
Abstract
Tao-Hong-Si-Wu-Tang (THSWT), a traditional Chinese herbal remedy, is commonly utilized for the treatment of female perimenopausal depression through regulating menstruation, but the mechanism remains unknown. In this study, ICR mice were randomly divided into six groups: low, medium, and high dose of THSWT (0.5, 1.5, and 4.5 g/kg), soy isoflavone (250 mg/kg), ovariectomy group, and control group. All mice, except the control group, had ovaries removed and were exposed to hypoxic stimulation for 28 days to establish a perimenopausal depression mice model. The mice, having unrestricted access to food and water, were administered THSWT treatment for a duration of 14 days. The Western blotting and Enzyme linked immunosorbent assay kits were used to determine protein and hormone levels, respectively. Experimental results showed that THSWT reduced the immobility time of mice from 150.8 s to 104.9 s in the tail suspension test, and it decreased the immobility time of mice from 165.7 s to 119.0 s in the forced swimming test, outperforming the results obtained with soy isoflavones. In addition, THSWT upregulated the protein expression of follicle-stimulating hormone receptor and downregulated the protein expression of corticotropin-releasing hormone-receptor 1 in the hippocampus. Compared with the oophorectomized group, treatment with THSWT decreased the levels of corticosterone and adrenocorticotropic hormone in serum by 173.7 and 23.4 ng/mL, respectively. These findings showed that THSWT could stimulate the perimenopausal nerve tissue and regulate the level of serum hormones in mice. THSWT exhibited promising potential as a viable alternative drug for hormone treatment of perimenopause in clinical use.
INTRODUCTION
Menopausal depression refers to a group of autonomic nervous system disorders in women caused by hormonal fluctuations. 1 Its main pathogenesis is ovarian dysfunction, which is caused by a great decrease in estrogen secretion because of hypothalamic–pituitary–ovarian axis dysfunction. 2 Menopausal depression is often characterized by lethargy, black mood, slow thinking, pessimism, and irritability. 3 Such pathological behaviors are primarily caused by the reduction of body’s ovarian hormone secretion. 4 As a result, women are vulnerable to external factors during menopause, which can lead to the development of menopausal depression. Menopause typically occurs in middle age, which is also the most common period for obstructive sleep apnea (OSA). OSA is a sleep-related respiratory disorder that causes respiratory arrest because of upper respiratory tract obstruction during sleep. 5 –7 Reduced oxygen saturation and disturbed sleep structure may contribute to neurocognitive dysfunction and mood disorders in patients with OSA. 8–9 Cognitive impairment caused by OSA injury is mainly characterized by reduced attention and alertness, learning and memory ability, and execution timeliness. 10 Some studies have shown that cognitive hypoxia can cause damage to specific brain regions from a pathophysiological standpoint. 11 Cognitive impairment, on the contrary, may be a consequence of OSA because of sleep deprivation. 12 The severity of OSA mood disorders is usually described as a symptom of depression and anxiety. 13 Overall, OSA in midlife is more likely to lead to anxiety or depression in postmenopausal women, which is also known as menopause.
Hypothalamic–pituitary–adrenal (HPA) axis hyperactivity is the most common neuroendocrine abnormality in menopausal depression. 14 It is characterized by elevated serum glucocorticoid, adrenocorticotropic hormone (ACTH), and corticotropin-releasing hormone (CRH). 15 The HPA axis is an important neuroendocrine network that controls hormone responses to internal and external stress, which shows significant gender-biased activity in humans and animals. 16 The HPA axis is more significantly regulated in women compared with men in response to abnormal chronic stress, such as significantly higher levels of corticosterone (CORT) and ACTH, 17 and this difference is largely due to changes in gonadal hormones. 18 Gonadal hormones interact with central and peripheral components of the HPA axis and regulate the synthesis and release of CRH, ACTH, and CORT. 19 Conversely, overactivation of the HPA axis also impairs reproductive function and hypothalamic–pituitary–gonadal (HPG) axis at all levels, including excessive release of follicle-stimulating hormone (FSH) and luteinizing hormone (LH). 20
Neurotrophic changes are closely associated with anxiety, depression, and cognitive deterioration associated with menopausal depression.
21
Brain-derived neurotrophic factor (BDNF), a highly prevalent neurotrophic factor in the mature brain, possesses significant importance in the survival, preservation, and regeneration of specific neurons, and is intricately linked with neurological disorders. BDNF is also highly expressed in the female reproductive system and promotes uterine growth and proliferation.
22
Previous experiments have shown that estradiol (E2) can increase the protein expression of BDNF in the prefrontal cortex, hippocampus, and uterus.
23
Tropomyosin-related kinase B (TrkB) also plays a key role in the pathogenesis of depression. Furthermore, it is important for neuron maturation, synaptic formation, and synaptic plasticity.
24
Cyclic adenosine monophosphate (cAMP) response element-binding protein (CREB) is another important transcription factor in the brain that controls biosynthesis of BDNF.
25
It has also been suggested that binding of BDNF to TrkB protein receptors activates two key downstream target signaling pathways. The first pathway induces CREB phosphorylation and activates mitogen-activated protein kinase (MAPK)/extracellular regulatory protein kinase (ERK) signaling pathway. The second pathway activates phosphatidylinositol 3-kinase (PI3K)/protein kinase B (PKB/AKT) signaling.
26–27
A lack of BDNF
Today, hormone replacement therapy (HRT) is considered as a mainstream treatment option, but much evidence suggests that women taking HRT have a 2.3% increased risk of breast cancer per year (1.1
Traditional Chinese herbs have been utilized for centuries in China for the treatment of various ailments. Contemporary medical research has provided evidence in support of the effectiveness of traditional Chinese herbs and extracts in addressing menopausal syndrome. Tao-Hong-Si-Wu-Tang (THSWT), which is composed of Prunus davidiana (CarriŠre) Franch. (Rosaceae), Carthamus tinctorius L. (Compositae), Radix Angelica sinensis (Oliv.) Diels (Apiaceae), Radix Rehmannia glutinosa (Gaertn.) DC. (Scrophulariaceae), Radix Paeonia lactiflora Pall. (Ranunculaceae), and Radix Ligusticum wallichii Franch. (Apiaceae), can promote blood circulation and remove blood clots. The whole formula is effective in blood nourishing, vasodilation, reducing hypoxia, and so on. THSWT has been shown to affect the growth and development of brain tissue through PI3K/AKT and Nrf2 signaling pathways. 32 Another study also showed that THSWT alleviates cerebral arterial embolism. 33
MATERIALS AND METHODS
Drugs
The following herbs for THSWT were purchased from Anhui Zhiyuan Traditional Chinese Medicine Co. Ltd. (Bozhou, China) on May 30, 2019: P. davidiana (Origin: Sichuan, China; Batch number: 180301), C. tinctorius (Origin: Shanxi, China; Batch number: 180302), A. sinensis (Origin: Guangdong, China; Batch number: 180401), R. glutinosa (Origin: Gansu, China; Batch number: 180402), P. lactiflora (Origin: Sichaun, China; Batch number: 180407), L. wallichii (Origin: Hebei, China; Batch number: 180306). Soy isoflavone (SI) was purchased from Beijing Solarbio Science and Technology Co., Ltd. (Beijing, China).
