
Abstract
Purpose:
This study aimed to investigate the protective effects of gallic acid (GA) against ovarian damage induced by bisphenol A (BPA) exposure in female rats. We evaluated whether GA can mitigate the adverse effects of BPA on ovarian structure, inflammatory markers, oxidative stress, apoptosis, and reproductive hormone levels.
Methods:
Thirty-two female rats were categorized into four groups: control, GA, BPA, and GA+BPA. Histopathological evaluations of ovarian tissue were performed using hematoxylin-eosin staining. The immunohistochemical analysis was conducted for inflammatory, oxidative DNA damage, and apoptotic markers (Tumor necrosis factor alpha [TNFα], cyclooxygenase-2 [COX2], interleukin-1 beta [IL-1β], 8-hydroxydeoxyguanosine [8-OHdG], and caspase 3). Oxidative stress was assessed by measuring malondialdehyde and superoxide dismutase levels. Furthermore, follicle-stimulating hormone (FSH), luteinizing hormone (LH), estrogen, and progesterone levels were quantified using enzyme-linked immunosorbent assay.
Results:
Histopathological outcomes revealed that BPA significantly induced follicular degeneration, which was effectively mitigated by GA treatment (P < 0.05). Immunohistochemical analysis highlighted the exacerbation of inflammatory responses and oxidative DNA damage and apoptosis (TNFα, COX-2, IL-1β, 8-OHdG, and caspase 3) in BPA-exposed tissues, which were reduced in the presence of GA (P < 0.05). The assessment of oxidative stress demonstrated that GA could significantly decrease lipid peroxidation and partially restore antioxidant defense mechanisms disrupted by BPA (P < 0.05). Hormonal profiling indicated that BPA exposure altered the levels of FSH, LH, estrogen, and progesterone, with GA treatment showing a capacity to modulate these changes, especially in progesterone levels (P < 0.05).
Conclusions:
The findings suggest that GA exhibits protective properties against BPA-induced ovarian damage through its antioxidative and anti-inflammatory activities, alongside its ability to modulate hormonal imbalances. This research underscores the therapeutic potential of GA in safeguarding reproductive health against environmental toxicants.
INTRODUCTION
Bisphenol-A (BPA), commonly used as a monomer in manufacturing polycarbonate plastics and epoxy resins, has been recognized as an endocrine disruptor. It possesses the capacity to disrupt the regular functioning of the endocrine system. 1 –3 The ubiquity of BPA in consumer products has led to widespread human exposure, as evidenced by detectable levels in bodily fluids, raising concerns about its role in various health disorders, including reproductive dysfunctions. 4
The ovary, a pivotal organ in female reproduction, is susceptible to toxic insults because of its role in oogenesis and steroidogenesis. It is a highly dynamic structure that undergoes cyclic changes and is responsible for the maturation of oocytes and the production of sex hormones, which are essential for reproductive function. 5 Experimental studies have demonstrated that BPA exposure can lead to ovarian toxicity, characterized by disrupted folliculogenesis, oxidative stress, and altered steroid hormone levels, which may culminate in compromised fertility. 4,6 The insights are crucial for developing strategies to protect ovarian function against environmental toxins, thereby preserving fertility and ensuring the health of future generations. The detailed exploration of these mechanisms adds depth to our understanding of ovarian physiology and pathophysiology, providing a solid foundation for future research and therapeutic interventions.
Gallic acid (GA), a plant-derived polyphenol, has been extensively studied for its robust antioxidant capabilities, which may confer a protective shield against cellular oxidative damage. 7 In the context of ovarian health, oxidative stress is a pivotal factor in the pathophysiology of BPA-induced ovarian toxicity. 4,8 The antioxidant action of GA is hypothesized to mitigate the oxidative damage in ovarian tissue by scavenging free radicals and upregulating the body’s own antioxidant defenses, thus preserving the integrity of ovarian cells and supporting normal follicular development and steroid hormone synthesis. 9,10 In a study with parallels, GA exhibited a protective role against oxidative toxicity in rat ovaries induced by cadmium, hinting at a similar protective potential against damage caused by BPA. 11 The underlying mechanisms are thought to involve the modulation of oxidative stress pathways and the stabilization of cellular antioxidant responses. This preclinical evidence supports the exploration of GA as a therapeutic agent in human medicine, with the potential to safeguard ovarian function against the onslaught of environmental toxicants like BPA.
