
Abstract
We aimed to determine the in vitro and in vivo synergistic antiallergic effect of guaijaverin and epigallocatechin gallate (EGCG) complex (GEC), and the antiallergic rhinitis (AR) properties of guaijaverin-rich Psidium guajava and EGCG-rich Camellia sinensis (ILS-F-2301). GEC showed synergistic inhibition of β-hexosaminidase by 4.20% and interleukin (IL)-4, -5, and -13 by 4.08%, 0.67%, and 4.71%, respectively, while increasing interferon (IFN)–γ by 12.43%, compared with EGCG only. In addition, 50 μg/mL of ILS-F-2301 inhibited β-hexosaminidase release, and inhibited IL-4, -5, and -13 by 61.54%, 58.79%, and 59.25%, respectively, while increasing IFN–γ (showing 133.14% activation). Moreover, 50 μg/mL of ILS-F-2301 suppressed p-STAT6 and GATA3, while p-STAT1 and T-bet increased, and 0.039 μg/mL of guaijaverin or 5.275 μg/mL of EGCG modulated T helper (Th)1- and Th2-related proteins. These data suggested that guaijaverin and EGCG in ILS-F-2301 was the main active compound involved in Th1/Th2 modulation. In the AR mouse model, the administration of ILS–F–2301 inhibited ovalbumin (OVA)-specific IgE, histamine in serum; it also inhibited IL-4 and -5 by 28.23% and 47.15%, respectively, while increasing IFN–γ (showing 37.11% activation), compared with OVA/Alu-treated mice. Taken together, our findings suggest that ILS–F–2301 is a functional food for alleviating anti-AR.
INTRODUCTION
Allergic rhinitis (AR) is an allergen-mediated disease that induces symptoms such as convulsive and repetitive sneezing, runny nose, nasal mucus, and nasal itching. 1 It has been reported that more than 40% of the population has suffered from or currently suffers from AR. The causes include the worsening environmental and atmospheric pollution. AR is a common disease worldwide and appears to be increasing in prevalence due to a host of reasons. In particular, house dust mites, dead cells from animal skin, and cockroach secretions are known to cause allergic symptoms. 2,3
To treat AR, drugs such as antihistamines, 4 leukotriene receptor antagonists, 5 and intranasal steroids plus antihistamine H-1 are used, 6 whereas in severe AR, surgery can be considered. 1 Allergies are symptoms caused by the excessive secretion of histamine in the body, and can be treated by inhaling or orally administering antihistamines. However, because these drug therapies have the disadvantage of not being fundamental treatments, methods for preventing and improving AR before it develops are required.
Guaijaverin is well known for its antiplaque 7 and antidiabetic activity, 8 while its antiallergic activity and mechanisms remain unclear. Moreover, epigallocatechin gallate (EGCG) is well known for its antiallergic effect; however, the synergistic antiallergic reaction and mechanism of guaijaverin and EGCG combination has not yet been studied. Kim et al. (2007) revealed that the mixture of Psidium guajava, Camellia sinensis, and Rosa hybrida successfully identified with ellagic acid, EGCG, and gallic acid, respectively, and they revealed the antiallergic effect of mixture extract. 9 However, Kim’s research did not reveal enough about anti-AR mechanism of the extract mixture. Therefore, this study evaluated the synergistic antiallergy improvement and/or therapeutic effects of guaijaverin-rich P. guajava and EGCG-rich C. sinensis extract mixture (ILS-F-2301) based on guaijaverin and EGCG complex (GEC) and their functional food application.
Guaijava (P. guajava), belonging to the Myrtaceae family, is a unique and traditional plant that is grown due to its diverse medicinal and nutritive properties. In particular, P. guajava leaf is used for its antispasmodic, 10 anti-inflammatory, 11 antidiarrheic, 10 antihypertensive, 12 antiobesity, 13 and antidiabetic effects. 14 –16 These effects are likely due to guaijaverin, a compound in the P. guajava leaf that possesses biological activities. Beidokhti et al. (2020) and Zhu et al. (2020) reported that guaijaverin isolated from P. guajava leave was associated with significant improvement in the function of pancreatic β-cells in diabetic mice. 13,17 Also, guaijaverin isolated from P. guajava leaf extract suppressed blood glucose homeostasis-related enzyme activity. 18
C. sinensis has anticariogenic 19 and antioxidant activity 20 with EGCG as a major compound. Gupta et al. (2023) revealed that EGCG decreased AR symptoms in ovalbumin (OVA)-treated mice and reduced Th2 cytokines (e.g., interleukin [IL]-4, -5, -13, and interferon-gamma [IFN-γ]). 21
Our study is the first to investigate the synergistic antiallergic effect of GEC. Also, in this study, we selected a mixture ratio of P. guajava and C. sinensis by checking the in vitro and in vivo synergistic antiallergic and anti-AR effects of ILS-F-2301.
MATERIALS AND METHODS
Materials
Dulbecco’s modified Eagle medium (DMEM), fetal bovine serum (FBS), minimum essential medium (MEM), and penicillin–streptomycin were obtained from Welgene (Kyongsan-si, Republic of Korea). HPLC analyses were obtained from J.T Baker and were HPLC Grade Solvent (Mallinckrodt Baker, Inc., Phillipsburg, NJ, USA). Standard compounds, avicularin (purity ≥98%), caffeine (purity ≥98%), ellagic acid (purity ≥98%), guaijaverin (purity ≥98%), epicatechin gallate (ECG, purity ≥98%), EGCG (purity ≥95%), and quercetin (purity ≥98%), were purchased from Chemfaces (Wuhan, Hubei, China). All other reagents used in quantitative analysis were of analytical grade. Aluminum hydroxide (Alu), 2,4-dinitrophenyl-human serum albumin conjugate (DNP–HSA), DNP–immunoglobulin E (DNP–IgE), ionomycin, p-nitrophenyl-N-acetyl-β-D-glucosaminide (p-NAG), methanol, ethanol, phorbol myristate acetate (PMA), sodium carbonate, sodium citrate, and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazilium reagent were purchased from Sigma–Aldrich (St. Louis, MO, USA). The antibodies to phospho–signal transducer and activator of transcription 1 (p-STAT1, #9167), T-box expressed in T cells (T-bet, #97135), p-STAT6 (#56554), GATA binding protein 3 (GATA3, #5852), β-actin (#8457), antirabbit secondary antibody (#7074), and antimouse secondary antibody (#7076) were purchased from Cell Signaling Technology (Danvers, MA, USA).
Preparation of the ILS-F-2301
P. guajava L. (Indonesia, autumn harvest) and C. sinensis (Ha-dong, Republic of Korea, autumn harvest) were extracted with 50% ethanol under reflux at 70 ± 2°C for 3 h, followed by filtration using 50 μm filter, concentration at 50°C for 3 h, and drying at 85°C for 12–24 h. Each P. guajava or C. sinensis extract powder homogenate could pass through 40 mesh. The ILS-F-2301 was prepared by mixing P. guajava L. and C. sinensis extract powders in a ratio of 2:8, and was stored at −20°C until use. We selected the 2:8 of mixture ratio of P. guajava and C. sinensis according to synergistic antiallergic activities of P. guajava and C. sinensis extract in DNP–HSA/DNP–IgE-sensitized RBL-2H3 mast cells according to various mixture ratios.