Preparation of THSWT
The herbs were authenticated by Prof. Wei Wang from Zhejiang Provincial People’s Hospital, China. The voucher specimens (No.2018021
Animal modeling and treatment
Sixty 10-week-old female ICR mice were selected and anesthetized with 2% ether after 1 week of adaptation to the experimental environment. Ten mice were randomly assigned to the control group and underwent a sham operation (surgery without ovary removal), whereas the remaining animals underwent bilateral ovariectomy (OVX) to establish a menopause model. Mice were given a 5-day recovery period, and vaginal smears were performed for 5 consecutive days to confirm complete ovarian resection. The mice were then randomly divided into 5 groups (n = 10 per group): model group, low, medium, and high dose of THSWT groups (0.5, 1.5, and 4.5 g/kg; crude drugs), and SI group (250 mg/kg). After recovery period, all animals were treated with OSA for 28 days, except for the control group. The OSA model device is 40 × 30 × 15 cm plexiglass transparent container with a vent connected to a circulating on/off control timer valve. Pure nitrogen was introduced into the chamber for 40 sec to reduce the oxygen level to 5–7% for 10 sec. Pure oxygen was then introduced for 10 sec to gradually increase the oxygen level in the chamber to 20.9%. The gas input was circulated once per min for 8 h every day (9:00 a.m.–5:00 p.m.). After 4 weeks of OSA treatment, the mice in the drug-treated groups were administered intragastrically (i.g.) with THSWT (0.5, 1.5, and 4.5 g/kg) or SI (250 mg/kg) daily for 14 days (all drugs were dissolved with saline solution and then were given to mice through intragastric administration). The schedule is shown in Figure 1. All procedures in this study were performed following the principles of Helsinki and were approved by the Ethics Committee and the Animal Experimental Committee of Zhejiang Provincial People’s Hospital (Reference No. SYXK20190013).
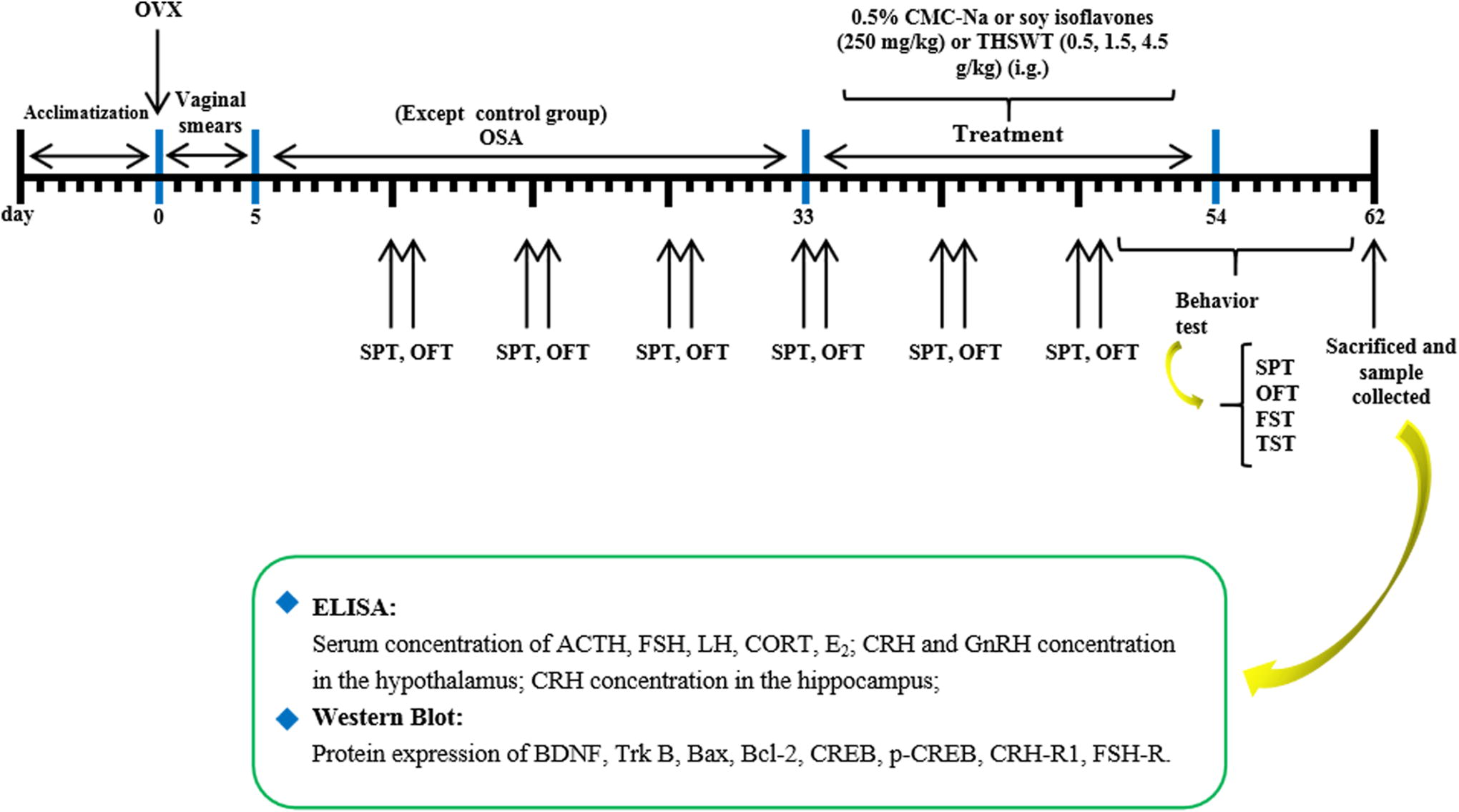
The schedule of the experimental design. FST, forced swimming test; OFT, open field test; SPT, sucrose preference test; TST, tail suspension test. The mice were divided into six groups: the sham-operated group (control group); the ovariotomy + OSA group (OVX-OSA); the low, medium, and high dose of THSWT groups (0.5, 1.5, and 4.5 g/kg); soy isoflavones group (250 mg/kg/day); adrenocorticotropic hormone (ACTH); follicle-stimulating hormone (FSH); luteinizing hormone (LH); corticosterone (CORT); estradiol (E2); corticotropin-releasing hormone (CRH); corticotropin-releasing hormone receptor 1 (CRH-R1); gonadotropin-releasing hormone (GnRH); FSH receptor (FSH-R). OSA, obstructive sleep apnea; OVX, ovariectomy; THSWT, Tao-Hong-Si-Wu-Tang.