In this study, the protective effects of GA against BPA-induced ovarian toxicity were explored through a methodology that incorporates biochemical, histological, and immunohistochemical analyses.
MATERIALS AND METHODS
Chemicals and animals
The chemicals used in this research were GA and BPA, sourced from Sigma-Aldrich, located in St. Louis, MO, USA, with catalog numbers G7384 and 42088, respectively. These compounds were selected for their relevance in the investigation of endocrine-disrupting effects and potential protective interventions. A total of 32 female Sprague Dawley rats, aged 3 months and weighing 250 ± 10 g, were obtained from ATADEM. Before initiating the experiment, the rats underwent a one-day period of acclimation to familiarize them with the laboratory environment. They were provided with a habitat that maintained regulated temperature and lighting, along with unrestricted access to nourishment and hydration. The experimental procedures received ethical clearance from the Atatürk University Animal Experiments Local Ethics Committee, under the authorization number 2021/11. Furthermore, all animal-related methods in this research adhered to the ethical guidelines established by the respective institution.
Experimental design
The rats were distributed into four distinct groups, each consisting of eight individuals, in a random allocation to ensure variability and scientific rigor in the experimental design.
Control group: The rats were administered a daily dose of 0.5 mL distilled water through oral gavage.
GA group: Each rat received a daily oral gavage of 20 mg/kg 11 body weight of GA, with no deviation from their regular diet.
BPA group: Rats in this group received a daily intraperitoneal injection of BPA at a dose of 100 mg/kg 12 body weight, maintaining their normal feeding schedule.
GA+BPA group: This set of rats was given 20 mg/kg of GA via oral gavage, succeeded by an intraperitoneal injection of 100 mg/kg of BPA, administered 30 min subsequent to the GA dose, all while their standard diet remained unchanged.
The duration of the experiment was 3 weeks.
Sample collection and euthanasia
Following the completion of the experimental protocol, the rats underwent deep sedation with thiopental sodium (20 mg/kg) and 5% sevoflurane to ensure humane handling during the final procedures. Subsequent to blood collection, euthanasia was carried out with an increased dose of thiopental sodium (50 mg/kg) to minimize suffering. The ovarian tissues were then meticulously harvested; with the right ovary being immediately stored at −80°C for subsequent biochemical assays, and the left ovary fixed in a 10% formaldehyde solution, preparing it for detailed histopathological and immunohistochemical analysis.
Hematoxylin-eosin staining method
Subsequent to the rats’ necropsy, the ovarian tissues were promptly preserved in a 10% solution of buffered formalin. Following preservation, the samples underwent standard histological processing. This entailed a progressive dehydration series using alcohol of increasing concentrations, clarification in xylene, and finally, impregnation with paraffin wax. The paraffin-embedded tissues were then sectioned at a thickness of 5 μm.
These sections were carefully arranged on glass slides and underwent staining using the hematoxylin and eosin (H&E) method, a technique integral for histological analysis. The H&E stain selectively accentuates the cellular and extracellular matrix components, thereby enhancing the contrast within the tissue sections. This contrast is crucial for distinguishing between healthy and diseased tissue structures under microscopic examination. Under light microscopy, the stained sections were evaluated for histopathological changes. The findings were semi-quantitatively assessed and categorized as absent (0), mild, 1 moderate, 2 or severe 3 based on the extent of the observed alterations. This method allows for a detailed examination of the tissue architecture and cellular morphology, providing insights into the potential impacts of the experimental treatments on ovarian structure and function.
