
Abstract
Lipolysis is the hydrolysis of triglycerides (TGs), commonly known as fats. Intracellular lipolysis of TG is associated with adipose triglyceride lipase (ATGL), which provides fatty acids during times of metabolic need. The aim of this study was to determine whether Coix lacryma-jobi L. var. ma-yuen Stapf (Coix) sprouts (CS) can alleviate obesity through lipolysis. Overall, we investigated the potential of CS under in vitro and in vivo conditions and confirmed the underlying mechanisms. Huh7 cells were exposed to free fatty acids (FFAs), and C57BL/6J mice were fed a 60% high-fat diet. When FFA were introduced into Huh7 cells, the intracellular TG levels increased within the Huh7 cells. However, CS treatment significantly reduced intracellular TG levels. Furthermore, CS decreased the expression of Pparγ and Srebp1c mRNA and downregulated the mutant Pnpla3 (I148M) mRNA. Notably, CS significantly upregulated ATGL expression. CS treatment at a dose of 200 mg/kg/day resulted in a significant and dose-dependent decrease in body weight gain and epididymal adipose tissue weight. Specifically, the group treated with CS (200 mg/kg/day) exhibited a significant modulation of serum lipid biomarkers. In addition, CS ameliorated histological alterations in both the liver and adipose tissues. In summary, CS efficiently inhibited lipid accumulation through the activation of the lipolytic enzyme ATGL coupled with the suppression of enzymes involved in TG synthesis. Consequently, CS show promise as a potential anti-obesity agent.
INTRODUCTION
The incidence of obesity has steadily increased in recent decades with consequent declines in both the average life expectancy and quality of life by detrimentally affecting body functions. 1 Obesity occurs when energy nutrients accumulate due to excess calorie intake, are converted to fat, and then stored in adipose tissue. Obesity is a vast global health problem that has placed a great economic burden on health care systems. 2 Nowadays, obesity has become a meaningful contributor to the exacerbation of morbidity and mortality, as it leads to respiratory disorders, weakens the immune system, and has implications for comorbidities, including hypertension, cardiovascular disease, kidney disease, and diabetes mellitus. 3,4 The US Food and Drug Administration (FDA)-approved anti-obesity drugs, including orlistat (pancreatic lipase inhibitor), phentermine/topiramate combination (appetite suppressant), liraglutide [glucagon-like peptide-1 (GLP-1) receptor agonists], and bupropion/naltrexone combination (opioid antagonist), have been prescribed for the purpose of treating obesity. 5 Especially Orlistat among these drugs has been widely prescribed for obese individuals with a body mass index (BMI) greater than 30 kg/m2 or greater than 27 kg/m2 with other concomitant risk factors (e.g., hypertension, diabetes, dyslipidemia). However, this drug has severe adverse effects, such as oily stools, fecal spotting, abdominal pain, subacute liver failure diarrhea, cholestatic hepatitis, and cholelithiasis. 6 Therefore, studies are needed on safe and effective drug targets for obesity management. Considering these points, herbal therapy has rapidly emerged as an essential component of human health care because it has few adverse effects and is a cost-effective alternative. 7
Lipid droplets (LDs) store fat and provide high-energy substrates for mitochondrial β-oxidation. Obesity, also known as metabolic syndrome, is characterized by abnormal buildup of LDs in the liver. 8 The degradation of triglycerides (TGs) stored in LDs, followed by their oxidation in mitochondria, a process called cytosolic lipolysis, is associated with such a process. 9 Lipolysis is defined as the catabolism (hydrolysis) of TGs stored in cellular LDs, which eventually splits them into fatty acids (FAs) and glycerol. At this time, adipose triglyceride lipase (ATGL), an important lipolytic enzyme, has been continuously investigated since its discovery in 2004. 10 ATGL is a crucial enzyme for releasing FAs from TG stores to produce FAs, which are needed for energy production in mitochondria. In addition, a variant (148M) in patatin-like phospholipase domain-containing protein 3 (PNPLA3), linked to the buildup of TGs in the liver, exists at high levels in LDs. 11 In this study, ATGL and PNPLA3 (I148M) compete for TG breakdown in LDs. 12
Coix lacryma-jobi L. var. ma-yuen Stapf (Coix) is a variety of Coix lacryma-jobi L. known as adlay or Job’s tears, and its seeds are primarily produced in Korea, China, Japan, Taiwan, and India. 13 Coix seeds are obtained by removing the peel after drying and are considered an alternative to rice because of their higher protein content (14%) compared to rice. Its bioactive components, including coixol, coixenolide, coixan, and total triterpenes, exert pharmacological effects on various diseases such as hypertension, asthma, arthritis, cancer, immunological disorders, and gastrointestinal and skin diseases. 14 –16 Consequently, numerous studies have reported that Coix seeds possess beneficial activities such as antioxidant, anti-inflammatory, antiapoptotic, antiproliferative, and diuretic effects. 17 –19 In addition, they exhibit hypoglycemic effects. 20 Coix sprouts (CS) are the young buds or leaves of Coix harvested approximately 10 days after sowing the seeds. 14 Interestingly, a recent study demonstrated that germinated Coix had higher antioxidant activities than ungerminated Coix. 21
In our previous research, we documented the anti-obesity effects of CS, which centered on the downregulation of factors linked to TG and Total cholesterol (TC) synthesis through the activation of LKB1/AMPK pathways. 22 Nevertheless, investigations into the mechanisms related to factors responsible for TG breakdown have been inadequately conducted. Therefore, as stated in the Introduction, our aim was to confirm the pharmacological effects and mechanisms of action of CS using a combination of cellular and animal experiments.
Based on these reports, we hypothesized that CS might be able to reduce TG accumulation by increasing lipolysis. Therefore, we confirmed our hypothesis through cellular experiments and investigated the inhibition of TG synthesis in an High-fat diet (HFD)-treated mouse model.
MATERIALS AND METHODS
Materials
Protease inhibitors were purchased from Wako Pure Chemical Industries, Ltd. (Osaka, Japan). Proteins from Santa Cruz Biotechnology, Inc. (TX, USA): SREBP-1, Histone, ACC, FAS, SCD1, ATGL, and β-actin. Secondary antibodies were purchased from GeneTex, Inc. (CA, USA). Coixol was purchased from ChemFaces (Shanghai, China). HBSS was purchased from Sigma-Aldrich.