HPLC analysis
The ILS-F-2301 (0.5 g) was dissolved in 50 mL of 50% ethanol by ultrasound (Power Sonic 520, Hwashin Co., Daegu, Korea) for 20 min. The sample solution was passed through a 0.45-μm membrane filter (Advantec, Tokyo, Japan) made from polytetrafluoroethylene and injected into an analytical HPLC. HPLC was performed on an Agilent 1260 Infinity II system (Agilent Technologies, California, USA) equipped with a diode array detector (DAD) at 254 nm (guaijaverin) and 280 nm (EGCG) and an autosampler. HPLC analysis was carried out using a YMC-Triart C18 column (4.6 mm i.d. × 250 mm, YMC Inc.). The separation was conducted using a linear gradient of two solvent systems (solvent A, 0.1% formic acid in H2O, and solvent B, methanol) at a flow rate of 0.8 mL/min. Guaijaverin and EGCG were identified by comparisons of their retention times with those of the purchased standards. Linear correlation coefficients were superior to 0.990 for each compound. Contents were expressed in mg per g of the extract weight of the ILS-F-2301.
Cell culture
EL-4 cells (mouse T cell lymphocyte) and RBL-2H3 mast cells were obtained from the Korean Cell Line Bank (Seoul, Korea) and were cultured in DMEM supplemented with 10% FBS and 1% penicillin–streptomycin in an incubator with 5% CO2 at 37°C. Also, RBL-2H3 cells were cultured in MEM supplemented with 10% FBS and 1% penicillin–streptomycin in an incubator with 5% CO2 at 37°C.
Cell viability
RBL-2H3 mast cells (1 × 105 cells/well) and EL-4 T lymphocytes (2 × 105 cells/well) were seeded into 24-well plates at 37°C for 24 h, and then the ILS-F-2301 (50, 100, and 250 μg/mL), guaijaverin (0.039, 0.25, 0.5, and 1 μg/mL), EGCG (0.5, 1, and 5.275 μg/mL), quercetin (0.012 μg/mL), ellagic acid (0.014 μg/mL), avicularin (0.007 μg/mL), caffeine (1.8 μg/mL), ECG (0.46 μg/mL), and GEC (mixture of 0.039 μg/mL of guaijaverin and 5.275 μg/mL of EGCG) were added to the wells at 37°C for 24 h. After 24 h incubation, WST-8 reagent (ab228554, Abcam Biotechnology, MA, USA) was added and the plates were then incubated at 37°C for 4 h. After 4 h incubation, cell viability was measured using an ELISA reader at 550 nm wavelength. Viability was expressed as the percentage of live cells in each well.
β-hexosaminidase release
Briefly, RBL-2H3 cells (1 × 105 cells/well) were harvested by trypsinization, transferred to 24-well plates (1 × 105 cells/well), and incubated overnight with DNP–IgE (50 ng/mL). After 24 h incubation, the cells were washed twice with MEM medium containing 1% FBS, and then added to MEM at 37°C for 20 min. After 20 min incubation, the ILS-F-2301 (50, 100, and 250 μg/mL), guaijaverin (0.039, 0.25, 0.5, and 1 μg/mL), EGCG (0.5, 1, and 5.275 μg/mL), quercetin (0.012 μg/mL), ellagic acid (0.014 μg/mL), avicularin (0.007 μg/mL), caffeine (1.8 μg/mL), ECG (0.46 μg/mL), and GEC (mixture of 0.039 μg/mL of guaijaverin and 5.275 μg/mL of EGCG) were added to each well at 37°C for 1 h. After 1 h incubation, DNP–HSA (100 ng/mL) was added for 4 h at 37°C. The supernatant was then centrifuged (13,000 rpm, 10 min), and an appropriate amount of the resulting supernatant was reacted with 10 mM of p-NAG (with 100 mM sodium citrate buffer, pH 4.5) at 37°C for 1 h. After 1 h, the stop solution (100 mM sodium carbonate buffer, pH 10.0) was added to each tube. The β-hexosaminidase release was measured with an ELISA reader at 405 nm.
ELISA
Supernatants from RBL-2H3 mast cells were analyzed for the IL-4, -5, -13, and IFN-γ concentrations by ELISA (Abcam biotechnology, MA, USA) kits according to the manufacturer’s instructions.
Modulatory effect of GEC or ILS-F-2301
Briefly, EL-4 T lymphocytes (2 × 105 cells/well) were seeded into a 24-well cell culture plate, and then incubated for 24 h at 37°C. After 24 h of incubation, the ILS-F-2301 (50, 100, and 250 μg/mL), guaijaverin (0.039, 0.25, 0.5, and 1 μg/mL), EGCG (0.5, 1, and 5.275 μg/mL), or GEC (mixture of 0.039 μg/mL of guaijaverin and 5.275 μg/mL of EGCG) plus OVA (200 μg/mL) were added to each well for 24 h. After 24 h of incubation, the supernatants from EL-4 T lymphocytes were collected and then the IL-4 and IFN-γ concentrations were determined by ELISA (Abcam Biotechnology, MA, USA) according to the manufacturer’s manuals. The supernatants were stored at −70°C until further analysis.
Western blot analysis
Briefly, EL-4 cells (2 × 105 cells/well) were seeded into a 6-well cell culture plate, and incubated for 24 h at 37°C. After 24 h incubation, the ILS-F-2301 (50, 100, and 250 μg/mL), guaijaverin (1 μg/mL), EGCG (1 μg/mL) plus OVA (200 μg/mL) were added to each well at 37°C for 24 h. EL-4 cells were harvested and lysed with RIPA buffer containing phosphatase/protease cocktails. In addition, the collected nasal tissues were lysed using tissue extraction buffer. Protein concentrations of samples were estimated by the BCA protein assay (Gendepot, Barker, TX, USA) prior to the addition of loading buffer. Samples were then heated at 100°C for 5 min and separated at 20 μg of protein/lane by 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis prior to electro-transfer to polyvinylidene fluoride (PVDF) membranes. After blocking with 5% skim milk for 1 h, the PVDF membranes were incubated overnight at 4°C with primary antibodies against p-STAT1, T-bet, p-STAT6, GATA3, and β-actin. Following incubation with secondary antibodies, signals were detected using enhanced chemiluminescence (ThermoFisher Scientific, Rockford, IL, USA).
Animals
Six-week-old male BALB/c mice were purchased from Hana Bio (Gyeonggi-do, Republic of Korea). The protocol for animal use in this study was reviewed and approved by the SEMI Institutional Animal Care and Use Committee (SEMI-23-011). The mice were housed under controlled temperature of 23 ± 3°C with relative humidity of 40–60% and 12 h light/dark cycles. Food and water were provided ad libitum.
Sensitization and challenges
The in vivo anti-AR model was based on a modification of a previous protocol. 22,23 The BALB/c mice were divided into five groups (seven mice/group): (1) normal group—no sensitization; (2) control group—sensitization with OVA (50 μg) and Alu (2 mg); (3) low dosage of ILS-F-2301 group—sensitization with OVA and Alu plus 100 mg/kg of ILS-F-2301 by oral administration (per os; p.o.); (4) middle dosage of ILS-F-2301 group—sensitization with OVA and Alu plus 150 mg/kg of ILS-F-2301 by oral administration (p.o.); and (5) high dosage of ILS-F-2301 group—sensitization with OVA and Alu plus 200 mg/kg of ILS-F-2301 by oral administration (p.o.).