Body weight
Body weight was recorded at the beginning of the OSA procedure and then weekly until the end of the study.
Behavior tests
Behavior tests were carried out once a week during the experiment. All tests were conducted in a soundproof environment from 8:30 to 14:30.
The sucrose preference test (SPT) involved depriving mice of food and water for 12 h and providing two bottles of water and allowing the mice to drink freely for 4 h (one containing 1% sucrose solution and the other containing pure water). The bottles were switched at any time during the test to prevent confounding the animals’ drinking behaviors. The consumption of water and sucrose solution and total fluid intake were recorded by weighing the bottle. The sucrose preference index was calculated as the ratio of consumption sucrose solution to total fluid intake.
The open field test (OFT) is a behavior test that measures both motor exploration ability and depression levels. The apparatus consisted of a black box (100 cm long × 100 cm × 50 cm high). The floor is divided into 25 equal squares. The animals were placed in the corner of the apparatus backward to the wall. The times of rearing, grooming, and defecation and standing in the center over a period of 5 min was recorded and observed by the person who was blind to the treatment.
The forced swim test (FST) involves placing animals in a transparent plexiglass container with dimensions of 60 cm in height and 25 cm in inner diameter. The water temperature is maintained at 24 ± 2°C, and the water depth is set at 40 cm. The total immobility time of the animals was recorded over the next 6 mins. When an animal did not attempt to escape (except for the necessary movements such as keeping its head above the water for breathing), it could generally be considered to be stationary. The immobility time was recorded by the researcher blind to the treatment.
Suspended tail test (TST) involves fixing the animal with adhesive tape 1/3 of the way from the tail tip and suspended it upside down on the fixing bracket with the head 15 cm above the surface of table. The total immobility time of the animals was recorded over the next 6 min. The animal was considered immobile when it was suspended passively, showing no resistance to the stress, and the immobility time was recorded by the researcher blind to the treatment.
Serum and tissue sample collection
The mice were sacrificed under deep anesthesia 24 h after the behavior test. Blood was collected from the abdominal aorta and centrifuged (6.2 g at 4°C for 15 min). The brain was quickly extracted from the head in an ice bath, and subsequently, the hippocampus and hypothalamus were rapidly dissected in the ice bath, and then cryopreserved with liquid nitrogen. The pituitary gland was also excised and quickly stored in liquid nitrogen. The uterus was extracted by ice bath and weighed. The uterus index was calculated as follows: uterus index = wet weight of uterus (mg)/mouse weight (g). All samples were frozen with liquid nitrogen.
Histological examination of uterus
After calculating the uterine index, the uteri in the mice were washed with 0.15M saline, fixed with 4% paraformaldehyde, dehydrated with alcohol, embedded in paraffin at 4°C overnight, and cut into 4-μm-thick sections. Uterine tissue was examined histologically by hematoxylin–eosin staining (HE) of paraffin-embedded sections and examined under a microscope at ×400 magnification (Olympus IX73, Japan).
ELISA measurement of E2, FSH, LH, ACTH, CORT, and 5-HT in serum, gonadotropin-releasing hormone, and CRH in hypothalamus
On the day following the behavior test, the mice were deeply anesthetized using ether. Enzyme linked immunosorbent assay (ELISA) kits were used to measure the concentrations of gonadotropin-releasing hormone (GnRH) and CRH in the hypothalamus. In addition, the levels of E2, ACTH, FSH, CORT, LH, and CRH in the serum were also determined using the ELISA kits from Jianglai (Shanghai, China).
Western blot assays
After the animals were anesthetized with ether, the hypothalamus and hippocampus were quickly separated and stored in liquid nitrogen. The tissues were pulverized, iced with appropriate amounts of lysate for 30 min, and centrifuged at high speed (4°C, 21 g) for 10 min. Protein was separated from the supernatant, and the concentration of it was quantified by bicinchoninic acid. After polyvinylidene fluoride electrophoresis, the protein was transferred to polyvinylidene fluoride membrane and sealed with 5% skim milk. The membrane was incubated overnight at 4°C, with antibodies targeting BDNF, TrkB, CREB, phosphorylated cAMP response element-binding protein (p-CREB), CRH-receptor 1 (CRHR1), FSH-receptor (FSHR), Bcl-2, Bax, or β-actin. The second antibody was incubated for 1 h at room temperature in a shaker, and the protein bands were obtained by chemiluminescence. The protein bands were numerically analyzed by ImageJ analysis software, and the expression levels of the protein bands were calculated based on the respective proportion of their β-actin proteins.
Statistical analysis
The data measured in this experiment were expressed as mean ± standard deviation. Calculations were validated by univariate homogeneity variance with SPSS 20.0, and comparison of between-group effects were performed by LSD. In this experiment, a P value of less than .05 was considered to indicate statistical significance.
RESULTS
Effects of THSWT on the change of body weight of OVX-OSA mice
The results in Figure 2A showed the changes in body weight of mice for 8 consecutive weeks. The weight of the control group increased gradually over time. The OVX mice initially gained weight briefly owing to estrogen deficiency, but they continued to lose weight under OSA stress. After the treatment of THSWT (0.5, 1.5, and 4.5 g/kg) or SI at the 4th week, the body weight of the mice was significantly reversed as expected.