Immunohistochemical method
The immunohistochemical analysis was performed to detect specific antigens within the ovarian tissues of the rats following necropsy. The tissues were fixed in 10% neutral buffered formalin and processed through routine alcohol-xylene series before being embedded in paraffin blocks. Tissue sections, precisely 5-μm thick, were prepared on slides coated with poly-L-lysine to enhance adhesion. These sections were first deparaffinized in xylene, then methodically rehydrated using a descending alcohol series. Following rehydration, the tissue sections were thoroughly rinsed with phosphate-buffered saline (PBS) and then incubated with a 3% solution of hydrogen peroxide for 10 min to inhibit endogenous peroxidase activity.
For antigen retrieval, the tissues were exposed to a designated antigen retrieval solution and microwaved for two 5-min intervals at 500 watts, a step essential for unmasking the target antigens. Prior to antibody application, a protein block was applied to the sections to minimize nonspecific antibody binding. The sections were then bathed in PBS once more and incubated at +4°C overnight with primary antibodies. These antibodies were directed against key inflammatory markers: TNFα, at a dilution of 1/1000 (Santa Cruz, Catalog No. sc-52746); IL-1β, also at a dilution of 1/1000 (Santa Cruz, Catalog No. sc-52012); and COX2, with a dilution of 1/300 (Abcam, Catalog No. ab15191). To determine the oxidative DNA damage and apoptotic pathways, the study included antibodies against 8-hydroxydeoxyguanosine (8-OHdG), with a dilution of 1/100 (Santa Cruz, Cat No. sc-66036) and cleaved caspase-3 at a dilution of 1/200 (Elabscience, Cat. No. E-AB-30004).
Upon completion of the primary antibody incubation period, the sections were then processed with a secondary antibody, using the Large Volume Detection System: anti-Polyvalent, HRP (Thermo Fisher, Catalog No: TP-125-HL) in concordance with the recommended guidelines provided by the manufacturer. The antigen–ntibody complexes were rendered visible by applying the chromogen 3,3′-diaminobenzidine. For histological differentiation, Mayer’s hematoxylin was used in the counterstaining process. The prepared slides were finally sealed under a layer of Entellan and read for detailed examination with a light microscope.
Immunopositivity in the ovarian tissues was assessed and scored based on the staining intensity and distribution of the immunoreactive products. 13,14
Oxidant and antioxidant parameter analysis
Sample preparation
Following histopathological evaluation, ovarian tissues were stored at −80°C. For biochemical analysis, tissues were pulverized under liquid nitrogen using a TissueLyser (Qiagen, Hilden, Germany). Homogenates were prepared by adding 1 mL of PBS to 50 mg of tissue. The homogenates were then centrifuged, and the supernatants were collected for further analysis.
Biochemical assays
The levels of malondialdehyde (MDA) and the activity of superoxide dismutase (SOD) in the ovarian tissue homogenates were determined. An enzyme-linked immunosorbent assay (ELISA) microplate reader was used to measure the absorbance of both standards and samples, which allowed for the construction of standard curves. The concentrations of MDA and SOD in the samples were calculated using the equations derived from the standard curves.
The assay to measure SOD activity uses the principle of nitroblue tetrazolium reduction inhibition. This inhibition is attributed to the superoxide anions produced by the xanthine/xanthine oxidase system. The methodology for this assay was delineated in detail by Sun et al. in their comprehensive study. 15 The MDA assay measures the reaction of lipid peroxidation products with thiobarbituric acid, as detailed by Ohkawa et al. 16 MDA levels were measured and expressed as nanomoles per milligram of protein (nmol/mg protein), whereas SOD activities were denoted as units per milligram of protein (U/mg protein). This normalization to protein content allows for the comparison of oxidative stress markers across samples with varying tissue densities and sizes.
Hormone biochemical analysis
Hormone levels of follicle-stimulating hormone (FSH) (BT Lab, Cat. No. EA0015Ra), luteinizing hormone (LH) (BT Lab, Cat. No. EA0013Ra), estrogen (BT Lab, Cat. No. EA0012Ra), and progesterone (BT Lab, Cat. No. EA0063Ra) in ovarian tissue homogenates were determined using specific ELISA kits (μQuant, Bad Friedrichshall, Biotek). For the preparation of the homogenates, 1 mL of PBS was added to 50 mg of dry tissue before centrifugation. The subsequent supernatant was applied to the antibody precoated wells of the microtiter plates. Following the manufacturer’s protocols, the hormone concentrations were calculated from the absorbance readings against standard curves, with results normalized to the protein content of the tissues for accurate comparison.