Plant materials
Coix, known as ‘Johyun,’ is a novel adlay variety that was cultivated by the Rural Development Administration in Jeonju-si, Jeollabuk-do, Korea, in 2004. The dried CS was stored at 4°C until required. To attain the target initial moisture content, the CS was submerged in room-temperature distilled water (DW) for 24 h and then subjected to a 48-h drying process at 20°C using an oven. Germination was carried out in a growth chamber under controlled lighting conditions (metal halide lamp illumination), with temperature and relative humidity set at 27°C and 98%, respectively, following the methodology from a prior experiment. 23 The extraction condition of the dried CS was boiled at 100°C for 2 h, and the yield was 20%.
Analyses of coixol
Dried CS powder (1 g) was mixed with 10 mL of pure methanol. Extraction was performed by ultrasonic shaking at room temperature for 30 min. The extract was filtered and analyzed by high-performance liquid chromatography (HPLC) using a 10 μL sample injected into the 1260 Infinity II LC System with an Agilent Eclipse Plus C18 column (4.6 × 250 mm, 5 μm). The mobile phase consisted of water containing 0.1% acetic acid (solvent A) and acetonitrile containing 0.1% acetic acid (solvent B). The gradient elution conditions were as follows: A:B = 82:18 (0 min), 82:18 (35 min), 0:100 (36 min), 0:100 (60 min), 82:18 (61 min), and 82:18 (70 min). The flow rate was 1.0 mL/min, and detection was performed at 290 nm using UV absorption. Coixol was detected as a major compound in the extract chromatograms. The coixol peak was identified by comparing its retention time and UV spectra with those of authentic standards. A representative HPLC chromatogram is shown in Figure 1. A calibration curve was established using coixol concentrations ranging from 62.5 to 1000 μg/mL, with 10 μL injected for each of the five replicates. The standard curve yielded a regression coefficient (R 2) of 1 with the equation y = 14.243x + 35.741. In addition, the analysis revealed that the coixol content of the extract was 18.54 ± 0.11 mg/g.
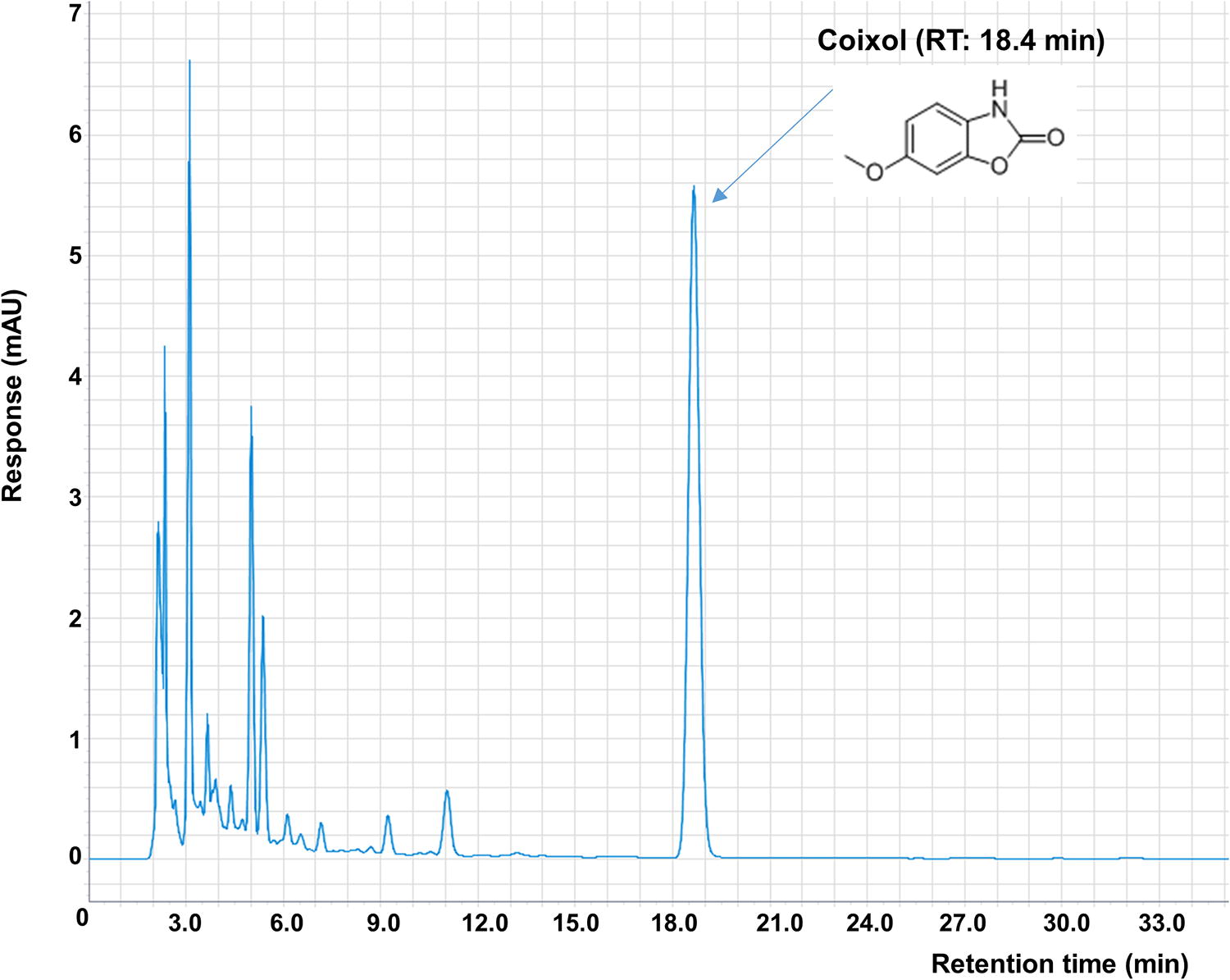
Analysis showed a coixol content of 290 nm in the CS extract.