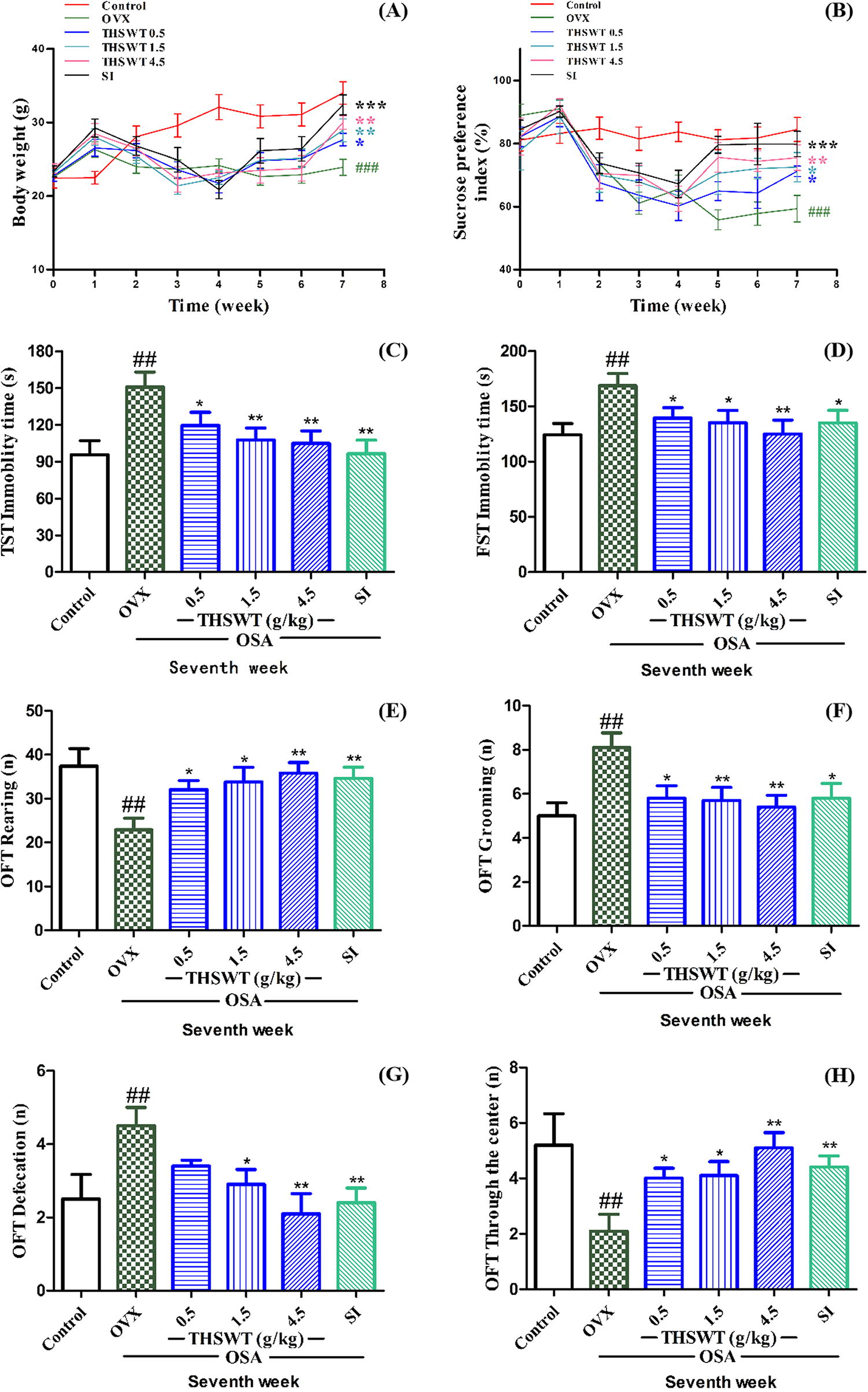
Effects of THSWT on the body weight and depression-like behaviors in OVX-OSA mice.
Effects of THSWT on pleasure deficiency, despair behavior, and memory impairment in mice
As shown in Figure 2B, in the sucrose preference test, the control group showed a preference for sucrose, indicating that normal mice preferred the pleasurable sensation produced by saccharin. The OVX mice showed a high sucrose preference index at the beginning of the experiment and we hypothesized that this was a self-protective behavior. The rationale behind providing ovariectomized mice with saccharin, which has a high caloric value, is that these mice tend to be weak and may benefit from the taste stimulation it offers. During the second week, the mice tended to be depressed owing to OSA stress. After the treatment of THSWT (0.5, 1.5, and 4.5 g/kg) or SI at the 4th week, the mice showed a significant increase in the sucrose preference index.
In TST (Fig. 2C) and FST (Fig. 2D) after 4 weeks of OSA, the immobility time of mice was significantly higher than that of the control group, whereas THSWT reduced the immobility time of mice from 150.8 to 104.9 s in TST, and it decreased the immobility time of mice from 165.7 to 119.0 s in FST.
OFT is a common behavior experiment in scientific research to evaluate the exploratory and depression activities of animals in a new environment. As shown in Figure 2E–H, OVX-OSA mice exhibited significantly less parental behavior and spent less time engaging in central activity compared with the control group. These findings suggest that the mice have lost their desire to explore. In contrast, OVX-OSA mice exhibited much grooming and defecation, which was a protective depression behavior. OVX mice showed significant depression-like symptoms after treatment with OSA; however, these behaviors were counteracted by THSWT (0.5, 1.5, and 4.5 g/kg) or SI, with the effect of the high dose of THSWT even surpassing that of SI. These results indicated that THSWT could improve the exploratory activity of mice.
Effect of THSWT on the uterine morphology and uterine index in OVX-OSA mice
As shown in Figure 3A, the uterine epithelial cells of mice in the control group and THSWT group were round in appearance. Compared with the control group, the uterine index of OVX mice decreased significantly, and the histopathological changes were obvious. In this study, THSWT (0.5, 1.5, 4.5 g/kg) or SI could effectively improve uterine lesions and uterine indices and reverse uterine growth inhibition.

Effect of THSWT on uterine index and morphology of OVX-OSA mice.
Effect of THSWT on the levels of gonadal hormones related to hypothalamic–pituitary–ovarian axis in OVX-OSA mice
The ovarian hormone levels in mice are shown in Figure 4. Compared with the control group, E2 levels of OVX-OSA mice in serum were lower, and FSH and LH levels in serum were significantly higher, as well as GnRH concentration in hypothalamus, which suggested that OVX can cause estrogen hypoplasia and ovarian hormone fluctuation in animals. Compared with the OVX group, 4.5 g/kg of THSWT increased serum E2 from 16.3 to 27.5 mIU/mL, and it reduced FSH and LH levels in serum by 22.1 and 2.6 mIU/mL, respectively. In addition, THSWT reduced GnRH by 159.4 mIU/mL in the hypothalamus.