Briefly, after mice were acclimated for 1 week, they were OVA-sensitized by intraperitoneal injection of 50 μg OVA in 200 μL phosphate buffered saline containing 2 mg alum on days 14 and 28 (Fig. 1). Sensitization was followed by daily intranasal challenges for 7 days with OVA/Alu. The mice were sacrificed one day after the last challenge on day 35. The mice were treated with ILS-F-2301 at 100, 150, and 200 mg/kg dosing by p.o. from days 7 to 35. The mice in the control and OVA/Alu groups were administered saline by p.o. At the end of 4 weeks, all mice were fasted overnight and then euthanized with CO2.

Antiallergic rhinitis animal design of ILS-F-2301.
Plasma analysis
The plasma was collected from sacrificed mice blood. The production of OVA-specific IgE, histamine, IL-4, -5, -13, and IFN-γ were measured using ELISA kits (Abcam, CA, USA) according to the manufacturer’s protocols.
Flow cytometric analysis of T helper 1 and T helper 2 cells
A single cell suspension was prepared by passing and disrupting spleens through a cell strainer (BD Biosciences, Durham, NC, USA). Briefly, the cells were stimulated with PMA/ionomycin before staining with antibodies. Next, the cells were labeled with fluorescently conjugated antimouse CD3 (fluorescein isothiocyanate) and CD4 (phycoerythrin; PE) antibodies (BD Biosciences, San Jose, CA, USA) at 4°C for 30 min. Following surface staining, cells were fixed and permeabilized in perm/wash buffer for 20 min at 4°C in the dark, then incubated with conjugated anti-IL-4 (allophycocyanin; APC) and anti-IFN-γ (APC) antibody (BD Biosciences) in each tube for 30 min at 4°C in the dark. After fixing with 1% paraformaldehyde, the stained cells were detected by flow cytometry (BD Biosciences).
Statistical analysis
Statistical analyses were performed using SPSS v21.0 software (SPSS, Chicago, IL, USA). Data are shown as means ± standard errors of the means (SEM) from three independent experiments in vitro and seven independent experiments in vivo, unless stated otherwise. Differences between treatment groups were evaluated using one-way analysis of variance (ANOVA) followed by Tukey–Kramer test and P < .05 was considered significant.
RESULTS
Selection of the active compound from P. guajava or C. sinensis extract
We successfully detected the main compounds isolated from P. guajava or C. sinensis using HPLC-DAD. The major compound from P. guajava was ellagic acid (RT = 52 min, 5.51 mg/g of P. guajava extract), and that from C. sinensis was EGCG (RT = 23 min, 133.2 mg/g of C. sinensis extract) (Fig. 2). The other main compounds from P. guajava were quercetin (RT = 38 min, 4.65 mg/g of P. guajava extract), guaijaverin (RT = 53.5 min, 4.12 mg/g of P. guajava extract), and avicularin (RT = 57 min, 2.94 mg/g of P. guajava extract), and those from C. sinensis were caffeine (RT = 21 min, 45.1 mg/g of C. sinensis extract) and ECG (RT = 31.6 min, 11.4 mg/g of C. sinensis extract).
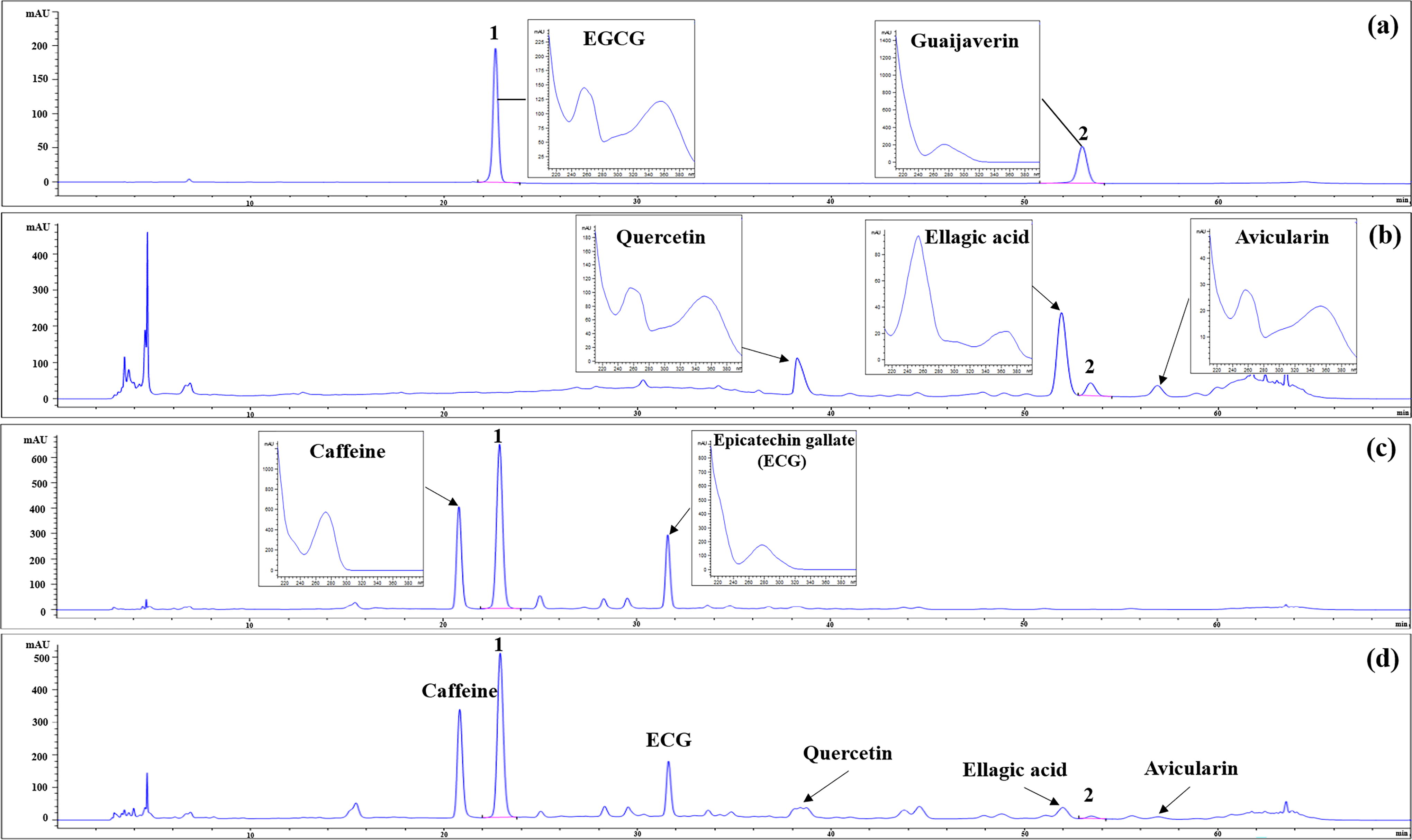
HPLC chromatograms of (a) two standard solutions; 50% ethanol extract of (b) Psidium guajava, (c) Camellia sinensis, and (d) ILS-F-2301. 1: (-)-epigallocatechin gallate; 2: guaijaverin.