The cell culture and cell proliferation assay
To validate the potential mechanisms of action of CS, we cultured Huh7 cells derived from hepatocellular carcinoma in high-glucose DMEM supplemented with 10% fetal bovine serum (FBS) and 1% penicillin. The cells were cultured in a 5% CO2 environment at 37°C. Initially, Huh7 cells were seeded in a 96-well microplate at a density of 2 × 103 cells/well to confirm cell proliferation and division. After seeding, the cells were treated with the respective drugs for 6 h. After drug treatment, the cells were further incubated for 48 h in the presence of free fatty acids (FFAs) such as oleic acid or bovine serum albumin. Finally, 10% of the reagent from the WST-8 Cell Proliferation Assay Kit was added to the cells and incubated for 1 h in a cell incubator. Measurements were performed at two wavelengths, 450 and 600 nm, using a microplate reader (VersaMax, Molecular Devices, Germany).
Measurement of intracellular triglyceride
Huh7 cells were seeded in 6-well plates at a concentration of 1 × 106 cells/mL. They were pretreated with CS at 100, 200, and 400 μg/mL for 6 h. After the pretreatment, the cells were exposed to 300 μM of oleic acid and subsequently cultured for an additional 48 h. Cells were detached using trypsin-ethylenediaminetetraacetic acid and washed by centrifugation. Next, a lipid extraction buffer (consisting of a 2:1 mixture of Methanol:Chloroform) was used to collect the lipid layer during fractionation. In this study, TG was quantified using a commercial kit (TR22421, Thermo Fisher Scientific, Waltham, MA, USA). The absorbance was measured at 450 and 600 nm using a microplate reader.
Measurement of lactate dehydrogenase release
The Lactate dehydrogenase (LDH) activity was assessed using a commercial assay kit (BioVision, Cat# K726, Waltham, MA, USA). Cell-free supernatant or lysate (40 μL) was mixed with nicotinamide adenine dinucleotide hydrogen (NADH) and 2,4-dinitrophenylhydrazine and incubated at 37°C for 15 min. NaOH (0.4 mol/L) was added and incubated for 3 min at 37°C. The absorbance was measured at 440 nm using a UV spectrophotometer (Infinite M200, Tecan, Salzburg, Austria).
Mice treatment
All procedures were performed in accordance with the Guidelines for the Use and Care of Laboratory Animals (DHU 2021-067, Daegu Haany University, Daegu, Korea), and the protocol was approved by the Institutional Committee for the Care and Use of Laboratory Animals. Healthy 5-week-old male C57BL/6J mice were obtained from DBL Co., Ltd. (Eumseong, Korea). All mice were housed under the following conditions: temperature (23 ± 2°C), humidity (50 ± 10%), and 12-h light/12-h dark cycle. All mice (excluding the normal group; n = 8) were fed a 60% HFD (Diet 12492; Research Diets, Inc., New Brunswick, NJ, USA) for 5 days to acclimatize them to the feeding regimen. After the acclimation period, HFD-fed obese mice were randomly allocated to four groups (n = 8/group): HFD Control (administered water), GG200, (administered Garcinia gummi-gutta extract 200 mg/kg/day), CS100 (administered 100 mg/kg/day), and CS200, administered CS 200 mg/kg/day. Drug treatments were administered orally for 5 weeks. GG, which was used as a positive control, is widely known for its anti-obesity effects, which are associated with the suppression of fatty acid synthesis. We performed a comparative analysis of factors related to fatty acid synthesis and lipid metabolism in the GG200 experimental group and the CS treatment group. 24 Furthermore, we previously reported the anti-obesity effect of CS in a mouse model induced by a 60% HFD. This effect involves the inhibition of fat synthesis through activation of LKB1/AMPK and suppression of the SREBP-1 and SREBP-2 pathways, which are involved in TG and cholesterol synthesis. Therefore, the concentrations used in this experiment were consistent with those used in our previous study conducting mechanistic research related to TG breakdown.
Histological examination
Liver and adipose tissues were examined microscopically. Adipose tissue was fixed in 10% neutral-buffered formalin, embedded in paraffin, and stained with hematoxylin and eosin (H&E). Liver sections were stained with Oil Red O (ORO) to analyze lipid accumulation. Frozen liver sections were mounted on ProbeOn-Plus slides (Thermo Fisher Scientific, Waltham, MA, USA) and subjected to ORO staining followed by counterstaining with Harris hematoxylin. Stained sections were examined using i-Solution software (IMT i-Solution Inc., Scarborough, ON, Canada).
Analysis of serum biochemistry
The blood was centrifuged at 1500×g for 10 min at 4°C. Serum TG and TC levels were measured using commercially available kits from Wako through spectrophotometry. High-density lipoprotein cholesterol (HDL-C) was measured using a kit supplied by Asan Pharmaceutical Co., Ltd. (cat. AM203; Hwaseong, Korea). Low-density lipoprotein cholesterol (LDL-C) was calculated from the measured levels of TG, TC, and HDL-C using the following formula: LDL-C level (mg/dL) = (TC-[HDL-C]-TG/5). Similarly, very low-density lipoprotein cholesterol (VLDL-C) was calculated using the TG level as follows: VLDL-C level (mg/dL) = TG/5.
Gene expression analyses
Huh7 cells were plated at 2 × 106 cells/dish in a 65-mm dish. On the next day, the cells were exposed to different drug concentrations for 6 h prior to treatment. An additional treatment of 300 μM oleic acid was administered, followed by washing with PBS after 48 h. mRNA was extracted from the Huh7 cells using TRIzol chloroform extraction. Following mRNA isolation, cDNA synthesis was carried out using the commercially available SuperScript™ One-Cycle cDNA Kit (Cat. A10752030, Thermo Fisher Scientific). Following that, quantitative polymerase chain reaction (qPCR) was conducted using the StepOnePlus Real-Time PCR system (Applied Biosystems, Foster City, CA, USA). The qPCR primers used are listed in Table 1.