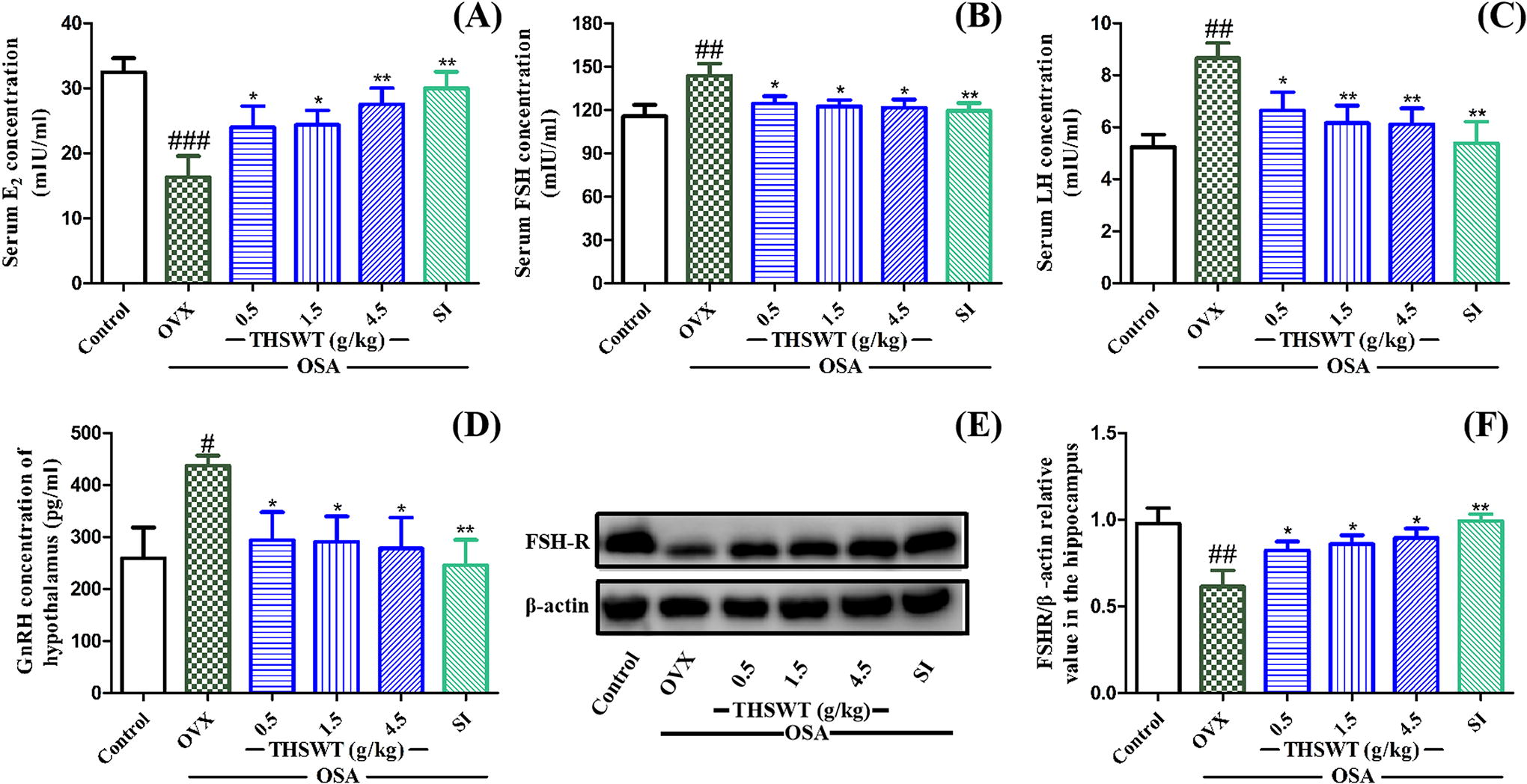
Effect of THSWT on ovarian hormones related to HPO axis in OVX-OSA mice.
The protein expression of FSHR in hippocampus tissues was also examined in this study. Figure 4E and 4F showed that FSHR protein expression level in the hippocampus of mice was significantly decreased when given OVX-OSA stress, which was improved by THSWT (0.5, 1.5, and 4.5 g/kg). These results suggested that TWSWT can promote the recovery of brain hippocampal tissue damage and improve depression-like behaviors.
THSWT treatment reduced OVX-OSA-induced hyperactivity of HPA axis
Compared with the control group, CORT and ACTH in serum and the concentration of CRH in hypothalamus and hippocampus were significantly increased in OVX-OSA mice. Compared with the OVX group, THSWT of 4.5 g/kg increased hypothalamus serotonin (5-HT) from 215.3 to 331.9 pg/mg, and it reduced CORT and ACTH levels in serum by 173.7 mg/mL and 23.7 ng/mL, respectively. In addition, THSWT reduced CRH by 2.6 ng/mL in the hypothalamus. Although low-dose THSWT had no significant effect on hippocampal CRH-R1, the overall treatment effect was clear (Fig. 5).
