results revealed that curcumin might regulate the gut metabolites and these signaling pathways, thereby exerting an antiliver fibrosis effect.
Metabolic Pathway of Differential Metabolites
DISCUSSION
In our study, we report the discovery that curcumin has the ability to alleviate CCl4-induced liver lesions and fibrosis. Moreover, we found that CCl4-induced liver fibrosis results in damage to intestinal permeability, gut dysbiosis, and intestinal microorganisms alterations, which curcumin is able to repair. In addition, we analyzed the functional genes of the intestinal flora, and differential metabolites of rat feces as well as their signaling pathways. Our study offers fresh perspectives on the potential of curcumin in treating liver fibrosis through its regulation of the gut–liver axis. This contributes to an enhanced comprehension of the interplay between curcumin and the gut–liver axis, presenting a novel viewpoint on the treatment of liver fibrosis.
Hepatic fibrosis refers to a number of pathological processes commonly associated with different chronic liver diseases. Reversing hepatic fibrosis effectively can prevent chronic liver diseases from progressing into cirrhosis and liver cancer. 19 –21
Curcumin, a polyphenol or diketone compound extracted from the rhizomes of plants in Zingiberaceae and Araceae, has been shown to exhibit liver-protective effect. Various studies have demonstrated its antiliver fibrosis effect in recent years. 22 –24 In line with these studies, we discovered that curcumin can decrease CCl4-induced liver fibrosis indicators such as HYP, HA, PC-III, and Collagen IV, indicating its potent antiliver fibrosis activity.
To uncover the detailed mechanism of the antiliver fibrosis effect of curcumin, great efforts have been made. For example, a previous study discovered that curcumin prevented liver fibrosis by inducing apoptosis and suppressing the activation of hepatic stellate cells. 25 In another study, curcumin was found to modulate cannabinoid receptors in liver fibrosis in vivo. 26 Besides, the antiliver fibrosis effect of curcumin was reported to be associated with TGF-β/Smad3 and NF-κB signaling. 27 However, the detailed mechanism still needs additional exploration.
The gut–liver axis is crucial for metabolic health, and its role in liver diseases has been increasingly recognized in recent years. 28 –31 Our study hypothesized that the antiliver fibrosis effect of curcumin may be related to the gut–liver axis, considering its importance in liver diseases. However, the specific mechanism remains unclear. Thus, we analyzed changes in intestinal permeability and intestinal flora in rats with liver fibrosis through intestinal permeability indicators (LPS, DAO, D-LA), Chao 1 index, Shannon index, and Simpson index, respectively. We found that curcumin repaired the intestinal permeability in rats with CCl4-induced liver fibrosis, and increased the abundance and diversity of intestinal flora.
In addition, curcumin corrected the imbalance in the flora composition of liver in rats with fibrosis and brought it closer to that of normal rats. Studies have shown that Bifidobacterium can improve liver injury by reducing the expression of inflammatory factors, increasing the intestinal barrier, and producing short-chain fatty acids. 32 –34 We believe that curcumin can play an antifibrosis role by balancing the ratio of Bifidobacterium/Firmicutes. Our results suggest that curcumin regulates intestinal permeability, improves intestinal microflora disorder caused by liver fibrosis, and upregulates the proliferation of beneficial bacteria, thereby exerting antiliver fibrosis effects through the gut–liver axis.
Intestinal flora functional genes and differential metabolites of feces were further analyzed, the results showed that cell process, cell movement, spore formation, flagella combination, cell chemotaxis, exogenous biodegradation and metabolism, membrane transport, environmental information processing, phosphotransferase system, and transcription factor functional genes were enriched in curcumin groups, indicating that curcumin might regulate these intestinal flora functional genes.
In addition, 24 differential metabolites were annotated, and these differential metabolites were mainly enriched in several signaling pathways, including glycerophospholipid metabolism and pantothenate and CoA biosynthesis. Glycerophospholipid metabolism is critical for normal gut functions, and recent studies suggested that glycerophospholipid metabolism is varied in inflammatory bowel disease.
Pantothenate is a fundamental element for human beings, which plays a key role not only in fatty acid synthesis but also in gut normal functioning. Considering our result that the biased metabolites were primarily enriched in glycerophospholipid metabolism and pantothenate and CoA biosynthesis, curcumin may regulate the glycerophospholipid metabolism and pantothenate biosynthesis in the gut, thereby exerting antiliver fibrosis effects through gut–liver axis. The detailed mechanism needs further investigation.
Although several studies already have reported the hepatoprotective effects of curcumin in animal model, 35,36 Feng et al. reported that curcumin alleviates high-fat diet–induced hepatic steatosis in an animal model through enhancing the function of intestinal barrier and diminishing endotoxin as well as liver inflammation. 37 However, most of them were focused on hepatic steatosis rather than liver fibrosis. The remaining studies that investigated the protective effect of curcumin against liver fibrosis did not explore the in-depth correlation with the gut–liver axis. Besides, seldom studies investigated the effect of curcumin with different concentrations. Therefore, our study has unique value and novelty.
Histopathology remains the gold standard for diagnosing liver fibrosis, as inspection items cannot be replaced. H&E staining, Masson's trichrome staining, and Sirius Red staining are commonly used to evaluate tissue injury, steatosis, and collagen deposition, which are important indicators in the process of liver fibrosis. The accumulation of collagen fibers and damage to normal liver cells are characteristic of liver fibrosis, and histopathological examination can clarify the degree of liver injury and fibrosis. However, due to the difficulty in obtaining pathological tissues, blood test indexes such as ALT, AST, ALB, and LDH are also commonly used to evaluate liver injury and fibrosis. 38 Four tests of liver fibrosis, including HYP, HA, PC-III, and Collagen IV, are important in the clinical diagnosis of liver inflammatory activity and degree of fibrosis. 39
HYP is a major component of collagen tissue, and its detection in liver tissue can evaluate the degree of liver fibrosis. HA can reflect the amount of fiber generated in the liver and the damage to hepatocytes. PC-III is a marker of ongoing collagen synthesis and has been shown to increase in the blood of patients with liver fibrosis. Collagen IV is a major component of the basement membrane, and is involved in the development and progression of liver fibrosis. These biomarkers could be useful in diagnosing liver fibrosis.
In summary, curcumin was found to reduce liver fibrosis and potentially play an important role in the gut–liver axis functioning, which paves the way for a deeper understanding of the mechanism of the gut–liver axis, as well as guiding future treatment strategies for liver fibrosis.
Footnotes
ACKNOWLEDGMENT
The authors acknowledge Gu Xiaosong's academician workstation.
AUTHOR CONTRIBUTIONS
Study conception and design: D.K. and H.H.; Material preparation and data collection: Q.G. and Y.X.; Data analysis: Y.H. and H.H.; Writing- original draft preparation: Q.G. and Y.X.; Writing - review and editing: Y.X., W.H., and H.J.; Supervision: D.K.
DATA AVAILABILITY STATEMENT
The data presented in this study are available upon reasonable request from the corresponding author. The data are not publicly available due to privacy.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This study was supported by Jiangsu Province Traditional Chinese Medicine Science and Technology Development Plan Project (Grant No. QN202112).
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
Supplementary Table S1