Next, to check the antiallergic efficacy of the detected compounds in P. guajava or C. sinensis, we investigated the inhibition of the β–hexosaminidase release of quercetin, ellagic acid, guaijaverin, and avicularin in P. guajava and of caffeine, EGCG, or ECG in C. sinensis. As a result, there was no cytotoxicity of quercetin, ellagic acid, guaijaverin, avicularin, caffeine, EGCG, or ECG (Fig. 3a). Also, guaijaverin inhibited β-hexosaminidase release by 42.8% compared with the DNP–HSA/DNP–IgE-sensitized mast cells, respectively (Fig. 3b). In addition, caffeine, EGCG, and ECG decreased the β–hexosaminidase release by 10.1%, 34.4%, and 14.5% compared with the DNP–HSA/DNP–IgE-treated mast cells. These data suggest that guaijaverin and EGCG from P. guajava and C. sinensis had high-efficacy. Therefore, we selected guaijaverin and EGCG as active compounds for their antiallergic activity. The contents of guaijaverin and EGCG of ILS-F-2301 were 0.78 ± 0.01 and 105.50 ± 1.00 mg/g, respectively.
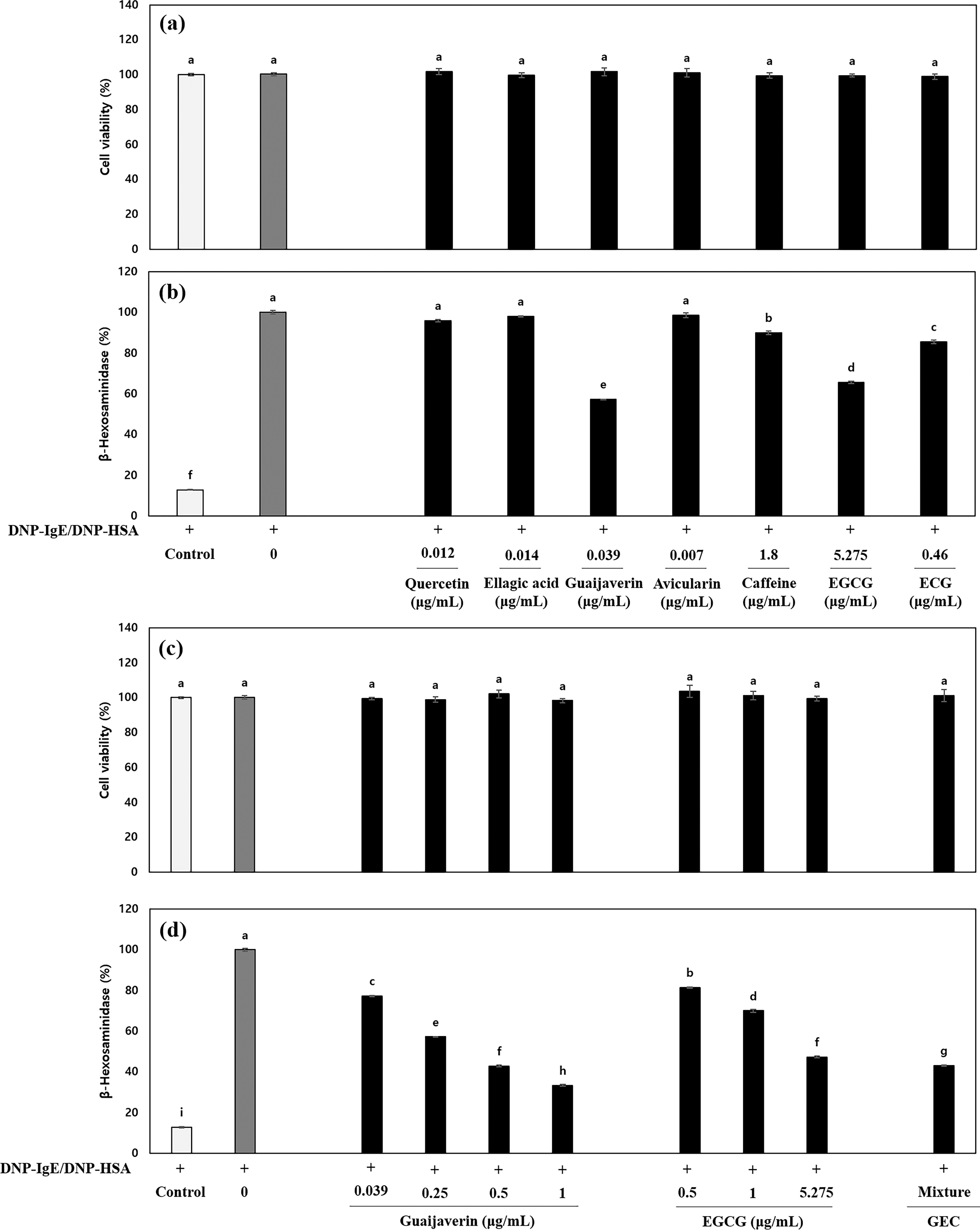
Antiallergic and additive effects of detected compounds in DNP–HSA/DNP–IgE-treated RBL-2H3 mast cells.
Synergistic antiallergic effect of GEC in DNP–HSA/DNP–IgE-treated RBL-2H3 cells
To investigate the synergistic antiallergic effect of GEC, we analyzed the β-hexosaminidase release and Th2-related cytokine secretion in response to GEC in the DNP–HSA/DNP–IgE-treated RBL-2H3 mast cells. The results show there was no cytotoxicity at any concentration of guaijaverin, EGCG, or GEC (Fig. 3c). In addition, GEC showed synergistic inhibition of β-hexosaminidase release by 4.20% compared with 5.275 μg/mL of EGCG (Fig. 3d).
Also, the increases of IL-4, -5, and -13 and the decrease in IFN-γ were synergistically inhibited by 4.08%, 0.67%, 4.71%, and increased by 12.43% by GEC, respectively, compared with the EGCG treated RBL-2H3 cells, indicating that guaijaverin and EGCG play key roles in the anti-allergenic effect through the modulation of T helper (Th)1 and Th2 cytokines (Table 1).
Th1 and Th2 Cytokine Responses of Guaijaverin, Epigallocatechin Gallate or Guaijaverin Complex in DNP–IgE/DNP–HSA-Mediated RBL-2H3 Mast Cells
Data represent mean ± SD of triplicate determinations. Differences between treatment groups were evaluated using one-way ANOVA followed by Tukey–Kramer test and P < .05 significantly different from DNP–HSA/DNP–IgE-treated cells by one-way ANOVA. GEC is a combination of 0.039 μg/mL of guaijaverin and 5.275 μg/mL of EGCG corresponding to 50 μg/mL of ILS-F-2301. Values with different superscript (a-j) letters are significantly different among groups by ANOVA with Tukey–Kramer test at P < .05.
