
Abstract
This study investigated the alleviating effect of fermented ginsenosides obtained through yeast strain fermentation transformation on acute liver injury (ALI) induced by CCl4. Strains were screened for their ability to produce β-glucosidase, the transformation ability of the strain was verified by high-performance liquid chromatography, and the Saccharomyces cerevisiae strain F6 was obtained by 26S rRNA sequencing. After fermentation by F6 strain, it was found that the content of ginsenosides Re, Rb1, and Rb2 was significantly decreased (P < 0.05), and rare ginsenosides were detected, with the content of Rh4 and Rg5 reaching 2.65 mg·g−1 and 2.56 mg·g−1. We also explored the preventive effect of fermented ginsenoside extract (FGE) on ALI. Mice were evenly divided into 9 groups as follows: control group, ALI model group, positive drug bifendate group, and treatment group, which included 3 ginsenoside extract (GE) groups and 3 FGE groups (dosage of 150, 300, and 450 mg·kg−1 b.w.). The results showed that compared with the ALI model group, FGE significantly increased the levels of glutathione peroxidase, hydroperoxidase, and superoxide dismutase and also decreased the malondialdehyde level. The levels of alanine aminotransferase, aspartate aminotransferase, and total bilirubin markers were significantly reduced, and the levels of inflammatory cytokines TNF-α, IL-6, and IL-1β were significantly decreased. Bioinformatics analysis combined with Western blot validation explored the molecular mechanism of the effect of FGE. It was found that FGE could downregulate the expression of the p-AKT/AKT and the p-mTOR/mTOR ratios. These results suggested that FGE played an alleviative role in ALI by promoting autophagy to inhibit the AKT/mTOR signaling pathway.
INTRODUCTION
The liver is the main detoxification organ in the human body, which can convert toxic substances produced by external or internal metabolism into nontoxic or highly soluble substances and then excrete them from the body. 1 More than 1000 drugs could cause liver toxicity, such as anticancer, analgesic, and anti-inflammatory drugs, leading to acute liver injury (ALI). 2 –4 ALI could cause liver cirrhosis and liver cancer, posing a serious threat to human life and health. The use of traditional chemical hepatoprotective drugs could lead to side effects and increase the burden on the liver. It was urgent to develop a liver protective drug with minimal side effects and minimal liver damage. 1,5 Therefore, it is important to develop new, safe, and effective hepatoprotective drugs.
Ginseng (Panax ginseng C.A. Meyer) had been used as a traditional herbal medicine or dietary supplement in China and Asia for more than 2,000 years. 6 Ginsenosides are the main active ingredient in ginseng, among them, ginsenosides Rg5 and Rh2 had a significant role in preventing liver injury, which has been confirmed. However, due to its rarity, it could only be obtained by breaking the glycosidic bonds of ginsenosides Re and Rb1 by means of transformation. 7 –9 The conversion methods of ginsenosides included physical conversion, chemical conversion, and biological conversion. 10 Among them, biotransformation had the advantages of mild reaction conditions, high specificity, and environmental protection. 11 Fermentation is an effective biological transformation method, and the glycosidases produced during its metabolic process could hydrolyze the sugars in ginsenosides, thereby transforming ginsenosides and producing new rare ginsenosides. 12,13 For instance, an Aspergillus niger strain converted ginsenoside Rb1 to Rg3 in the presence of β-glucosidase. 14 The β-glucosidase produced by armillaria converted ginsenoside Rb2 into the rare ginsenoside CK. 15 According to reports, rare ginsenosides had better biological activity, and the transformation of ginsenosides had high research and development significance. 13,16
In this study a strain was selected by screening to transform ginsenosides in panax ginseng and analyzed the components of the transformed ginsenosides using high-performance liquid chromatography (HPLC). We also established a CCl4 induced mice ALI model to study the efficacy of fermented ginsenosides for alleviating ALI. We also used comprehensive bioinformatics analysis to explore potential signaling pathways and verified the main metabolic pathways regulated by FGE in the treatment of ALI using Western blot, aiming to further develop and utilize ginseng, and provided innovative strategies for the treatment of ALI.
MATERIALS AND METHODS
Screening of fermentation strains
Fresh grapes were mashed and naturally fermented for 14 days, and the fermentation liquor was coated on YEPD (yeast extract peptide glucose) solid medium then incubated for 48 h at 28°C. Using the p-nitrophenol method, strains that produced β-glucosidase were screened, and the transformation ability of each strain was verified using HPLC after separation and purification; the purified strains were sequenced at 26S rRNA. Genomic DNA was extracted using the ZR fungal/Bacterial DNA MiniPrep Kit. The purities and concentrations of the extracted DNA were determined by NanoDrop analysis. The extracted 26S rRNA was amplified by polymerase chain reaction with the universal primers NL1 (5′-GCATATCAATAAGCGGAGGAAAAG-3′), NL4 (5′-GGTCCGTGTTTCAAGACGG-3′). The same primers were used for sanger sequencing, and the fermented strain sequences were screened and compared using the NCBI BLAST online database system (https://blast.ncbi.nlm.nih.gov/Blast.cgi). A phylogenetic tree of identified sequences was generated using the molecular evolutionary genetic analysis software (MEGA 7).