Statistical analysis
For the statistical evaluation of the data, the SPSS software (version 25.0; IBM SPSS Inc, Chicago, IL, USA) was used. The distribution of the dataset was assessed for normality using the Kolmogorov–Smirnov test. Descriptive statistics were presented as mean ± standard deviation. To discern significant differences among the groups, a one-way ANOVA test was conducted, followed by post hoc analysis with the Tukey test. A P value threshold of < 0.05 was established to denote statistical significance within a 95% confidence interval.
RESULTS
Histopathological outcomes of GA intervention on BPA-induced follicular degeneration through hematoxylin-eosin staining analysis
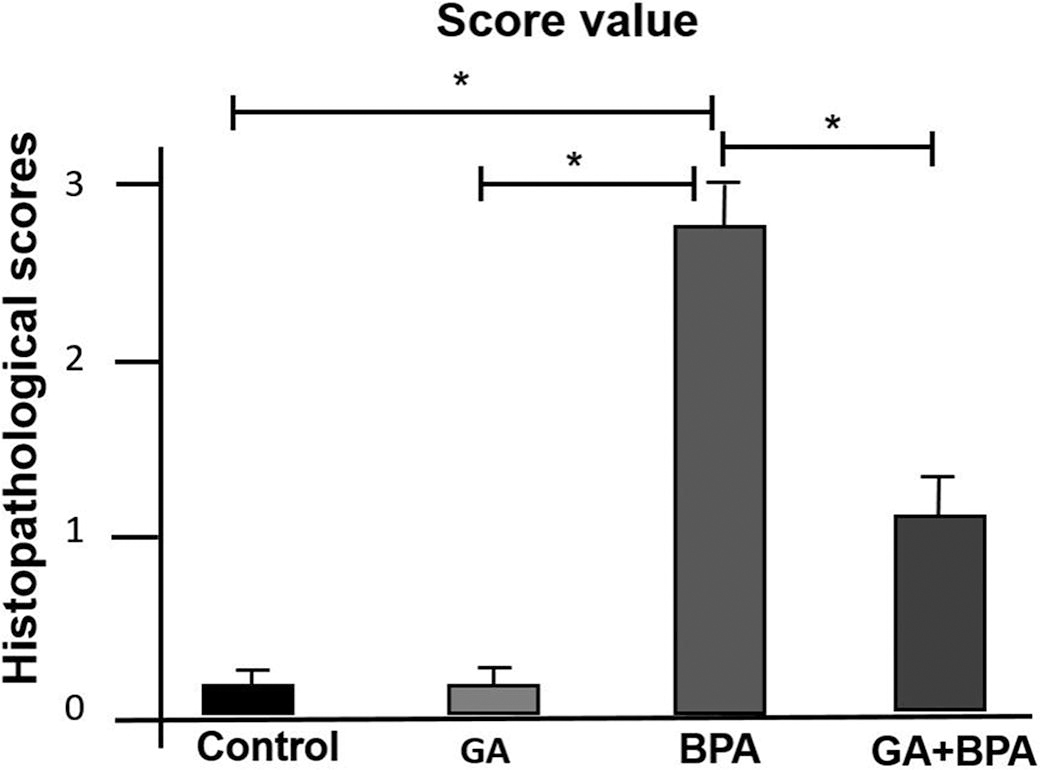
The histopathological evaluation, as illustrated in Figure 1 and Figure 2, substantiated significant differences among the experimental groups (P < 0.05). In the BPA-induced group, a substantial increase in the degeneration of secondary follicles was evident, underscoring the deleterious impact of BPA on ovarian morphology. Contrastingly, the GA group maintained follicular integrity akin to the control, indicating no inherent toxicity of GA on ovarian structures. Notably, the concomitant administration of GA with BPA attenuated the extent of follicular degeneration, suggesting a mitigative influence of GA on BPA-induced ovarian compromise. These variations, marked by alterations in granulosa cell configuration and follicular architecture, highlight the potential therapeutic efficacy of GA in preserving ovarian health against environmental toxicants. The statistical significance (P < 0.05) across different groups underscores the biological relevance of these findings.