Anti-allergic efficacy of ILS-F-2301 according to the mixture ratio
To select the mixture ratio of P. guajava and C. sinensis, we investigated the β-hexosaminidase release using the DNP–HSA/DNP–IgE-sensitized RBL-2H3 mast cells. In our data, there was no cytotoxicity at any concentration of P. guajava and C. sinensis mixture or of P. guajava and C. sinensis extract (Fig. 4a). In addition, the 5:5, 4:6, 3:7, and 2:8 mixture ratios of P. guajava and C. sinensis significantly inhibited the β-hexosaminidase release by 72.1 ± 0.43%, 75.4 ± 0.80%, 77.9 ± 0.38%, and 79.8 ± 0.48% (P < .001) at 100 μg/mL, respectively (Fig. 4b). Furthermore, the 2:8 mixture ratio of P. guajava and C. sinensis had synergistic anti-allergic efficacy of 12.3% and 5.4% compared with P. guajava alone (67.5 ± 0.22%) or C. sinensis alone (73.4 ± 0.34%), respectively. Therefore, we selected a mixture ratio of 2:8 for P. guajava and C. sinensis.
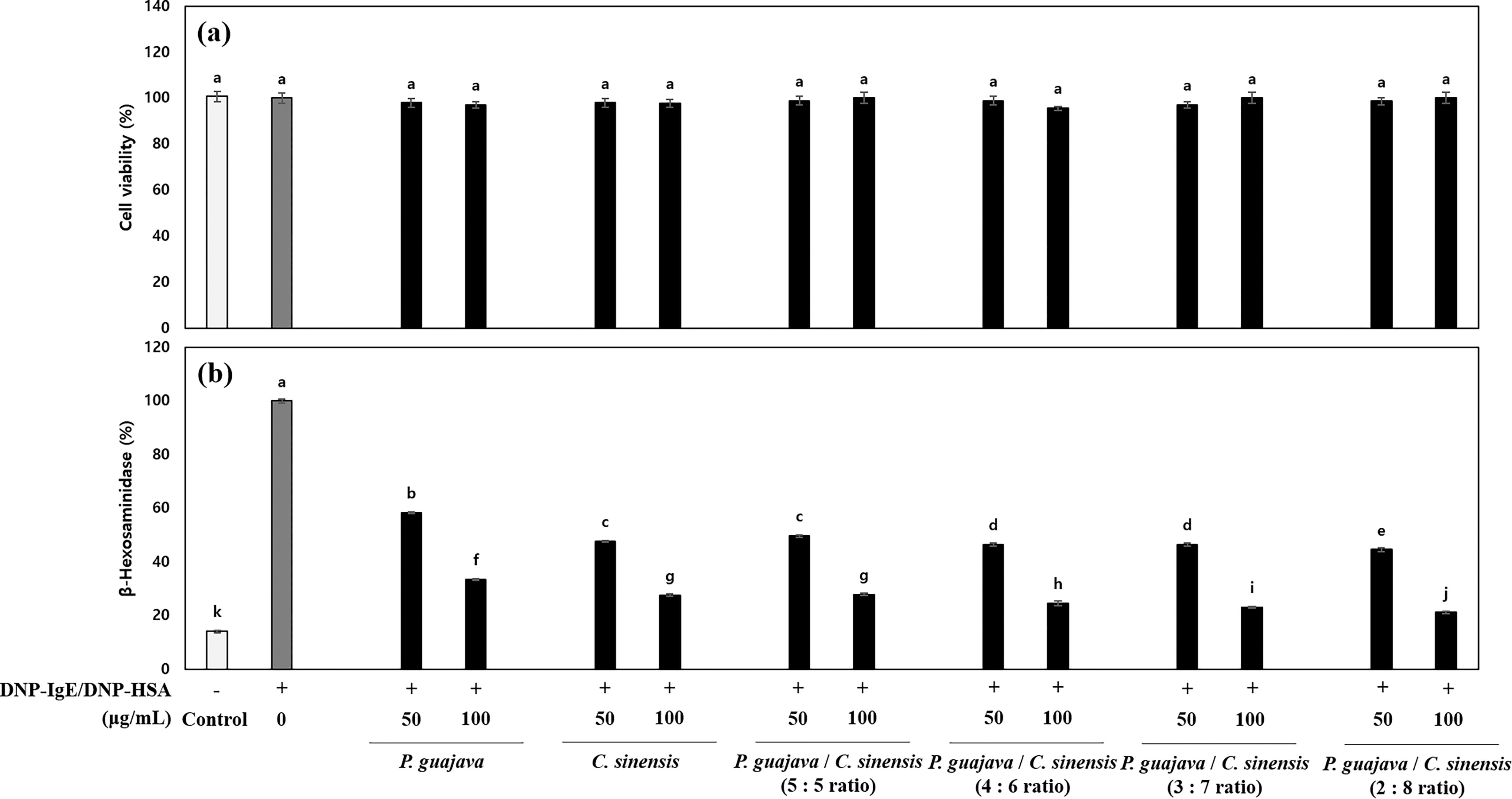
The antiallergic effect of mixture of P. guajava and C. sinensis in DNP–HSA/DNP–IgE-treated RBL-2H3 mast cells according to mixture ratio.
Antiallergic effect of the ILS–F–2301 in DNP–HSA/DNP–IgE-treated RBL–2H3 cells
Next, we examined the antiallergic effect of the ILS-F-2301 in allergy-induced mast cells using ELISA analysis. The increases in IL-4, -5, and -13 secretion by DNP–IgE/DNP–HSA were inhibited by 79.0%, 77.3%, and 83.2% respectively (P < .001) upon treatment with 100 μg/mL of ILS-F-2301 (Table 2). Furthermore, the DNP–HSA/DNP–IgE-stimulated RBL-2H3 mast cells inhibited the secretion of IFN-γ by about 0.34-fold (P < .001) compared with the untreated cells, while 100 μg/mL of the ILS-F-2301 increased IFN-γ by about 263.6% (P < .001) compared with the DNP–IgE/DNP–HSA-sensitized RBL-2H3 cells.
Th1 and Th2 Cytokine Responses of ILS-F-2301 in DNP–IgE/DNP–HSA-Mediated RBL-2H3 Mast Cells
Data represent mean ± SD of triplicates. Differences between treatment groups were evaluated using one-way ANOVA followed by Tukey–Kramer test and P < .05 significantly different from DNP–HSA/DNP–IgE-treated cells by one-way ANOVA. ILS-F-2301 expressed by the 2:8 mixture ratio of P. guajava and C. sinensis extract. Values with different superscript (a-e) letters are significantly different among groups by ANOVA with Tukey–Kramer test at P < .05.
Modulation activity of the ILS-F-2301 in OVA-treated T lymphocytes
We then examined Th1 and Th2-related cytokine secretion by ILS-F-2301 in OVA-treated T lymphocytes to investigate the modulatory activity of the ILS-F-2301. There was no cytotoxicity exhibited at any concentration of the ILS-F-2301 (Fig. 5a). The OVA-treated T lymphocytes stimulated the secretion of IL-4 while inhibiting IFN-γ secretion by about 4.6- and 0.4-fold (P < .001), respectively, compared with the untreated cells (Fig. 5b and 5c). Furthermore, with ILS-F-2301 at 100 μg/mL, OVA-treated T lymphocytes demonstrated a significant inhibition of IL-4 secretion of about 0.54-fold (P < .001), while exhibiting an increase in IFN-γ by about 1.9-fold (P < .001).