Preparation of fermented ginseng extraction
Ginseng (Panax ginseng C. A. Meyer) was sourced from Wanliang (Jilin, China) in May 2023. The botanical identification was carried out by Professor Z. He, and a voucher specimen (No. 20230411) was maintained at the College of Food Science and Engineering, Jilin Agricultural University, China. Fresh ginseng was washed and dried, then crushed and sieved through a 200 μm mesh filter. A 100 g sample of ginseng powder was added to 1200 mL of distilled water and then added 4% F6 strain and was fermented at 28°C for 6 days. Another 100 g of ginseng powder was added to 1200 mL of distilled water and were both decocted and extracted for 2 h, respectively. After filtration, the corresponding extraction solution was obtained, and 1:1 water saturated n-butanol was added for extraction 3 times. The extraction solution was combined, concentrated, and freeze-dried to obtain fermented ginsenoside extract (FGE) and ginsenoside extract (GE), respectively.
HPLC analysis of fermented ginsenosides
One gram of ginseng powder was added to 12 mL of distilled water and prepared the corresponding FGE and GE according to methods mentioned above. The samples were dissolved in 2 mL of chromatographic methanol, passed through a 0.22-μm filter membrane, and then analyzed using HPLC. The chromatographic column was an Analytical C18 column (4.6 × 250 mm, 5 μm, PerkinElmer, USA), the mobile phases were 0.05% phosphate acid water (A) and acetonitrile (B), and the gradient elution methods are described in Table 1. The flow rate was 1.0 mL·min−1, and the injection volume of each sample solution was 10 μL. The sample chamber temperature of the autosampler was 4°C, and the column temperature was maintained at 35°C.
Gradient Elution of Ginsenosides Contained in the FGE
Animal experimentation
Seventy-two male ICR mice (SPF, 20–22 g, 7-weeks old) were purchased from Liaoning Changsheng Laboratory Animal Co. Ltd. (Liaoning, China) and bred in SPF barrier environment, with constant temperature of 21 ± 2°C, relative humidity of 40%–70%, and 12 h light/12 h dark (6:00 a.m. to 6:00 p.m.) cycle, and given ad libitum access to food and water. The animal experiments were approved by the Laboratory Animal Welfare and Ethics Committee of Jilin Agricultural University with the ethical review acceptance number 20230616004.
After a week of acclimatization, 72 mice were randomly divided into 9 groups on average, including the normal control (CON) group and ALI model group, which were both given saline by gavage during the experiment. The positive drug bifendate (BIF) group was daily given at a dose of 120 mg·kg−1 (body weight, b.w.) BIF dissolved in saline. The low-dose GE (L-GE) group, medium-dose GE (M-GE) group, and high-dose GE (H-GE) group were daily given at 150, 300, and 450 mg·kg−1 (b.w.) GE, respectively. The low-dose FGE (L-FGE) group, medium-dose FGE (M-FGE) group, and high-dose FGE (H-FGE) group were daily given at doses of 150, 300, and 450 mg·kg−1 (b.w.) FGE, respectively. The intervention lasted for 14 days.
After 14 days of intervention, mice in each group were intraperitoneally injected with 50% CCl4 oil solution at the dosage of 1 mL·kg−1 (b.w.) after the last gavage for 2 h, whereas mice in CON group were only injected with olive oil. After fasting overnight for 12 h, blood was collected and then the mice were euthanized. After coagulation at room temperature for 60 min, the plasma was centrifuged at 3000 g for 15 min at 4°C to obtain the mice serum, which was then stored at −80°C.
Histopathological assessment
After killing the mice, fresh liver tissue samples were frozen and sectioned, and the sections were stained with hematoxylin and eosin (H&E). Stained sections were examined, and the histopathological changes in the sections were evaluated. Histological assessment of liver tissue is based on the severity of inflammation, the structural integrity of the liver plate and hepatic cord, and the integrity of the hepatocytes.
Determination of biochemical parameters
Inflammatory factors IL-6, IL-1β, and tumor necrosis factor (TNF)-α in mouse serum samples were measured by enzyme-linked immunosorbent assay. Alanine aminotransferase (ALT), aspartate aminotransferase (AST), superoxide dismutase (SOD), malondialdehyde (MDA), glutathione peroxidase (GSH), hydroperoxidase (CAT), and total bilirubin (T-BIL) indicators were measured using a microplate reader.