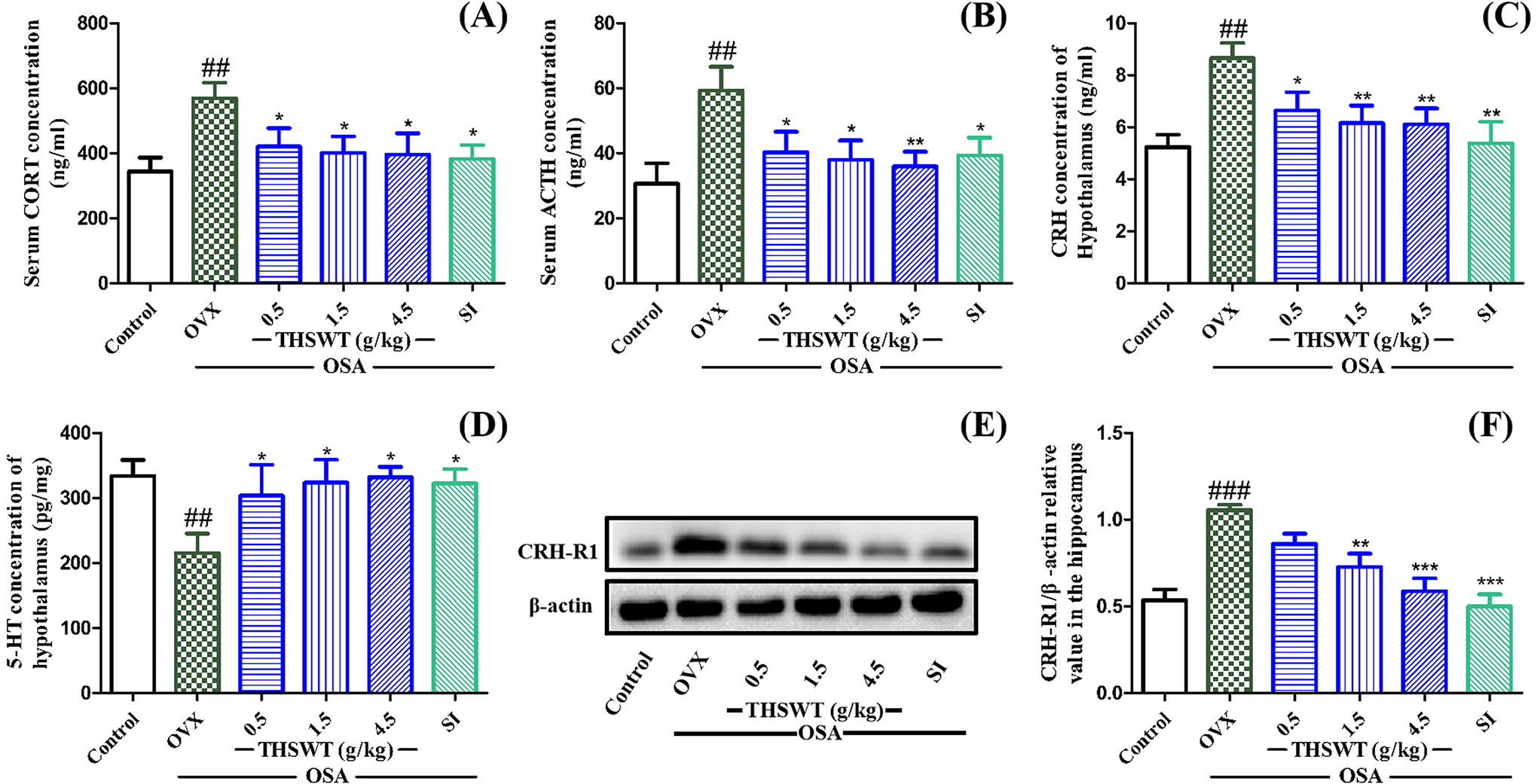
Effect of THSWT on the hormones related to HPA axis in OVX-OSA mice.
THSWT enhanced BDNF-TrkB-CREB signaling pathway in hippocampus in OVX-OSA mice
As shown in Figure 6, the effects of THSWT on the levels of BDNF, TrkB, Bax, Bcl-2, and p-CREB/CREB in the hippocampus of OVX-OSA mice are shown. The protein levels of BDNF and TrkB were significantly reduced in OVX-OSA mice compared with the control group, and THSWT treatment (0.5, 1.5, and 4.5 g/kg) effectively ameliorated these reductions. In addition, the expression of Bcl-2 was decreased and that of Bax was increased in OVX-OSA model group. Furthermore, there was no notable disparity in CREB phosphorylation between the control group and the OVX-OSA group. However, treatment with THSWT (0.5, 1.5, and 4.5 g/kg) significantly elevated the p-CREB-to-CREB ratio in OVX-OSA mice, with the high dose of THSWT exhibiting an effect similar to that of the control group. These results suggested that THSWT may play an antidepressant role by enhancing the BDNF-TrkB-CREB signaling pathway in the hippocampus.
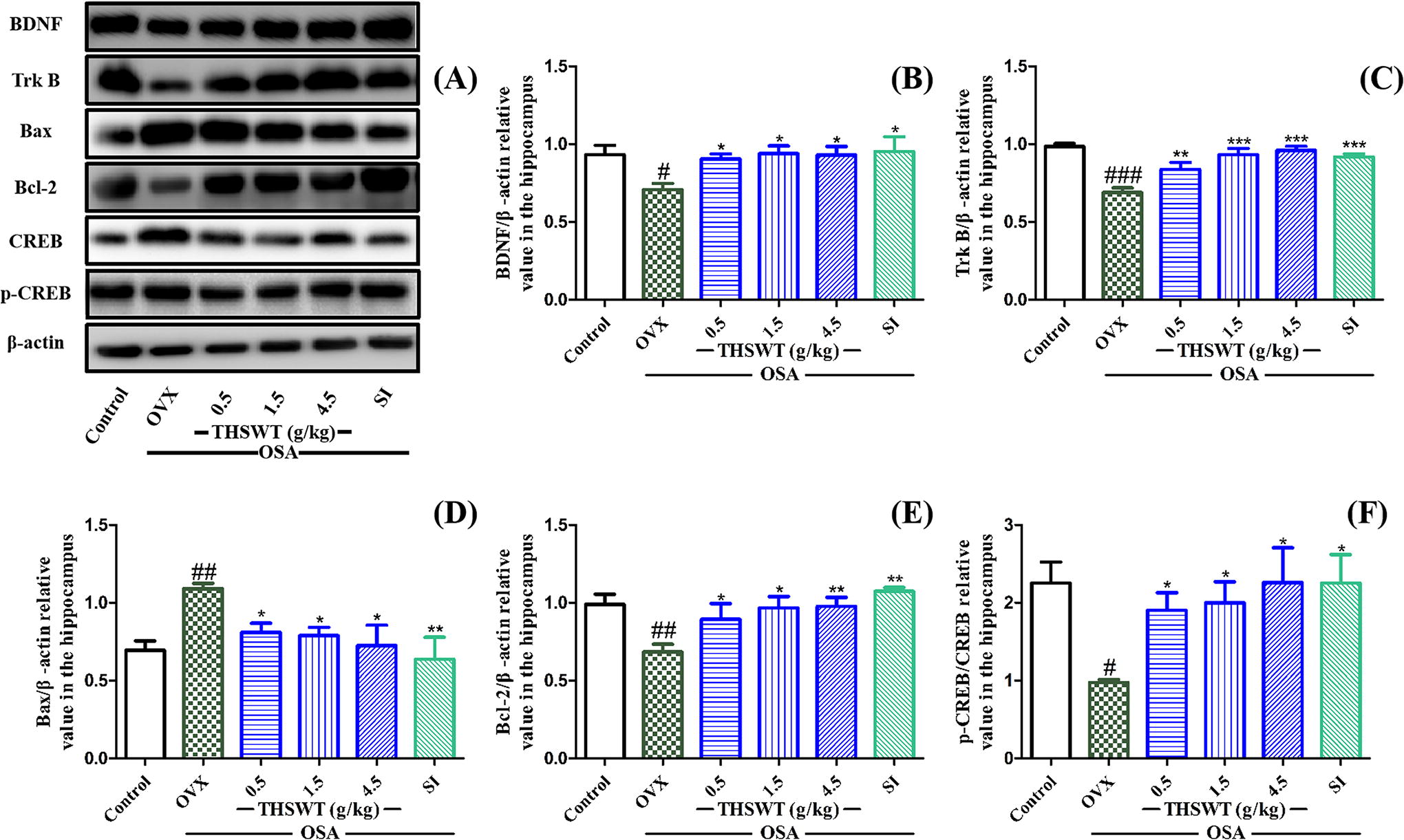
Effect of THSWT on the related protein expression of BDNF-TrkB-CREB pathway. The mice in OVX-OSA and treatment groups were operated by OSA from the 4th to the 7th week after ovariectomy. Data are presented as mean ± SD, # P < .05, ## P < .01, ###
DISCUSSION
This study investigated the antidepressant effect of THSWT on perimenopausal depression (PMD) and the possible mechanisms involved. The results demonstrated that THSWT effectively ameliorated depression-like behavior in OVX-OSA mice by increasing the sucrose preference index, reducing immobility time in tail suspension and forced swimming tests, increasing rearing actions, and decreasing depression-protective behaviors such as standing and defecation. THSWT was effective in mice with an OVX-induced uterine injury model, and increased the level of serum E2 and balanced gonadal hormones (LH, FSH, and GnRH) associated with the hypothalamic–pituitary–ovarian (HPO) axis. Compared with the model group, THSWT significantly reversed the HPA axis hyperactivity by reducing levels of GnRH and CRH in hypothalamus and CORT and ACTH in serum. In addition, it enhanced the expression of BDNF and TrkB proteins, increased the level of phosphorylated of CREB as well as antiapoptotic protein Bcl-2, and decreased the expression of apoptotic protein Bax in the hippocampus.
Currently, most studies have utilized chronic unpredictable mild stress (CUMS) to induce menopausal depression in mice. This involves subjecting animals to various external stressors on a daily basis during menopause and prolonged stimulation, resulting in the development of depressive symptoms. 28 However, in daily life, stress does not constantly stimulate the animal’s brain, and different forms of stress do not occur in a cyclical pattern. 13,29 Therefore, we selected OSA as the modeling method for this study since it is more likely to occur in reality than CUMS, so it has more clinical significance and can provide realistic reference for doctors to treat menopausal depression. In addition, OVX was used to establish the animal model of menopause.