Immunohistochemical analysis of TNF-α, COX-2, IL-1β, 8-OHdG, and caspase 3 in ovarian tissue
Immunohistochemical staining for inflammatory markers TNFα, COX2, IL-1β, 8-OHdG, and caspase 3 revealed statistically significant differences among the groups, as shown in Figure 3, Figure 4 and Table 1 (P < 0.05). The BPA group exhibited intense immunoreactivity, particularly in the ovarian stroma and granulosa cells of the follicles, indicating a strong inflammatory response. In contrast, the GA+BPA group displayed moderate levels of immunoreactivity, suggesting that GA may exert an anti-inflammatory effect, mitigating the BPA-induced inflammation.
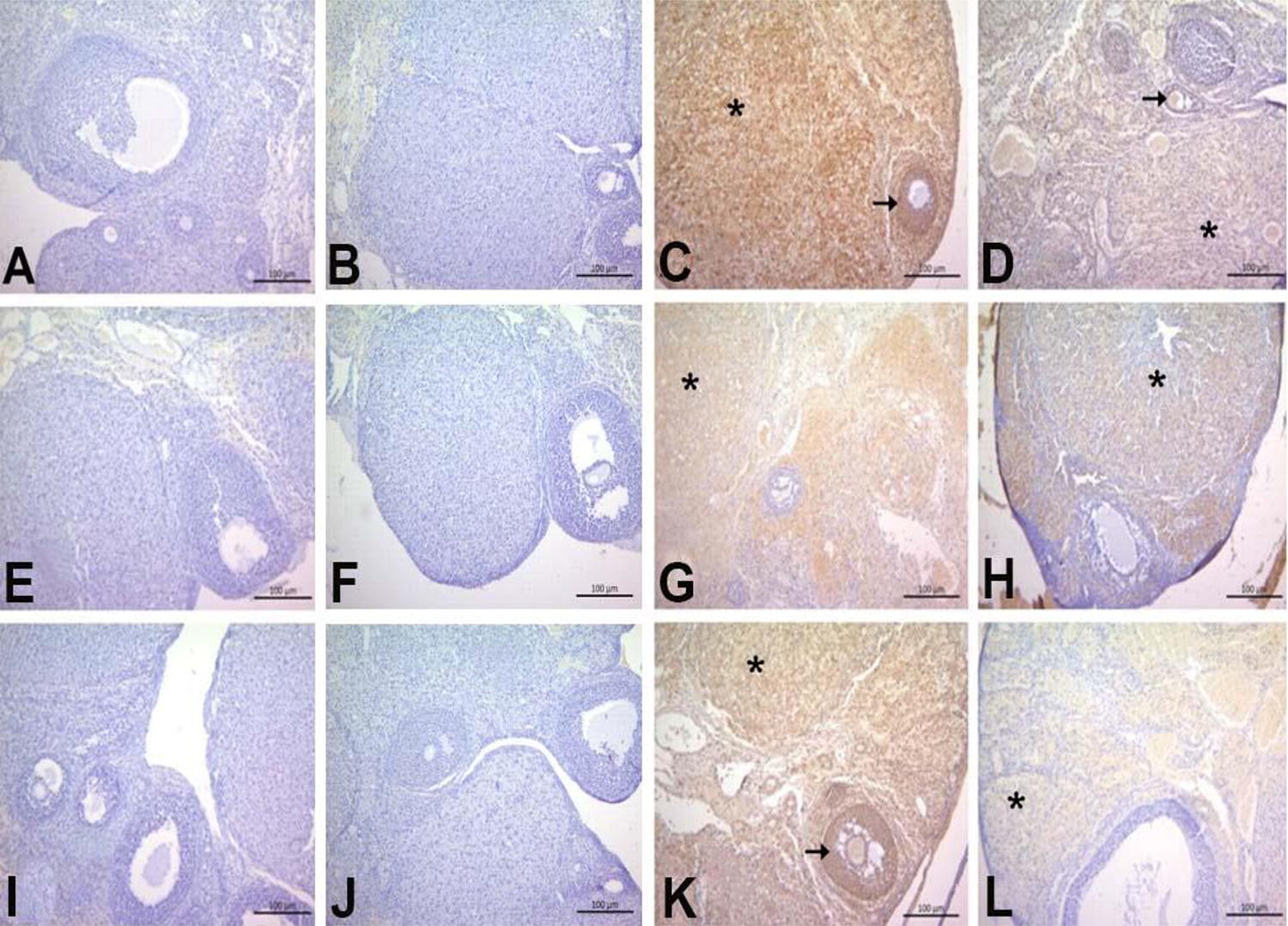
Immunohistochemical staining for TNFα, COX2, and IL-1β.