Bioinformatics analysis
Gene databases were used to search for targets of “Acute liver injury” and potential targets of the major ginsenosides in the FGE (http://www.swisstargetprediction.ch/) and plotted on a Venn diagram in (https://www.bioinformatics.com.cn/login/); common targets of FGE and ALI were obtained. These common targets were imported into the String database (https://string-db.org/) to obtain the protein–protein interaction (PPI) network and were imported into the Metscape database (https://metascape.org/gp/index.html/main/step3) to obtain Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genome (KEGG) enrichment analysis.
Western blotting
Total protein was extracted from the liver tissue for denaturation and isolation by 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then transferred to polyvinylidene fluoride membrane. After blocking with 5% skim milk for 2 h, the membranes were incubated with the primary antibody (1:1000) at 4°C overnight. After incubation, the membrane was washed thrice with Tris-Buffered Saline with Tween (TBST), and the membrane was incubated with alkaline phosphatase-conjugated secondary antibody (1:2000) for 2 h. Finally, the target proteins were visualized using the ALP chromogen (NBT/BCIP) and analyzed for their intensity relative to β-actin. The signal intensity was captured by the ChemiDoc MP imaging system, and the protein bands were analyzed using ImageJ analysis software.
Statistical analysis
Data were presented as means ± standard deviations (SD). Statistical significance was analyzed using analysis of variance and followed by t test; P < 0.05 was statistically significant.
RESULTS AND DISCUSSION
Strain screening and characterization
After 14 days of fermentation, 10 strains with the ability to produce β-glucosidase were obtained. The enzyme activity of the 10 strains were measured by p-nitrophenol test, and the results are shown in Figure 1A; the results showed that strains F6, D5, A3, A5, and R1 had strong β-glucosidase production capacities. The transformation capacity of these 5 strains with high enzyme activity was identified by HPLC, these strains could convert ginsenosides, so the lower the residual content of ginsenoside Re, the stronger the transformation ability of the strain, as shown in Figure 1B. The F6 strain could reduce the content of ginsenoside Re from 4.98 mg·g−1 to 1.84 mg·g−1 (P < 0.01), indicating that the F6 strain had better enzyme production activity and stronger transformation ability compared with other strains. We next used BLAST analysis of 26S rRNA sequencing results, and 15 strains with the highest similarity to F6 (percent identity > 99.9%) were selected, and phylogenetic tree analysis was performed using the neighbor-joining algorithm with 1000 bootstrap in MEGA 7.0 software in Figure 1C. The identification of isolates based on the highest fit match resulted in strain F6 being identified as Saccharomyces cerevisiae.