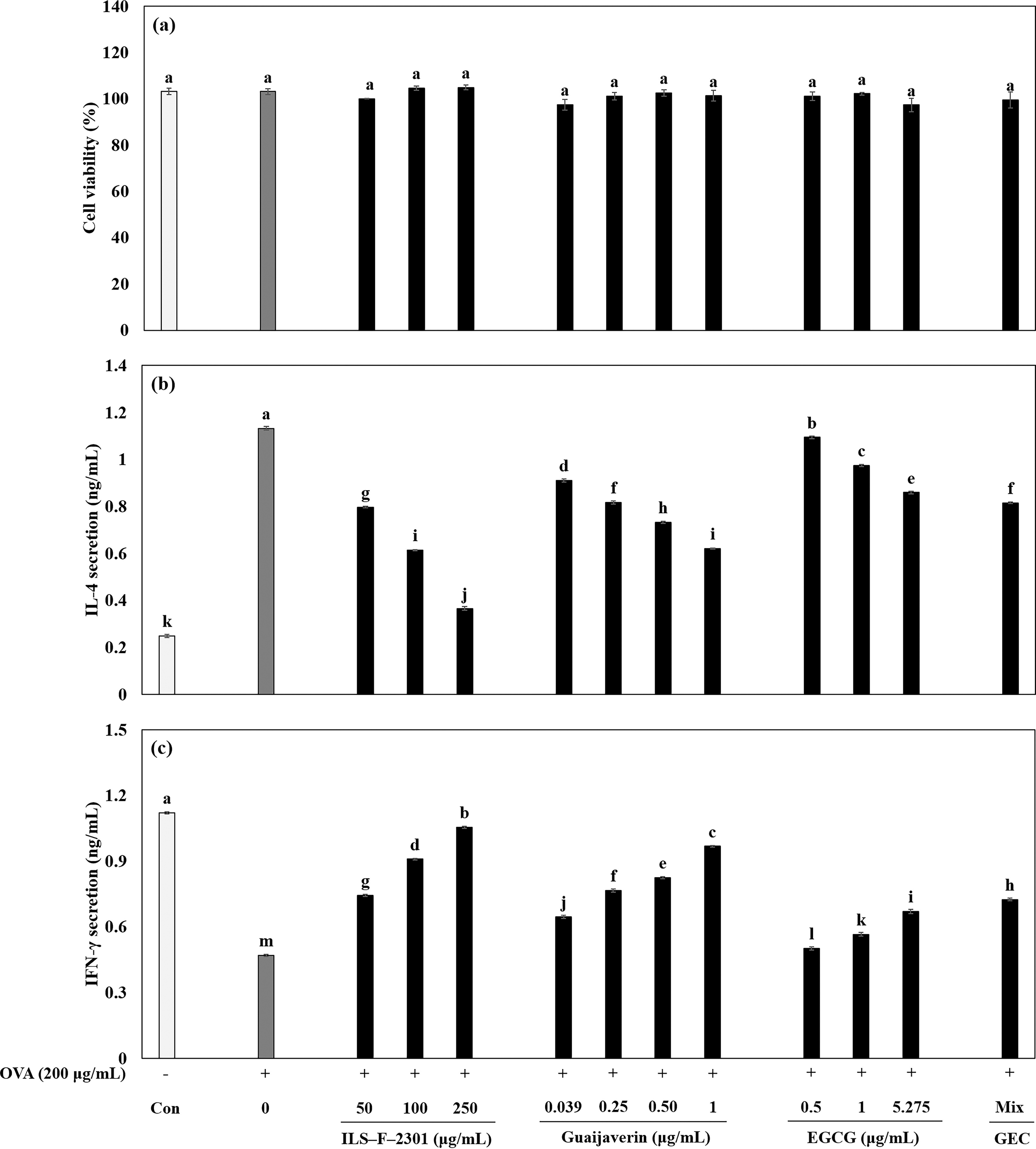
The immune-modulatory effect of ILS-F-2301 on Th1 and Th2 cytokines in OVA-treated T lymphocytes.
Th1/Th2-related protein expression of ILS-F-2301 in OVA-treated EL-4T lymphocytes
Next we examined the mechanisms of ILS-F-2301 activity using western blot analysis. The 100 μg/mL of ILS-F-2301 with the OVA-treated EL-4 T lymphocytes significantly increased p-STAT1 and T-bet by about 66.3% and 78.4%, respectively (P < .001) compared with the OVA-treated EL-4 T lymphocytes (Fig. 6, lane 2 and 4). In addition, over-expressed p-STAT6 and GATA3 in response to OVA was inhibited by 100 μg/mL ILS-F-2301 by about 118.3% and 141.6%, respectively (P < .001), compared with the OVA-treated EL-4 T lymphocytes.
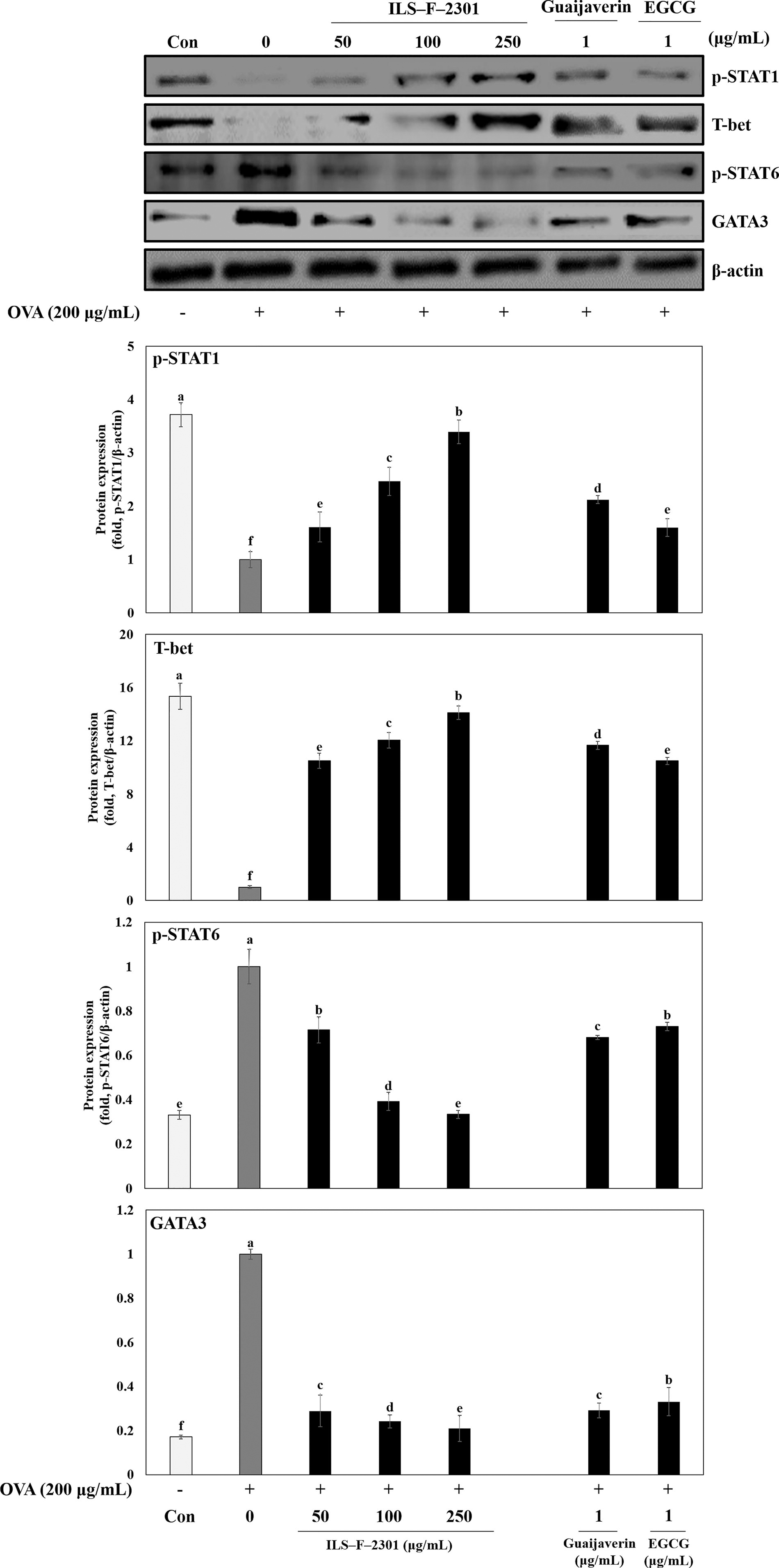
The modulatory mechanisms of guaijaverin, EGCG, or ILS-F-2301 in OVA-treated EL-4 T lymphocytes. Briefly, EL-4 T lymphocytes were seeded into 6-well cell culture plates, and then cultured for 24 h, after which ILS-F-2301 (50, 100, and 250 μg/mL) or guaijaverin or EGCG plus OVA (200 μg/mL) were added, and then further incubated for 24 h. Cell pellets were then collected, lysed and subjected to western blotting. Data are represented as the mean ± standard error of mean (SEM) from three independent experiments, unless stated otherwise. Differences between treatment groups were evaluated using one-way ANOVA followed by Tukey–Kramer test and P < .05 significantly different from OVA-treated cells, as determined by one-way ANOVA.