Primer List for Real-Time Polymerase Chain Reaction
Analysis of immunofluorescence assay
An IF assay was conducted to examine the impact of CS on TG hydrolysis. Huh7 cells were cultured in high-glucose DMEM with 10% FBS and 1% penicillin in a 5% CO2 atmosphere at 37°C. Huh7 cells (2 x 105 cells/mL) were seeded in a 35-mm dish and were pretreated with CS at concentrations of 100, 200, and 400 μg/mL for 6 h. Following this, an additional treatment of 300 μM oleic acid was administered, and the cells were cultured for an additional 48 h. After the treatment period, the cells were washed and fixed with 4% paraformaldehyde at room temperature for 1 h. Subsequently, the cells were washed thrice with PBS for 10 min each, and 2.5% normal goat serum was added. After blocking, ATGL was applied at a ratio of 1:200, respectively, and incubated at 4°C for 18 h. The cells were then washed twice with Phosphate-buffered saline with Tween 20 (PBST) and once with PBS. Next, the cells were treated with BODIPY (D3821, Thermo Fisher) and incubated at 37°C for 30 min to observe the lipid droplets. Finally, cells were stained with 4′,6-diamidino-2-phenylindole (DAPI). IF images were captured using a light microscope (Olympus BX51; Tokyo, Japan).
Western blotting
To obtain the cytosolic fraction, liver tissues were lysed in buffer A, and buffer C was used to collect the nuclear fraction. The detailed method of Western blotting was performed as described previously. 16 Each transferred membrane was blocked in a 5% (w/v) skim milk solution for 1 h and then visualized using an ECL solution. A Sensi-Q 2000 ChemiDoc Imaging System (Lugen Scientific Co. Ltd., Gyeonggi-do, Korea) detected the bands.
Statistical analysis
SPSS 26.0 software (IBM, Armonk, NY, USA) was used for the statistical analysis. All results were expressed as the mean ± Standard error of the mean (SEM). Statistical comparisons were performed using One-way ANOVA, followed by Tukey’s post hoc test. If P < .05, the data were considered statistically significant.
RESULTS
CS reduced cell proliferation without cytotoxicity
To ascertain the effect of CS on cell proliferation in Huh7 cells, we used the WST-8 assay. Cell proliferation was assessed 24 h after treatment with specified concentrations of CS. Notably, cell proliferation was significantly inhibited by CS200 and CS400. Furthermore, the results of the LDH release assay revealed no meaningful cytotoxic effects following treatment with CS concentrations of 100, 200, and 400 μg/mL. Throughout the study, we selected the range 100–400 for subsequent analyses (as shown in Fig. 2).

Effect of CS treatment on cell proliferation through WST-8 and LDH release.
CS attenuated intracellular TG contents
The TG content was quantified in Huh7 cells after treatment with CS. The TG content decreased in a concentration-dependent manner after CS treatment. In particular, CS400 showed dramatically reduced TG levels compared with those in the FFA Control group (Fig. 3).

Effect of CS treatment on intracellular TG contents in FFA-induced Huh7 cells.
CS decreased mRNA expressions encoded for TG synthesis
We measured the mRNA expression of genes encoding transcription factors involved in TG synthesis, namely Pparγ, Srebp-1c, and Pnpla3 (I148M). FFA treatment significantly increased the expression of all three genes [Pparγ, 1.73-fold; Srebp-1c, 1.62-fold; Pnpla3 (I148M), 2.62-fold], whereas CS treatment led to marked reductions. Notably, in the CS400 treated cells, these increased levels were noticeably decreased by 39.5%, 45.2%, and 50.1%, respectively (Fig. 4).
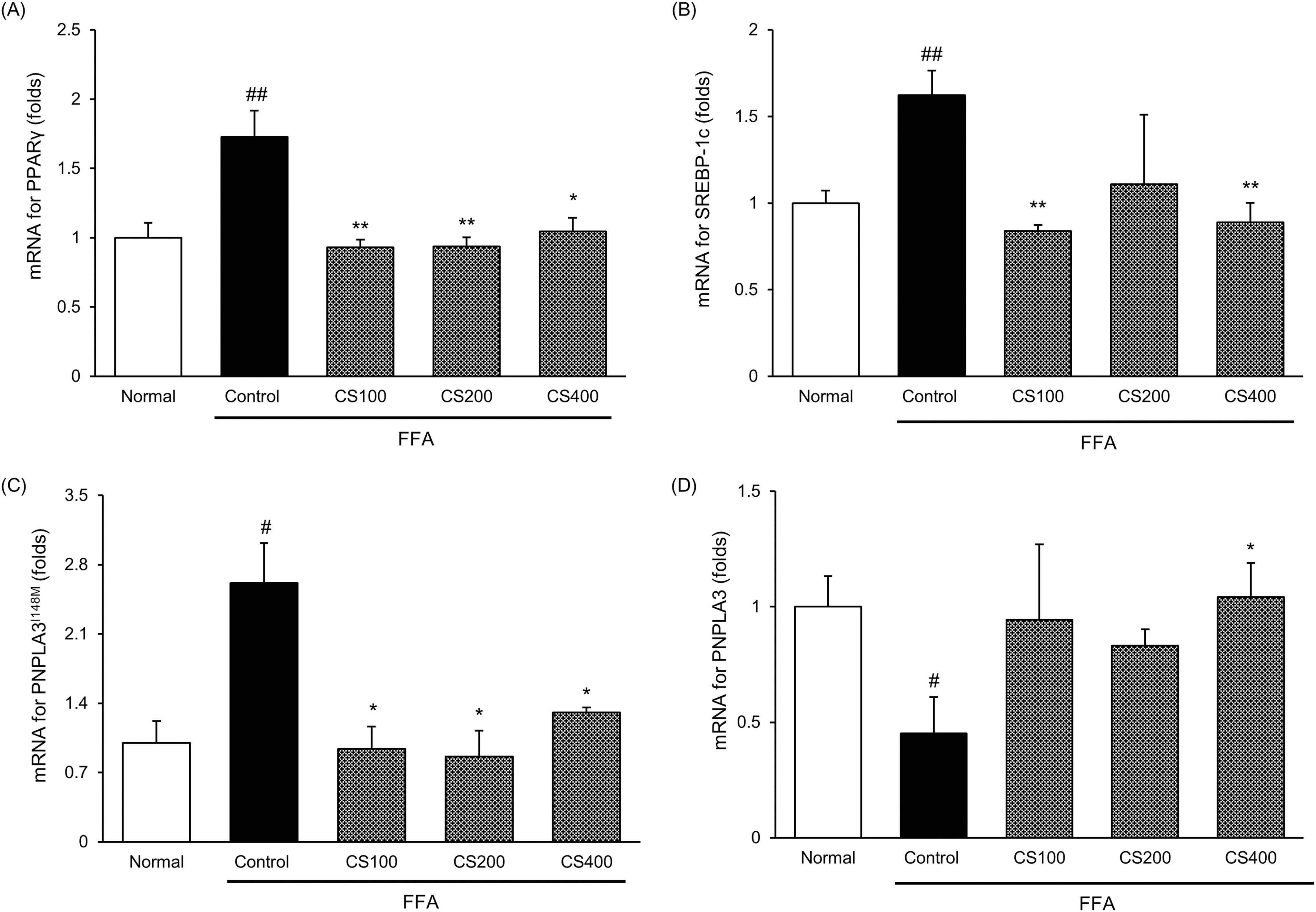
Effect of CS treatment on gene expressions encoded for TG synthesis in FFA-induced Huh7 cells. The mRNA levels of
CS led to lipase activation such as ATGL
Exposure of Huh7 cells to FFA resulted in reduced ATGL activity. Interestingly, CS treatment upregulated the ATGL activity in a dose-dependent manner. In the FFA Control group, ATGL activity was dramatically reduced to 60.5% of that in the normal group. However, under CS treatment, ATGL activity was improved by 1.4-, 1.6-, and 2.0-fold for the respective concentrations compared with the FFA Control group (Fig. 5).