β-glucosidase competence and Re transformation ability determination and identification of strain.
Analysis of fermented ginsenoside components
The content determination analysis based on HPLC showed that the F6 strain could effectively convert ginsenosides. As shown in Table 2, after fermentation, the content of ginsenosides Rb1, Rb2, and Re in FGE significantly decreased, from 5.05 mg·g−1, 2.24 mg·g−1, and 1.81 mg·g−1 in GE to 2.56 mg·g−1, 1.28 mg·g−1, and 1.04 mg·g−1 in FGE, respectively. The content of rare ginsenosides in FGE significantly increased, and ginsenosides Rh4, Rh2, and Rg5 were detected, with concentrations of 2.65 mg·g−1, 1.07 mg·g−1, and 2.56 mg·g−1, respectively. The standard curve and methodological study for the content determination are shown in Table 3 and Table 4, and the HPLC chromatograms of GE, FGE, and ginsenoside standard substances are shown in Figure 2. It showed that the F6 strain could transform common ginsenosides in ginseng and increase the content of rare ginsenosides in ginseng. Studies have shown that the generated ginsenosides Rh2 and Rg5 could activate the LKB1/AMPK/mTOR signaling pathway and modify the gut microbiota to alleviate liver injury. 8
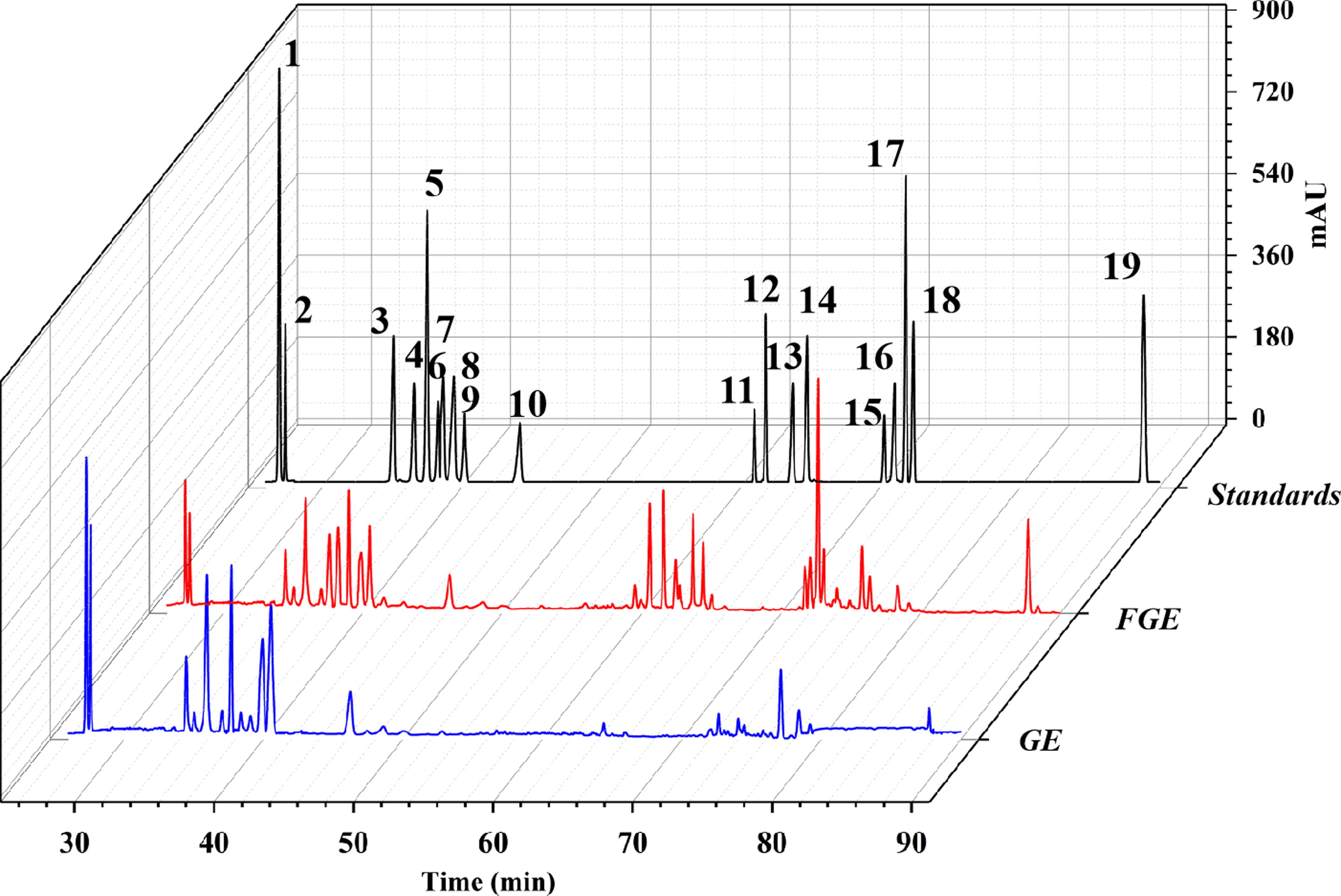
HPLC chromatograms of the ginsenoside standards, fermented ginsenoside extract (FGE), and ginsenoside extract (GE). These ginsenosides were 1-Rg1, 2-Re, 3-Rf, 4-Rb1, 5-Rc, 6-Rh1, 7-Rg2, 8-Rb2, 9-Rb3, 10-Rd, 11-Rk3, 12-Rh4, 13-Rg3, 14-PPT, 15-CK, 16-Rk1, 17-Rg5, 18-Rh2, and 19-PPD.
Content and Variation of Ginsenosides Before and After Fermentation
Linear Relationship and Standard Curve of the Ginsenosides Contained in the FGE
Methodological Investigation of the Ginsenosides Contained in the FGE
Histological evaluation
The histological changes in the mice were evaluated in H&E-stained liver sections. As shown in Figure 3, the hepatocytes in the CON group were intact and tightly organized, with no inflammatory cell infiltration. The ALI group developed severe structural disturbances of the hepatic cords and hepatic lobules with marked inflammatory infiltrates associated with large areas of inflammatory necrosis. The hepatocytes in the FGE group showed partial local necrosis with no obvious inflammatory infiltration and clear cellular structure. With the increasing dose, the necrosis decreased in the FGE group, and the cell structure became clearer, and the hepatic lock membrane and hepatic sinusoids appeared normal. The results showed that FGE had alleviated CCl4-induced ALI in a concentration-dependent manner. M-GE and H-GE in the GE group also reduced the degree of inflammatory cell infiltration, but they were not as effective as the M-FGE and H-FGE groups.