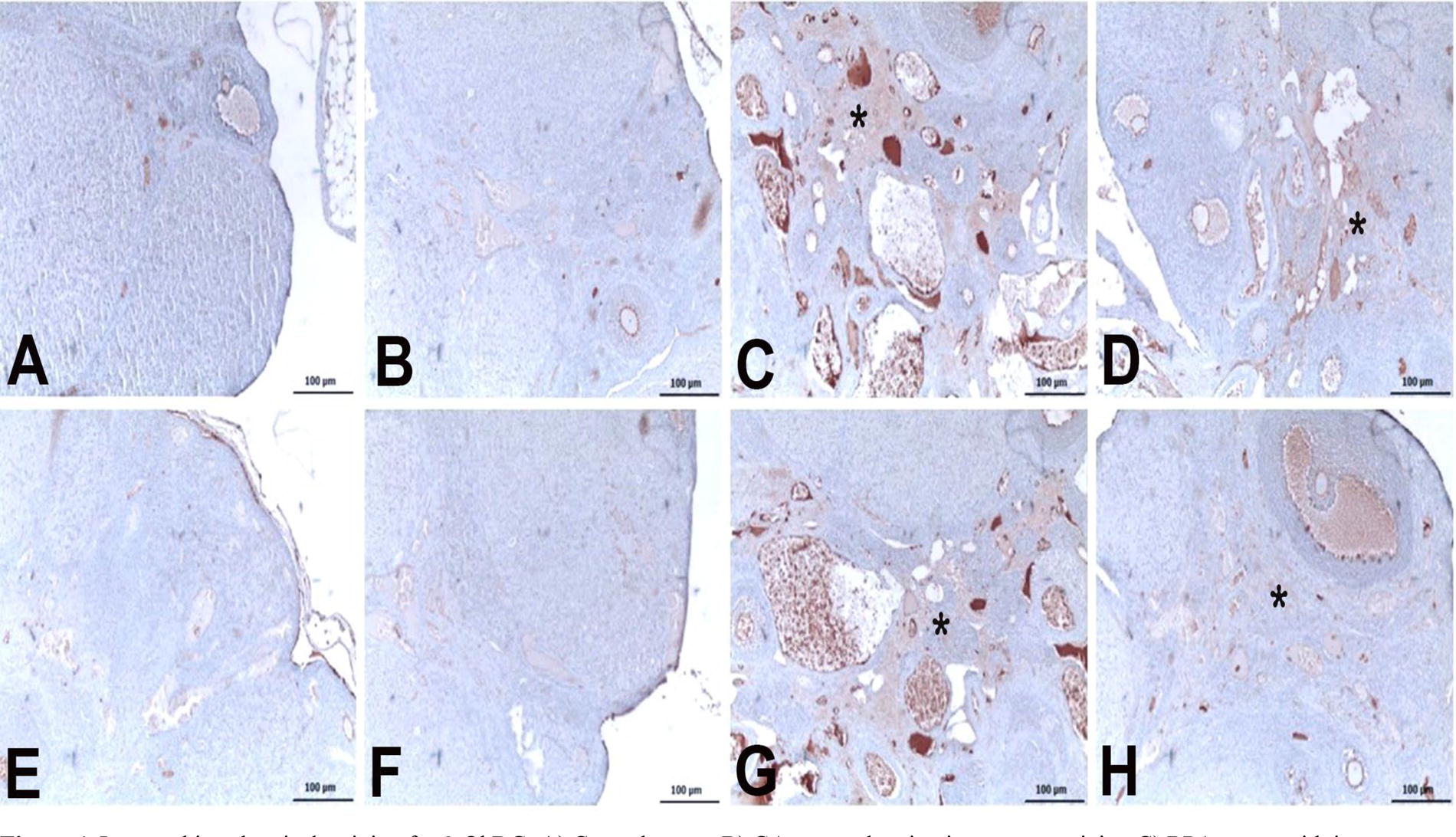
Immunohistochemical staining for 8-OHdG:
Immunohistochemical Cell Score Values for TNFα, COX2, IL-1β, 8-OHdG, and Caspase 3 Across Groups
Different superscript letters within the same column denote statistically significant differences between groups (P < 0.05).