Some evidence showed that abnormality of the HPO axis may be the direct cause of menopausal depression. The HPO axis regulates sex hormones by releasing GnRH from the hypothalamus, which, in turn, stimulates the pituitary gland to release FSH and LH into the bloodstream. These hormones then stimulate the ovaries to produce estrogen. In a study of women with PMD, estrogen levels in PMD women with HPG disorder tended to be much lower, yet GnRH, FSH, and LH levels were significantly higher in these patients compared with the normal group. 36 Although HRT has been considered as the mainstream treatment, much evidence indicates that women using HRT have a 2.3% (1.1–3.6%) increased risk of breast cancer during the perimenopausal period. 1 In the general population, approximately 4% of adult males and 2% of adult females are affected by OSA. However, OSA in women is more likely to increase the risk of depression during menopause. 37 In this study, OVX-OSA mice not only displayed typical depressive-like behavior but also had significantly elevated concentration of GnRH in hypothalamus and FSH and LH in serum. The concentration of ovarian estrogen E2 in serum was decreased. However, THSWT treatment can reduce GnRH, FSH, and LH levels by increasing serum E2 levels. The abnormal HPO axis of OVX-OSA mice returned to normal, and further improved the depression-like behavior as same as the positive group of SI. In addition, THSWT has a positive effect on the uterus of perimenopausal mice. These findings suggested that the antidepressant effect of THSWT on OVX-OSA mice may be attributed to its regulation of HPO axis.
As a key component of the HPO axis, FSH plays an important role in the biological function regulation of the HPO axis. The high serum FSH level is a signal of menopausal transition and is considered as a risk factor for women with PMD. 38 In rodents, FSH and FSHR have been found to be simultaneously located in the CA1 to CA4 regions of the hippocampus and cerebellar cortex, which play an important role in regulating the hormone levels and emotional stability of animals. 39 Inactivation mutations of FSHR have been demonstrated to result in primary or secondary amenorrhea, infertility, and premature ovarian failure. 40 In our study, the increase of serum FSH concentration in the OVX-OSA group demonstrated the dysfunction of the HPO axis in perimenopausal state. The expression of FSHR mRNA in the hippocampus was decreased in the current study, but no significant differences were observed, consistent with a previous report. 40 Compared with the model group, THSWT treatment significantly upregulated the mRNA expression of FSHR in OVX-OSA mice hippocampus. This finding is partly consistent with recent reports that FSHR deficiency increases cellular oxidative stress and that FSH-treated mouse neuroblastoma can exert antidepressant effects by exerting antioxidant effects. 40–41 However, the current study did not investigate which specific antioxidant factors are stimulated by FSH in cells, nor did it examine whether FSH affects upstream hormones to establish a negative feedback regulation. These issues need to be further investigated in the future.