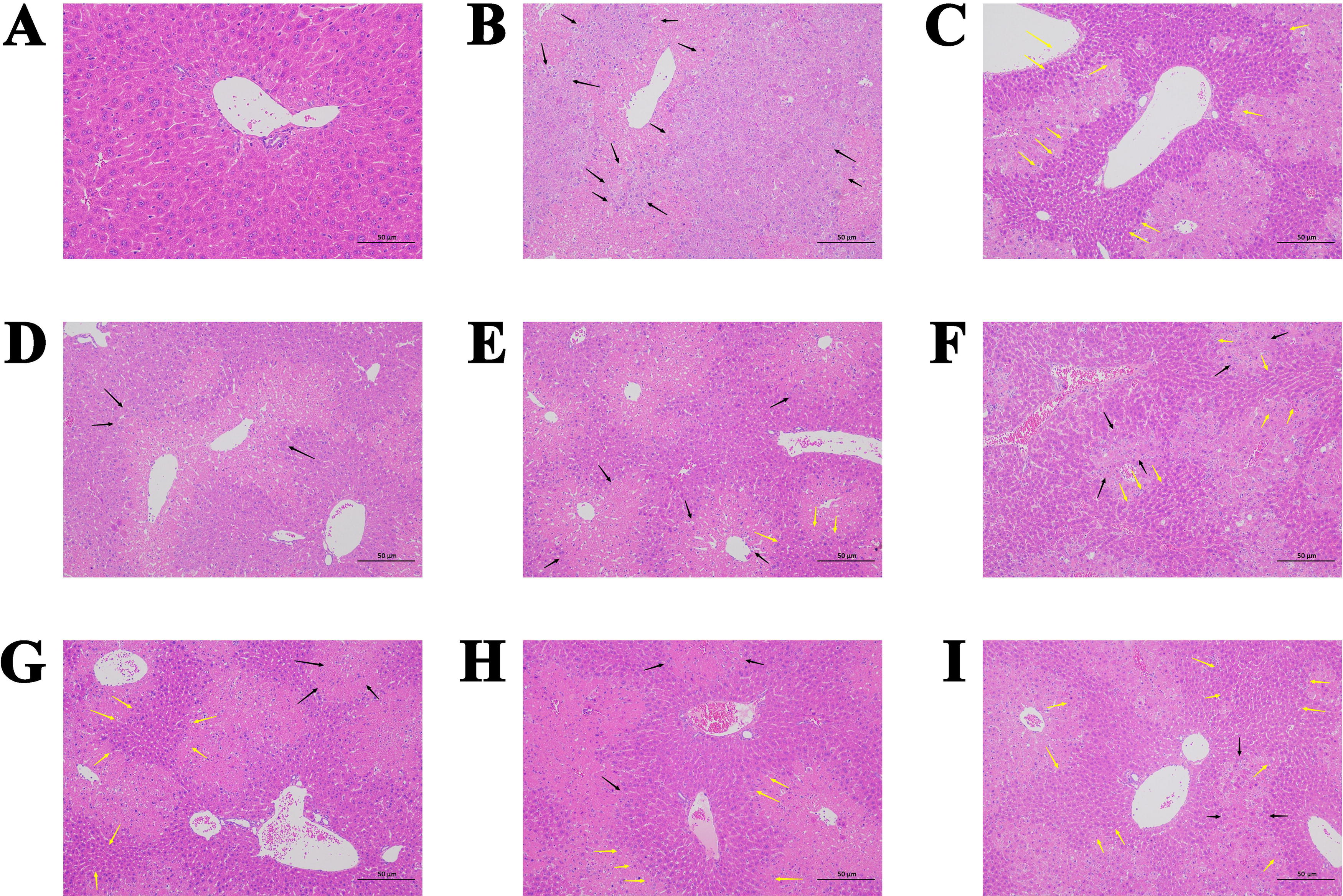
The images of mouse liver sections stained with H&E to show histopathology of livers (magnification: 200×, Scale bar: 50 μm).
Effect of FGE on the serum markers in mice
As shown in Figure 4A, B, and C, compared with the CON group, the serum levels of ALT, AST, and T-BIL in the ALI group were significantly higher (P < 0.01), indicating damage to the liver tissue. Compared with the ALI group, L-FGE group could significantly reduce the serum level of ALT (P < 0.05), in the M-FGE group the serum levels of AST and T-BIL were lower (P < 0.05), H-FGE group exhibited much lower serum levels of ALT, AST, and T-BIL (P < 0.01), and the BIF group was consistent with the H-FGE group.