Furthermore, guaijaverin is increased the suppressed p-STAT1 and T-bet by OVA about 2.12-or 15.9-fold (P < .001) compared with OVA-treated cells, respectively (Fig. 6, lanes 6). Also, EGCE is increaed the inhibited p-STAT1 and T-bet by OVA about 11.7- or 10.5-fold (P <.001), respectively (Fig. 6, lane 7). Also, guaijaverin or EGCG significantly inhibited the over-expressed p-STAT6 and GATA3 by OVA by about 68.0% or 73.0% and 29.1% or 33.1%, respectively (Fig. 6, lanes 6 and 7), suggesting that guaijaverin and EGCG were the key molecules related to antiallergic activity.
Effects of the ILS-F-2301 on OVA-specific IgE and histamine in plasma
Key roles are played by IgE and histamine production in the pathophysiology of AR during the early immunological response to OVA-sensitization. 24 Therefore, we measured the OVA-specific IgE and histamine content in OVA/Alu-treated mice. The OVA/Alu treated group exhibited significantly increased concentrations of OVA-specific IgE and histamine compared with the normal group (Fig. 7a and b). The administration of ILS-F-2301 dramatically inhibited these markers (P < .001) compared with the OVA/Alu group, suggesting that the ILS-F-2301 has antiallergenic effects, especially in inhibiting OVA-mediated AR activity.

Antiallergic rhinitis effect of ILS-F-2301 in OVA/Alu-sensitized mice according to dosing levels.
The effects of ILS-F-2301 on Th1 and Th2 cytokines in plasma
Next, to investigate the modulatory effects of the ILS-F-2301 on OVA/Alu-treated mice, we assayed IL-4, -5, -13, and IFN-γ levels using ELISA. The IL-4, -5 and -13 secretion into plasma was stimulated 2.4-, 1.7-, and 2.2-fold respectively in the OVA/Alu group compared with the normal group (P < .001, Table 3). In contrast, the ILS-F-2301 at 150 mg/kg inhibited IL-4 and -5 secretion to about 48% and 43% of OVA/Alu stimulated values (P < .001); however, IL-13 secretion decreased by only 9% (P > .05), and significantly decreased at 200 mg/kg administration of the ILS-F-2301. In addition, OVA/Alu treatment significantly inhibited IFN-γ secretion to about half (P < .001) of that observed in the normal group, while ILS-F-2301 supplementation rescued IFN-γ levels (P < .001).
Th1 and Th2 Cytokines of ILS-F-2301 in OVA/Alu-Sensitized Mice
Statistical analyses were performed by using the Tukey–Kramer test and P < .05 as significantly different from OVA/Alu-sensitized mice by one-way ANOVA (n = 7). ILS-F-2301 is the mixture of P. guajava and C. sinensis extract. The ILS-F-2301 group administered 100, 150, and 200 mg/kg. Values with different superscript (a-e) letters are significantly different among groups by ANOVA with Tukey–Kramer test at P < .05.
Effects of the ILS-F-2301 on Th1 and Th2 balance
Then, we assayed the Th1 and Th2 populations and Th1/Th2 ratios using flow cytometric analysis. The balance between Th1 and Th2 cells a plays key role in the evaluation of allergic reactions. The proportion of CD4+ IL-4+ Th2 cells was increased, while CD4+ IFN-γ+ Th1 cells were significantly decreased by treatment with OVA and Alu (Fig. 8a and 8b). ILS-F-2301 treatment modulated the Th1 and Th2 cells compared with the OVA/Alu group. Furthermore, the Th1/Th2 ratio significantly increased in the 100, 150, and 200 mg/kg dosage of the ILS-F-2301 group compared with OVA/Alu alone (Fig. 8c).

Effect of ILS-F-2301 on Th1 and Th2 cell populations in OVA-sensitized mice according to dosing levels.
DISCUSSION
Guaijaverin has efficacy for antiplaque 7 and antidiabetic activity, 8 while its antiallergenic effects have not been previously studied. In a similar study, Kim et al. (2007) reported the antiallergic effects of P. guajava, C. sinensis, and R. hybrida; however, they only performed an individual analysis method of mixture extract. The anti-AR mechanisms of active compounds and interactions of their mixtures have not yet been clarified. 9 Our study is the first to investigate the synergistic antiallergic mechanism of GEC and we examined the anti-AR effect of guaijaverin-rich P. guajava and EGCG-rich C. sinensis mixture extract, ILS-F-2301, as an application example.