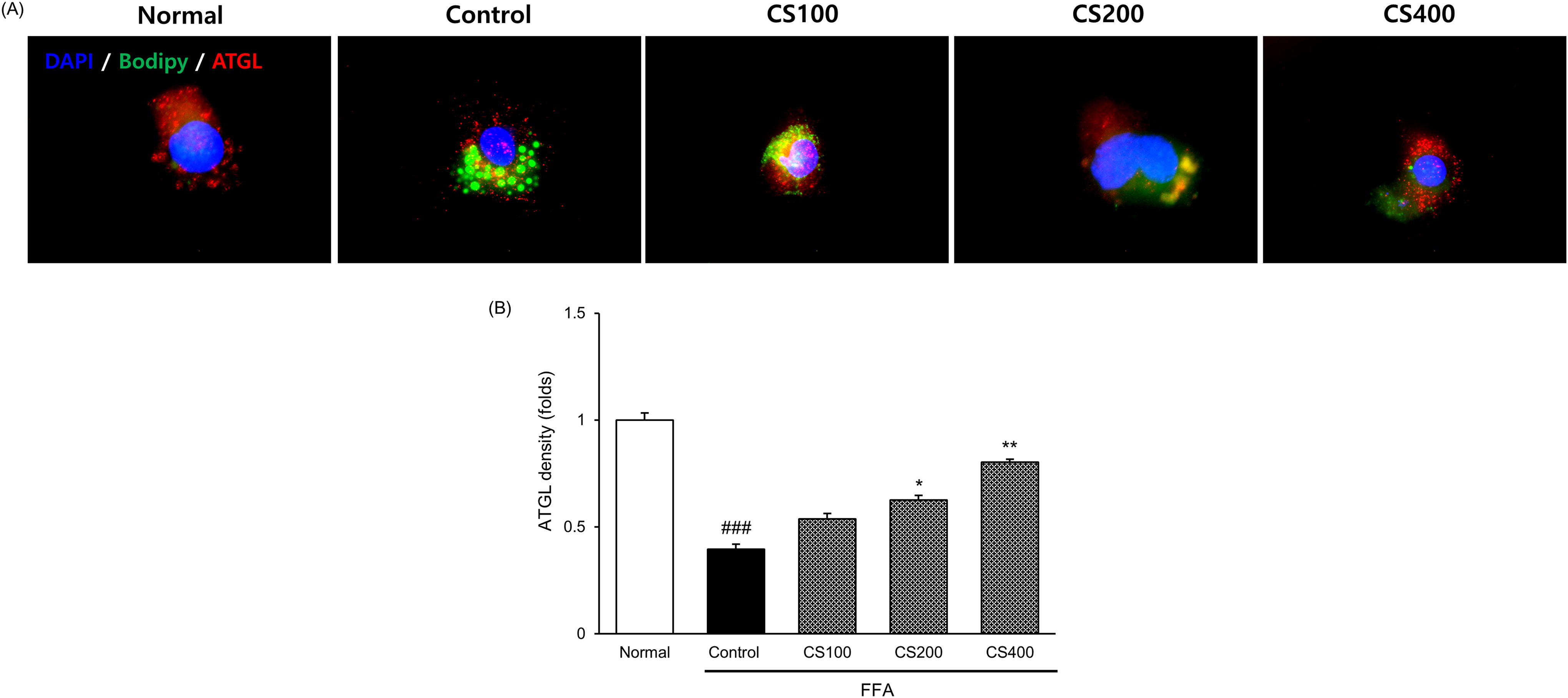
Effect of CS treatment on ATGL activation in FFA-induced Huh7 cells.
CS regulated body weight gain and liver and epididymal adipose weights in HFD-fed obese mice
Table 2 presents the body weight gain and liver and epididymal adipose tissue weights observed throughout the experimental period. HFD supplementation significantly increased body weight gain (P < .001) and reduced epididymal adipose weight compared with the HFD Control group (P < .001). In contrast, CS treatment led to a concentration-dependent reduction in body weight gain and epididymal adipose tissue weight. Notably, the CS100 and CS200 groups exhibited significantly reduced epididymal adipose tissue weights, with the CS200 group showing a particularly substantial reduction of 9.9% compared with the CS100 group (CS100, 16.6%; CS200, 26.5% vs. HFD Control group). Surprisingly, there was no significant difference in the liver weights, although a decreasing trend was observed. Collectively, these results suggested that CS treatment effectively reduced body fat mass in HFD-fed obese mice.
Body Weight Gain and Liver and Epididymal Adipose Tissue Weights
Control, HFD Control mice; GG200, GG 200 mg/kg-treated and HFD fed mice; CS100, CS 100 mg/kg-treated and HFD fed mice; CS200, CS 200 mg/kg-treated and HFD fed mice. Data are the mean ± SEM. (n = 8).
Significance: ### P < .001 versus the Normal mice and * P < .05, ** P < .01, *** P < .001 versus the HFD Control mice.
CS, Coix Sprouts; GG, Garcinia gummi-gutta.