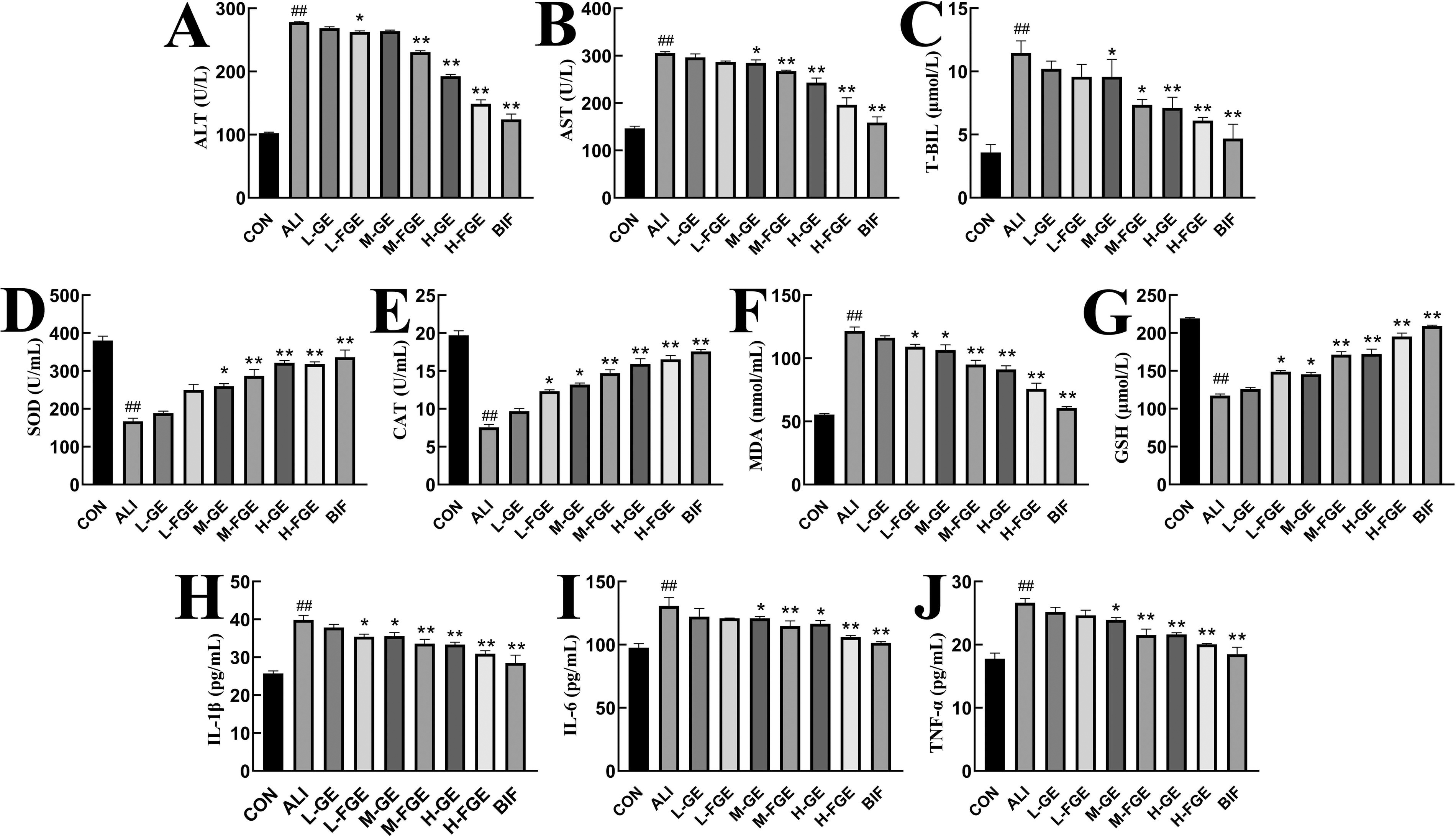
Biochemical analysis of serum after CCL4 injection in different groups. All data are expressed as the means ± standard deviations, n = 8. Compared with the CON group, ## P < 0.01; compared with the ALI group, *P < 0. 05, **P < 0.01.
As shown in Figure 4D, E, F, and G, compared with the CON group, the serum levels of SOD, GSH, and CAT in ALI group were significantly lower (P < 0.01), and the level of MDA was also significantly higher (P < 0.01). Compared with the ALI group, L-FGE group significantly elevated serum levels of GSH and CAT (P < 0.05) and significantly lowered the serum level of MDA (P < 0.05). M-FGE and H-FGE groups exhibited greatly increased serum levels of SOD, GSH, and CAT (P < 0.01), and serum levels of MDA were much lower (P < 0.01). The BIF group was consistent with the H-FGE group.
The results showed that FGE could significantly reduce the serum ALT, AST, and T-BIL levels. Meanwhile, FGE inhibited the CCl4-induced Reactive Oxygen Species (ROS) levels by increasing hepatic GSH levels and antioxidant enzyme (CAT, SOD) activity and reduced CCl4-induced lipid peroxidation by reducing MDA levels, which suggested that FGE had the effect of preserving the liver and potential defense mechanism to scavenge free radicals and relieve oxidative stress.