In our data, we successfully identified four major compounds (quercetin, ellagic acid, guaijaverin, and avicularin) in P. guajava and three major compounds (caffeine, EGCG, and ECG) in C. sinensis (Fig. 2). Among these compounds, we selected guaijaverin and EGCG in the ILS-F-2301 based on the biological activities and contents of the ILS-F-2301. Figure 3b shows that there was no antiallergy efficacy for quercetin, ellagic acid, and avicularin in the DNP–HSA/DNP–IgE-mediated RBL-2H3 mast cells. Also, among the three active compounds in C. sinensis, EGCG effectively inhibited β-hexosaminidase release compared with caffein or ECG. Nugrahini et al. (2019) revealed that caffeine inhibited the FcεRI-mediated intracellular signaling pathway through suppressing the phosphorylation of α-synuclein (Syn), bruton tyrosine kinase (Btk), phosphoinositide 3-kinase (PI3K), and A serine/threonine protein kinase (Akt). 25 In addition, ECG effectively suppressed FcεRI expression. 26
In our in vitro experiments, we examined the effects of the synergistic antiallergic effect of GEC and its application, ILS–F–2301, in allergy-stimulated cells. Mast cells stimulated by IgE mainly produced Th2 cytokines such as IL-4, -5, and -13. According to Hwang’s report (2018), Aster yomena extract statistically reduced the increase of IL-4, -5, and -13 secretion in stimulated RBL-2H3 mast cells. 22 In addition, Lee et al. (2020) revealed that increased IL-4, -5, and -13 secretion by IgE stimulation was reduced on exposure to enzyme-treated Zizania latifolia extract. 27 These reports mainly evaluated the suppression of Th2 cytokines through the stimulation of RBL-2H3 mast cells. Therefore, we focused our observations on the anti-allergic activity of GEC or ILS-F-2301 in DNP–HSA plus DNP–IgE-sensitized RBL-2H3 mast cells. Our data demonstrate that GEC had an additional antiallergic effect (Fig. 3d) compared with guaijaverin or EGCG only. By calculating the synergistic antiallergic effect, the optimal mixture ratio of P. guajava and C. sinensis extract was 2:8 (Fig. 4). In addition, guaijaverin or EGCG and ILS-F-2301 significantly inhibited Th2 cytokines, IL-4, -5, and -13, indicating that guaijaverin or EGCG in ILS-F-2301 remarkably inhibits the over-production of Th2 cytokines (Table 1 and 2). In addition to the Th1 cytokines, IFN-γ by IgE synthesis and Th2 cell differentiation are important markers for evaluating allergic activity. 28 As another indication of the efficacy of guaijaverin, EGCG, or ILS-F-2301, we showed that guaijaverin, EGCG, or ILS-F-2301 significantly increased the inhibition of IFN-γ in RBL-2H3 mast cells, suggesting that the medicinal formulation modulated the imbalance of Th1 and Th2 cytokines by IgE mediation.
Next, to probe the mechanisms underlying guaijaverin functionality more deeply as well as EGCG and its functional blend (ILS-F-2301) biological potency, we examined STAT1/T-bet and STAT6/GATA3 expression in OVA-treated EL-4 T lymphocytes. STAT1/T-bet signaling pathway is well known to modulate Th differentiation. Also, the STAT6/GATA3 signaling pathway mediates Th2 cytokines. Hwang et al. (2018) and Ren et al. (2022) showed that IL-4, IL-13, and IFN-γ production induced STAT1/T-bet and STAT6/GATA3 imbalances in an OVA-induced AR mouse model. 22,23 Also, according to Jung et al. (2023) and Li, Wang, and Han (2023), GATA3 is a major factor for Th2 cell differentiation and in vivo cytokine production in GATA3. 3,29 Therefore, we focused on Th1 and Th2 balance in T lymphocytes by OVA treatment. Also, ILS-F-2301, guaijaverin, or EGCG modulated Th1 and Th2 cytokines in OVA-treated T lymphocytes (Fig. 5b and 5c). Furthermore, guaijaverin, EGCG, or their blend, ILS-F-2301, significantly suppressed STAT6/GATA3 and reduced STAT1/T-bet proteins (Fig. 6). These findings demonstrate that the key molecules of guaijaverin or EGCG of ILS-F-2301 are related to modulation of OVA-mediated the Th1 and Th2 cytokines.
IgE is a major biomarker for AR in OVA/Alu-co-treated mice, and the OVA/Alu-sensitized mouse model has been well characterized and its utility has been established for studying AR. 20,24,30,31 It is also well understood that Th1/Th2 cytokines, IgE, and histamine are involved in clinical allergic presentations such as atopic dermatitis, 32 asthma, 33 and AR. Allen (2022) revealed that activation of B cells stimulates the release of histamine in blood, and that the hyper-secretion of histamine is caused by allergic reactions. 33 Therefore, we focused on the modulatory effects of ILS-F-2301 on OVA-specific IgE and histamine in plasma and confirmed that indeed, ILS-F-2301 administration significantly inhibited OVA-specific IgE and histamine secretion (Fig. 7a and 7b).
Cytokines IL-4 and -13 are major participants in switching recombination to IgE. 28 The ILS-F-2301 significantly inhibited the secretion of IL-4, -5, and IL-13. Also, we observed that 200 mg/kg of ILS-F-2301 significantly suppressed the secretion of IL-13 compared with the OVA/Alu group (Table 3). Furthermore, ILS-F-2301 increased the secretion of IFN-γ compared with OVA/Alu alone. In addition, we measured the populations of Th1 and Th2 cells using flow cytometric analysis. The CD4+ IFN-γ+ T (Th1) and CD4+ IL-4+ T (Th2) cell populations play key roles in modulating the activity of active compounds. Hwang et al. (2018) revealed that Aster yomena extract remarkably modulated Th1 and Th2 cells through the inhibition of Th cell activation. 22 Also, Ishida et al. 32 reported that aqueous extract from Cuminum cyminum L. seed alleviates OVA-induced AR through the balancing of Th1 and Th2 cells via the downregulation of the PI3K pathway, which is known to be involved in T cell activation and differentiation. 32 Figure 8 shows that the ILS-F-2301 group exhibited significantly increased numbers of Th1 cells and fewer Th2 cells in splenocytes, indicating that ILS-F-2301 regulates and alleviates the imbalance of Th1 and Th2 cells observed with OVA and Alu treatment.
CONCLUSIONS
We first demonstrate that te synergistic anti-allergic effect of GEC and ILS-F-2301. The comprising natural sources can serve as a functional food th ameliorate AR symptoms by modulating the allergen-mediated IL-4/STAT6/GATA3 and IFN-γ/STAT1/T-bet signaling pathways. Our finding might be expected to explain the anti-AR activity related to the guaijaverin and EGCG combination in the ILS-F-2301. We also anticipate that if adopted by the food industry, ILS-F-2301 will show efficacy in preventing or ameliorating the symptoms of AR.
Footnotes
AUTHORS’ CONTRIBUTIONS
S.-H.P.: Conceptualization (lead); investigation (equal); formal analysis (lead); supervision (equal); visualization (lead); writing—original draft (equal); Methodology (lead); formal analysis (equal). Y.-H.J.: Writing—original draft (equal); investigation (equal); formal analysis (equal); validation (lead). P.Y.J.: Writing—original draft (supporting); Software (supporting); investigation (supporting). K.J.S.: Formal analysis (supporting). L.J.B.: Project administration (lead); resources (lead); supervision (equal); writing review and editing (lead).
DATA AVAILABILITY
All data are reported in the article.
ETHICS APPROVAL STATEMENT
All the procedures were performed according to the guidelines approved by the National Institutes of Health. All animal experiments were approved by the SEMI Institutional Animal Care and Use Committee (SEMI–23–011).
AUTHOR DISCLOSURE STATEMENT
The authors declare they have no conflicts of interest.
FUNDING INFORMATION
This research received no external funding.