CS improved serum lipid profiles in HFD-fed obese mice
In general, an HFD increases fat mass and negatively affects serum lipid profiles. Therefore, we aimed to determine whether CS exerts anti-obesity effects on serum lipid profiles. Treatment with CS200 resulted in a notable decrease in serum TGs, TC, and LDL-C levels of 33.8%, 24.4%, and 29.5%, respectively. In addition, CS200 increased HDL-C levels by 40.4% compared with the HFD Control group. Notably, CS200 exhibited more pronounced effects than CS100 (Fig. 6).
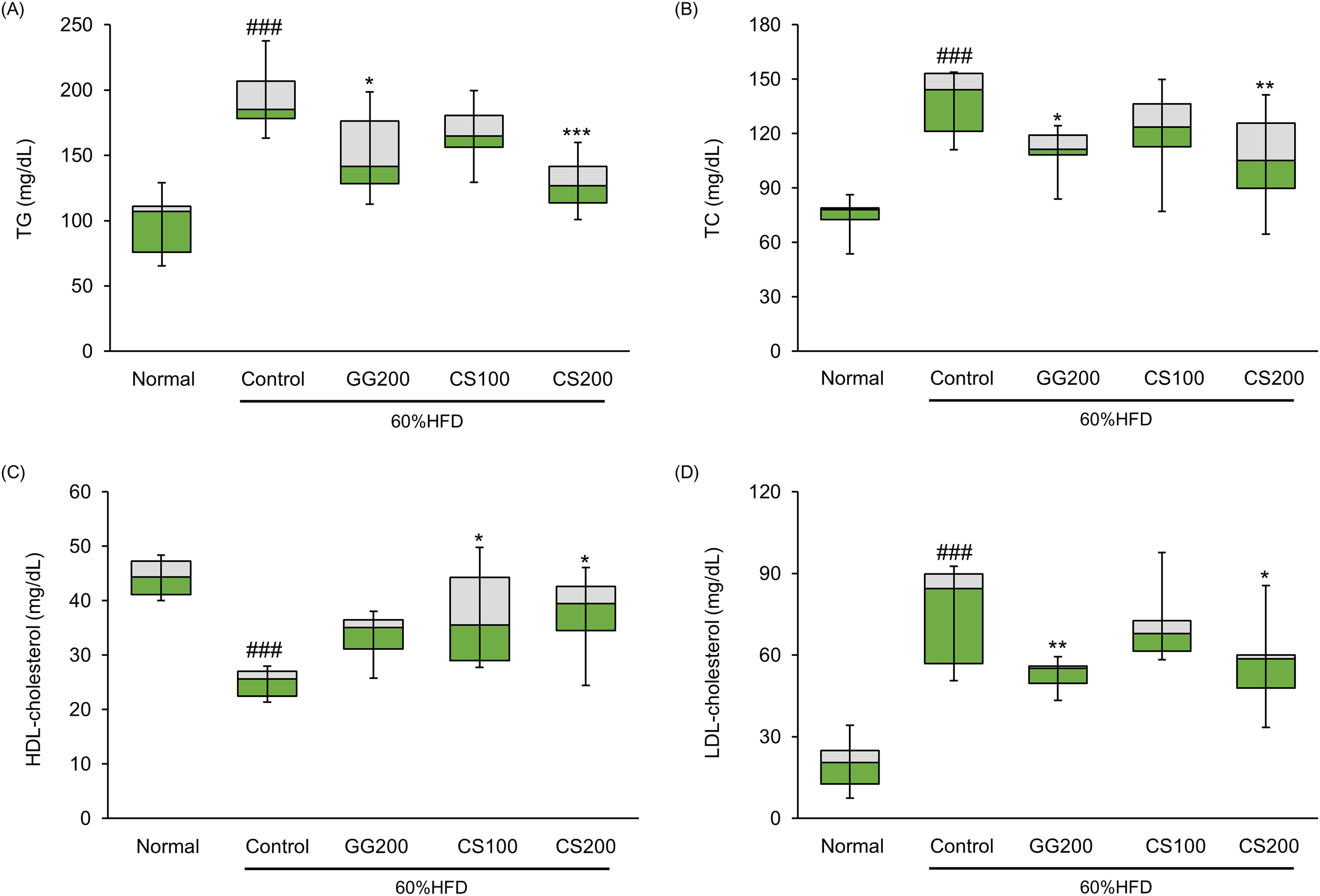
Effect of CS treatment on serum lipid profiles in HFD-fed obese mice. Serum lipid profiles include
CS improved the enzymes associated with TG synthesis in HFD-fed obese mice
Overall, the immunoblotting results revealed that HFD intake increased the expression of the transcription factor SREBP-1, as depicted in Figure 7. However, the administration of CS200 significantly inhibited this elevated SREBP-1 (P < .01). Suppression of SREBP-1 resulted in a reduction in the expression of lipogenic genes such as ACC and FAS. These findings suggest that CS effectively regulates the expression of proteins related to TG synthesis.

Effect of CS treatment on protein expressions associated with TG synthesis.
CS alleviated the histopathological changes by HFD induction
Histological alterations in the mice were evaluated using H&E and ORO staining, as depicted in Figure 8. We investigated whether the enhanced lipid metabolism led to alterations in lipid accumulation. In the Normal group, liver sections showed neatly arranged cells without LD accumulation or infiltration of inflammatory cells. In contrast, the HFD Control group exhibited numerous LDs adjacent to liver cells, accompanied by significant infiltration of inflammatory cells. Furthermore, H&E staining showed that the adipocyte size in the CS200-treated group was noticeably reduced compared with that in the HFD Control group. Consequently, CS treatment mitigated the pathological conditions more effectively than in the HFD Control group.