Effect of FGE on inflammatory factors in liver tissue of mice
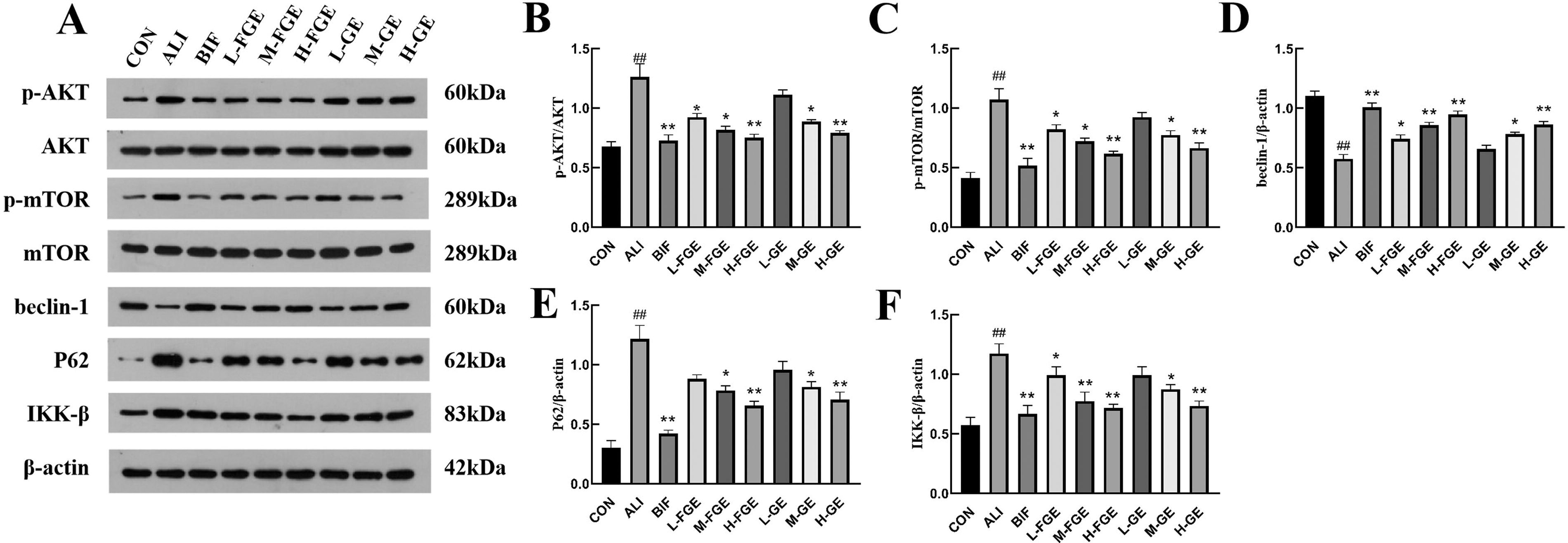
As shown in Figure 4H, I, and J, compared with the CON group, the serum levels of IL-1β, IL-6, and TNF-α in ALI group were significantly increased (P < 0.01). L-FGE group could significantly reduce the serum level of IL-1β (P < 0.05). M-FGE and H-FGE groups could extremely significantly reduce the serum levels of IL-1β, IL-6, and TNF-α (P < 0.01). The BIF group was consistent with the H-FGE group. As shown in Figure 6A and F, compared with the CON group, IKK-β levels in ALI group were significantly increased (P < 0.01). The GE and FGE groups gradually decreased IKK-β levels. Among them, IKK-β protein expressions in H-GE, H-FGE, and BIF groups were significantly decreased (P < 0.01).
Analysis of immune protein results.
The abnormal metabolic pathway of unsaturated fatty acids could activate the transcription and expression of inflammatory factors IL-1β, IL-6, and TNF-α and increase the phosphorylation level of NF-κB and induce an inflammatory response. 17 AKT1 was a subtype of serine/threonine kinase, and abnormal tyrosine metabolism regulates the activity of serine/threonine kinase, leading to the release of inflammatory cytokines. 18 IKK-β was a key protein factor on the NF-κB signaling pathway. When cells received external stimulation, it could activate IKK-β, and IKK-β catalyzed terminal serine phosphorylation and initiated the inflammatory response and downstream pathways. Thus, inhibition of IKK-β activation alleviated inflammatory responses. 19 Overall, it was suggested that FGE may suppress inflammatory factor levels by improving the metabolism of unsaturated fatty acids and amino acids.