Hyperactivation of the HPA axis is one of the most recognized neurobiological features in depression, which is mainly characterized by high levels of serum glucocorticoids (GCs, CORT in rodents, and cortisol in humans). Elevated serum cortisol levels and excessive negative feedback of the HPA axis are considered direct factors contributing to depression. 42 The main target of GCs is the hypothalamus, which is the main control center of HPA axis. Sustained high levels of GCs may damage hippocampal neurons by reducing dendrization of nerve cells, BDNF production, and neural tissue formation. When the HPA axis is activated (as in chronic physical injury), stressors stimulate hypothalamic neurons to release CRH, which, in turn, stimulates the anterior pituitary to produce ACTH. Once ACTH is released into the peripheral circulation, it acts directly on the adrenal cortex and stimulates the production of GCs into the systemic circulation. 43 In addition to GCs, CRH is an important part of the HPA axis, and has a significant effect on many neuropathies. CRH exerts physiological activity by binding to hormone receptors. Continuous stimulation of CRH-CRHR signaling pathway by stressors may inhibit the formation of signaling dendritic structures in developing hippocampal neurons. 44 Studies have demonstrated that knockout of the CRH-R1 gene or the use of CRH-R1 protein receptor antagonists can enhance the growth of dendritic structures during the formation of nerve cells. Furthermore, they can prevent anxiety-like behavior and mitigate the loss of synaptic function induced by chronic stress. 45–46 In this study, OVX-OSA induced significant hyperactivity of the HPA axis, that is, significantly increased CORT and ACTH in serum, as well as CRH in hypothalamus and hippocampus. The results showed that THSWT could reverse HPA axis abnormality by reducing CORT, ACTH, and CRH concentrations and protein expression of CRH receptor CRHR1. These findings suggest that THSWT may play an antidepressant role by regulating the HPA axis dysfunction induced by OVX-OSA in PMD mice. However, the results regarding the HPA axis in this study just provide one possibility. It remains unclear whether the reduction of CRHR1 protein following THSWT treatment directly contributes to the loss of synaptic function in neural cells.
BDNF plays a crucial role in depression-like behaviors and antidepressant responses as a key neurotrophin. 47 BDNF is essential in synaptic plasticity, neuronal survival, and formation in brain neural tissue, and it is active in the treatment of depression in humans or animals. 48 BDNF plays a neurotrophic role by binding to its protein receptor, TrkB, thereby activating the CREB signaling pathway. CREB serves as a downstream target of BDNF/TrkB, and phosphorylation of CREB can trigger the transcription of additional BDNF in the hippocampus. This process is important for many antidepressant treatments. 24 In general, the levels of BDNF were extremely low in women with perimenopausal syndrome and rats with abnormal estrogen. 49 It has been shown that estrogen receptor β (ERβ) knockout mice have lower BDNF levels than normal mice, and that estrogen or ERβ agonists can significantly enhance the physiological function of BDNF-TrkB signaling pathway in hippocampal neurons. 50 Bcl-2 and Bax are antiapoptotic and proapoptotic members of the Bcl-2 family, respectively. Bcl-2 interacts with Bax to prevent apoptosis, maintain the integrity of mitochondrial membrane, and prevent cytochrome C release from mitochondria to cytoplasm, which can promote the growth of hippocampal neurons and delay apoptosis to some extent. 51–52 In this study, the protein expression of BDNF, TrkB, and CREB in OVX-OSA mice were significantly downregulated, suggesting that the BDNF-TrkB-CREB signaling pathway was involved in the pathophysiological process of PMD. In addition, the increased phosphorylation level of CREB indicates that the BDNF-TrkB-CREB signaling pathway is, in turn, stimulated through the expression of BDNF in the hippocampus. Therefore, it is plausible to hypothesize that the antidepressant effect of THSWT on OVX-OSA mice may be associated with the activation of the BDNF-TrkB-CREB signaling pathway. However, it remains unclear whether THSWT influences the activities of other protein receptors closely associated to the BDNF-TrkB-CREB signaling pathway. In future studies, further exploration will be conducted to investigate the involvement of nerve growth factor and glial cell-derived neurotrophic factor in the BDNF-TrkB-CREB signaling pathway. Nerve growth factor and glial cell-derived neurotrophic factor are critical for the growth and proliferation of nerve cells, and their potential interactions within the BDNF-TrkB-CREB signaling pathway will be investigated.
TCM formula has been well accepted that its effects lie in the interaction between individual chemical components in the herbs with multitargets. Based on previous studies, we found that the main components of each herb in THSWT have an antidepressant pharmacological activity. Amygdalin in P. davidiana exhibits neurotrophic properties by activating the ERK1/2 pathway. 53 Curcumin in C. tinctorius is effective in the treatment of neurological diseases, such as Parkinson’s disease, 54 through improving the levels of glutathione and tyrosine hydroxylase in brain tissue. Ferulic acid, the main active compound in A. sinensis, displayed an antidepressant-like effects in corticosterone-induced mice model 55 and promoted neural progenitor cell proliferation in the hippocampus of corticosterone-treated mice in vivo. 56 Mannitol is the main component of R. glutinosa, which can increase the threshold of antielectroshock-induced seizures and reduce the percentage of epileptic response induced by γ-aminobutyric acid antagonist (pentylenetetrazole). Paeoniflorin, a monoterpenoid glycoside compound extracted from P. lactiflora, can improve the depressive behavior induced by reserpine in mice, which is characterized by prolonged activity time in the tail suspension and forced swimming experiments as well as ameliorating abnormal states of synaptic plasticity in the hippocampus. 57 Tetramethylpyrazine in L. wallichii can effectively improve neurobehavioral deficits and reduce neuronal apoptosis after subarachnoid hemorrhage in rats. 58 Additional research is needed to investigate the precise antidepressant mechanism of THSWT and identify the key active components within the formula.
CONCLUSIONS
This study demonstrated that THSWT has potential antidepressant effects in OVX-OSA-induced PMD mice model. THSWT alleviates hyperactivity of the HPG-HPA axis by improving serum hormone levels. At the same time, it can enhance the BDNF-TrkB-CREB signaling pathway in hippocampus. Few side effects of THSWT have been reported clinically and it can greatly ameliorate the serious side effects associated with hormone therapy. 1,59 In this study, we have confirmed the effectiveness of THSWT in the treatment of perimenopausal depression, but the study design has some shortcomings. For instance, OSA models do not completely replicate the actual symptoms of respiratory tract stenosis in animals. In addition, the specific components of THSWT that play a predominant role have not been definitively identified, and it remains uncertain whether they align with the intended mechanism we aimed to investigate in this study. Thus, the studies in the future will be focused on the investigation of the main active ingredients in THSWT.
Footnotes
Acknowledgments
The authors thank the staff of the Centre for Scientific Research of Zhejiang Provincial People’s Hospital for their valuable help in the experiment.
AUTHORS’ CONTRIBUTIONS
W.J.: Conception and design, acquisition of data, analysis and interpretation, literature searching, generating figures, and drafting the article. X.J.: Conception and design, data collection, analysis and interpretation, revising the article, and final approval of the version. W.H.: Literature searching, generating figures, and drafting the article. H.W.: Acquisition of data and analysis and interpretation.
AUTHOR DISCLOSURE STATEMENT
All authors declare that they have no conflicts of interest.
FUNDING INFORMATION
The study was supported by the