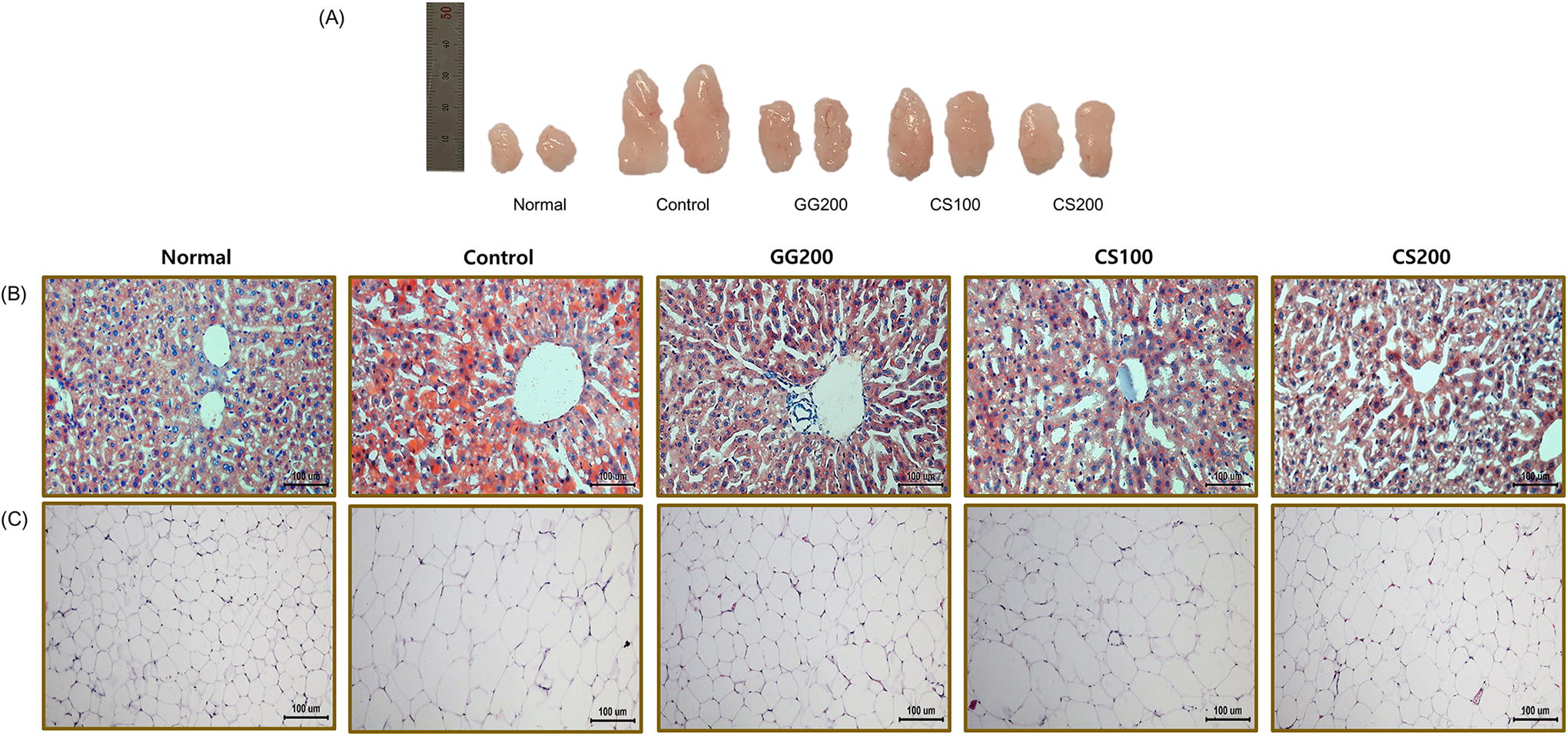
Effect of CS treatment on the histological alterations in the liver and epididymal adipose tissue of high-fat diet-fed obese mice.
DISCUSSION
The worldwide prevalence of obesity has significantly increased in recent decades. Many obese people suffer from a variety of metabolic disorders, including hypertension, heart disease, type 2 diabetes mellitus, and cancer. 25 Lifestyle modification is recommended for reducing abnormal body weight; however, drug intervention is essential for morbidly obese patients. 26 In addition, several dietary supplements containing herbs, their active ingredients, minerals, or dietary fiber aimed at weight loss are gaining popularity and are now available on the market. 27 So far, numerous anti-obesity medications have not been licensed for long-term usage due to an elevated risk of adverse effects or inadequate efficacy. Therefore, researchers are focusing on the development of new anti-obesity drugs that are highly effective and have minimal adverse events. 28
The liver, fat tissue, and skeletal muscle are fundamental sites for the metabolic regulation of fatty acids and the maintenance of lipid homeostasis. The liver is not primarily intended to store fat; instead, it serves a specific function in maintaining and regulating homeostatic lipid flux under normal circumstances. Imbalanced lipid metabolism in the liver, observed in conditions such as obesity and diabetes, can lead to fat accumulation. 29 This accumulation marks the beginning of nonalcoholic fatty liver disease (NAFLD), a condition encompassing steatosis, nonalcoholic steatohepatitis, liver fibrosis, cirrhosis, and hepatocellular carcinoma. 30 NAFLD has emerged as a global epidemic in recent years. Thus, the inhibition of fat accumulation is emerging as an effective strategy for controlling both obesity and NAFLD.
Lipids must be stored and used intracellularly to maintain cellular energy balance. Hepatic TGs are synthesized from dietary FFA stored as LDs and then hydrolyzed into fatty acids for energy. While increased TG reserves are normally inactive and not particularly detrimental to cells, the overaccumulation of FFA and TC may induce a cytotoxic state. 31 For that reason, multiple mechanisms associated with the regulation of hepatic lipid levels exist, including lipolysis, the oxidation of fatty acids, autophagy, de novo synthesis, and the release of lipoprotein particles. 32 Among which TG lipolysis, controlled by the expression of lipases, is an emerging strategy for improving obesity. Particularly, ATGL and HSL are key enzymes in the process. 33 During periods of increased energy demand, LDs, primarily consisting of triacylglycerols (TAGs), undergo TAG hydrolysis to release fatty acids (FAs) through processes such as lipolysis or lipophagy. These processes are tightly modulated by the regulator like insulin and catecholamines. 34 Lipolysis is primarily facilitated by the action of ATGL, HSL, and monoacylglycerol lipase (MGL), which sequentially convert TAGs into glycerol and FAs.
We used CS, which contained coixol at a concentration of 18.5421 mg/g (Fig. 1). Huh7 (a human hepatoma cell line) was evaluated using the WST-8 assay. With CS200 and CS400, a clear antiproliferative effect was observed. Importantly, we confirmed that the decrease in cell proliferation was not due to cytotoxicity, as shown by the LDH assay (Fig. 2).