Bioinformatics analysis
As shown in Figure 5A, the 203 common targets for FGE and ALI were obtained by Venn analysis, then these 203 common targets were analyzed using STRING and Cytoscape software to obtain the PPI network, and the darker the color, the larger the area, indicating a higher degree value for the nodes. The results are shown in Figure 5B, and AKT1, mTOR, EGFR, GRB2, HSP90AA1, and PIK3CB were the core targets of FGE to alleviate ALI of mice. A total of 617 entries of GO analysis were obtained through the metascape database, and the top 10 entries in Biological Process, Cell Conponent and Molecular Function (BP, CC, and MF) were selected for visualization as shown in Figure 5C. KEGG analysis generated 174 signaling pathways, with the top 20 enriched pathways as shown in Figure 5D, including PI3K/AKT/mTOR signaling pathway, calcium signaling pathway, and FoxO signaling pathway. The PI3K/AKT signaling pathway could inhibit autophagy, through the activation of mTOR. 20,21 Inhibition of the AKT/mTOR pathway promoted the mTOR-mediated autophagy, thereby preventing liver injury. 22,23 NAADP-mediated Ca2+ signaling was an important mediator to promote autophagy, and NAADP protected the liver from autophagy by mediating Ca2+ CD38/NAADP signaling to initiate autophagy processed and induced autophagy-related gene expression. 24 Signaling mediated by the FoxO family relieved oxidative stress and apoptosis, thereby regulating the liver, and Wnt/β-catenin signaling provided hepatocyte protection by inhibiting FoxO3. 25

Screening of key targets to construct PPI networks and analyze biological processes and enrichment pathways of important targets.
FGE inhibits CCl4-induced ALI through the AKT/mTOR signaling pathway
Bioinformatics analysis showed that the AKT/mTOR signaling pathway played a crucial role in the treatment of ALI by FGE. This pathway mediated autophagy, 26 and Western blot was used to verify the expression of related proteins, as shown in Figure 6A.
As shown in Figure 6B ,C and Table 5, compared with the CON group, p-AKT/AKT and p-mTOR/mTOR ratios in the ALI group were greatly increased (P < 0.01). The GE and FGE groups gradually decreased the phosphorylation level. The H-FGE group was superior to both L-FGE and M-FGE groups for reducing phosphorylation, as well as decreasing p-AKT/AKT and p-mTOR/mTOR ratios. The expressions of autophagy-related proteins beclin-1 and P62 were measured as shown in Figure 6D and E. The GE and FGE groups prompted increased beclin-1 protein expression and decreased P62 expression in mouse liver tissues. Among them, beclin-1 protein expression in H-GE, H-FGE, and BIF groups was significantly increased (P < 0.01), and P62 expression was significantly decreased (P < 0.01). The results showed that GE and FGE could intervene in AKT/mTOR signaling pathway to alleviate ALI by inducing the occurrence of autophagy in liver tissues and had a positive correlation with the administered dose.
Quantitative Values of Each Band in the Western Blot Data
It has been shown that inhibiting the activation of mTOR, a downstream target of the AKT/mTOR signaling pathway, contributes to the initiation of autophagy. 6,27 Autophagy is a survival mechanism that promotes the maintenance of cellular homeostasis through degradation of long-lived proteins and damaged organelles and is involved in many human diseases and physiological processes. 28 The expression amount of p-AKT and p-mTOR proteins is critical to reflect the activation of the AKT/mTOR signaling pathway. The beclin-1 protein was able to promote the formation of autophagosomes and was a key marker protein reflecting the level of autophagy. However, the increases in beclin-1 protein could be due to enhanced autophagic activity or an obstruction of the autophagic process, which could be distinguished by detecting P62 protein. While P62 was a substrate for autophagic degradation, downregulation of P62 expression indicates increased autophagy and upregulation of P62 expression indicates autophagy arrest. The results showed that CCl4 promoted decreased beclin-1 protein expression and elevated P62 expression, indicating that cellular autophagy in the tissues was inhibited. The GE and FGE groups exhibited reduced P62 protein expression, indicating their ability to induce cellular autophagy. In conclusion, the GE and FGE groups experienced enhanced autophagy by inhibiting Akt/mTOR signaling pathway, thus alleviating liver tissue damage. Moreover, the FGE group was significantly better than the GE group at the same dose, indicating a better pharmacological activity of FGE.
CONCLUSION
The results showed that FGE fermented by the F6 strain could alleviate ALI caused by CCl4, reduce inflammatory infiltration, and relieve oxidative stress and increase antioxidative enzyme markers in a concentration-dependent manner, and the greater the dose the more significant the protective effect. The mechanism of the suggested hepatoprotective effect of FGE, as assessed by Western blot detection, may be the inhibition of AKT/mTOR signaling to enhance autophagy, thereby alleviating liver tissue damage and exerting a protective effect in ALI mice. This study provides new insights into the application of natural drugs for the prevention or treatment of ALI.
Footnotes
AUTHORS’ CONTRIBUTIONS
Conceptualization, S.LN., Y.B., and H.W.; methodology, S.LN. and H.W.; software, S.LN., and P.Y.; investigation, S.LW. and L.W.; validation, S.LW.; data curation, S.LN.; visualization, S.LN. and H.W.; formal parsing, B.X. and X.S.; writing—original draft preparation, S.LN. and H.W.; writing—review and editing, Y.B.; supervision, L.Y.; and funding acquisition, Y.B. All authors have read and agreed to the published version of the article.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This research work was supported by the Science and Technology Development Plan Project of Jilin Province (No.