Next, we conducted an intracellular TG accumulation assessment using IF staining (Fig. 3). 35 Intracellular TG levels, which were used to evaluate lipolysis, were considerably higher in the FFA group than in the normal group; however, they decreased in a concentration-dependent manner. Notably, the intracellular TG levels in the CS400 group were similar to those in the normal group. In addition, we measured the mRNA expressions of genes involved in transcriptional regulation related to TG synthesis, including Pparγ, Srebp-1c, and Pnpla3 (I148M). Several studies investigating the anti-obesity effect have suggested the suppression of Pparγ, a key transcription factor in adipogenesis and lipogenesis. 36,37 Furthermore, in mammalian cells, patatin-like phospholipase domain-containing protein 3 (PNPLA3) is primarily associated with LDs. The PNPLA3 gene is encoded by adiponutrin (ADPN), a 481 amino acid protein. Substitution of methionine with leucine occurs at amino acid position 148 of ADPN because of the sequence variation in PNPLA3 (I148M). 38 The human PNPLA3 gene is moderately expressed in adipose tissue, skin, kidney, and brain, but highly expressed in the liver. 39,40 However, the PNPLA3 gene is highly expressed in mouse adipose tissue. PNPLA2 (also known as ATGL) and abhydrolase domain containing 5 (ABHD5) typically work together to break down TGs from LDs. The I148M mutation disrupts PNPLA3 turnover, leading to LD accumulation by reducing ATGL activity. 41 However, PNPLA3 knockout mice maintained normal plasma and hepatic TG levels and did not develop fatty liver disease (FLD). Ultimately, the I148M mutation causes steatosis and loss of function, increasing vulnerability to hepatotoxins. 42 Moreover, PNPLA3 is regulated by SREBP1c in mouse and human hepatocytes. 43 Similarly, in our results, FFA treatment significantly increased the expression of three genes, while CS treatment markedly reduced these elevated levels. In the CS400 treatment, these increased levels were noticeably decreased by 39.5%, 45.2%, and 50.1%, respectively (Fig. 4). ATGL expression in FFA-treated Huh7 cells was considerably reduced, indicating that FFA administration prevented FFA breakdown. However, CS400 treatment largely reversed the decrease in ATGL expression (Fig. 5). A comprehensive analysis of the cellular experiments showed that both CS200 and CS400 inhibited cell proliferation without inducing cytotoxicity, with no significant differences between the two groups. In addition, intracellular TG concentrations (μg/mg protein) were reduced considerably at both concentrations, but no significant difference was seen. Finally, considering the density of ATGL and the mRNA levels of PNPLA3 (I148M) competing for TG breakdown within LDs, we concluded that PNPLA3 (I148M) mRNA levels were lower in CS200 than in CS400. Based on these results, we determined the optimal dosage for the animal experiments.
First, we hypothesized that CS exerts anti-obesity effects in HFD-fed obese mice, a commonly used experimental model for hyperlipidemia and hyperglycemia studies. Table 1 displays the body weight gain, as well as the liver and epididymal adipose weights, during the experimental period. The mean initial body weights did not show significant differences among all groups, ranging from 19.82 g to 21.52 g. Normal mice had a slight increase in body weight gain during the 5-week experimental period, significantly different from the HFD Control group (P < .001). All drug-treated groups showed significantly reduced body weights compared with the HFD Control group (GG200, P < .01; CS100, P < .05; CS200, P < .01). In addition, epididymal adipose tissue weights in both the CS100 and CS200 groups significantly decreased by 16.6% and 26.5%, respectively. Furthermore, HFD-fed mice exhibited a noticeable increase in body weight gain and an improvement in dyslipidemia, similar to those observed in mice treated with GG200. The HFD induced an increase in the serum levels of lipid factors, including TGs, TC, and LDL-C (P < .001). Elevated TG and TC levels significantly decreased in all of the groups, but HDL-C levels increased and LDL-C levels decreased only in the GG200 and CS200 groups, respectively. When comparing the GG200 and CS200 groups, GG200 was more effective at reducing LDL-C levels (18.1%), whereas CS200 was more effective at reducing TG levels (7.8%) (Fig. 6). In general, HFD intake led to increased expression of the transcription factor SREBP-1. However, CS200 supplementation significantly inhibited this elevated protein level (SREBP-1, P < .01). Subsequently, inhibition of SREBP-1 resulted in the reduced expression of lipogenic genes (ACC and FAS). These findings indicate that CS treatment effectively modulated the expression of proteins involved in TG synthesis (Fig. 7).
We also analyzed the effects of CS on the liver and adipose tissues of HFD-induced obese mice. Histopathological examination of the liver, as assessed by ORO staining, demonstrated that CS treatment markedly attenuated increased lipid accumulation in the liver. As shown in Figure 8, fat depots were lower in CS-treated groups than in the HFD Control group, similar to the findings of a previous study’s findings. 22 H&E staining of epididymal adipose tissue showed that the size of adipocytes in CS-treated mice was smaller than that in HFD Control mice. Therefore, CS treatment improves HFD-induced obesity by regulating TG lipolysis.
In the TG breakdown process, two key enzymes, ATGL and HSL, ultimately break down TG into glycerol and fatty acids. However, in this study, the evaluation of the final product was insufficient. It would be valuable to assess the changes in the final products in future research. Furthermore, it is necessary to evaluate changes in TG accumulation in the liver tissues to determine whether there has been an improvement in the liver tissue.
CONCLUSION
In conclusion, our study highlighted the potential of CS as a promising agent for the treatment of obesity. CS has shown a remarkable ability to prevent lipid accumulation by effectively targeting enzymes involved in TG synthesis, activating the lipolytic enzyme ATGL, and inhibiting PNPLA3 (I148M). These findings suggest that CS may be a valuable addition to the arsenal of therapies for obesity.
Footnotes
AUTHORS’ CONTRIBUTIONS
Mi-Rae Shin and Min Ju Kim contributed equally to this study. All authors reviewed and approved the final version of the article for publication.
AUTHOR DISCLOSURE STATEMENT
There are no financial conflicts of interest to declare.
FUNDING INFORMATION
This study was supported by the “Cooperative Research Program for Agriculture Science and Technology Development (