BPA, bisphenol A; GA, gallic acid.
Table 1 reports the immunoreactive cell scores for TNFα, COX2, IL-1β, 8-OHdG, and caspase 3 across different groups. In the control and GA groups, the scores were low, indicating minimal immunoreactivity similar to baseline levels. For the BPA group, significantly higher scores were observed for all markers, demonstrating a marked inflammatory response and increased apoptosis as indicated by elevated levels of TNFα, COX2, IL-1β, as well as 8-OHdG and caspase 3, which are markers of oxidative stress and apoptosis, respectively. The GA+BPA treatment group showed reduced scores across all markers compared with the BPA-only group, with significant decreases in the levels of 8-OHdG (P < 0.05) and caspase 3 (P < 0.05).
The staining intensity for TNFα was markedly severe in the BPA group, whereas it was moderate in the GA+BPA group. The immunopositivity was localized to the stroma and granulosa cells of the follicles. COX2 immunopositivity was severe in the BPA group but moderate in the GA+BPA group, with localization exclusively in the stroma. Similar patterns were observed for IL-1β, with the BPA group showing intense immunopositivity in the stroma and granulosa cells, which was reduced to moderate levels in the GA+BPA group (Fig. 2).
Figure 4 illustrates the immunohistochemical staining for 8-OHdG and caspase 3 across experimental groups. Figure 4A and Figure 4E display minimal to no staining in the control group, suggesting negligible oxidative stress and apoptotic activity. Similarly, Figure 4B and Figure 4F indicate an absence of immunoreactivity in the GA group, aligning with the control results. In contrast, Figure 4C and Figure 4G show intense staining in the BPA group within the stroma, denoted by asterisks, indicative of a high level of oxidative DNA damage and apoptosis. Figure 4D, 4H, which represent the GA+BPA treatment group, show a moderate level of staining for 8-OHdG and a mild level for caspase 3, respectively, suggesting a mitigated response to oxidative stress and apoptosis compared with the BPA group. These findings are consistent with the quantitative data presented in Table 2, confirming the visual assessment of the protective effect of GA against BPA-induced cellular damage.
Biochemical Hormone Parameters in Ovarian Tissues Across the Groups
Results are expressed as mean ± standard deviation. P value from one-way ANOVA test. Superscript letters (a,b,c,d) indicate post hoc Tukey test results following one-way ANOVA. a,b,c,dDifferent superscript letters within the same column denote statistically significant differences between groups, the same superscript letters within the same column denote not statistically significant differences between groups (P < 0.05).
ANOVA, analysis of variance; FSH, follicle-stimulating hormone; LH, luteinizing hormone.
Oxidative stress marker variations among treatment groups
The assessment of oxidative stress markers, specifically MDA levels and SOD activities, revealed significant variances among the experimental groups (Fig. 5, P < 0.001 for both MDA and SOD).
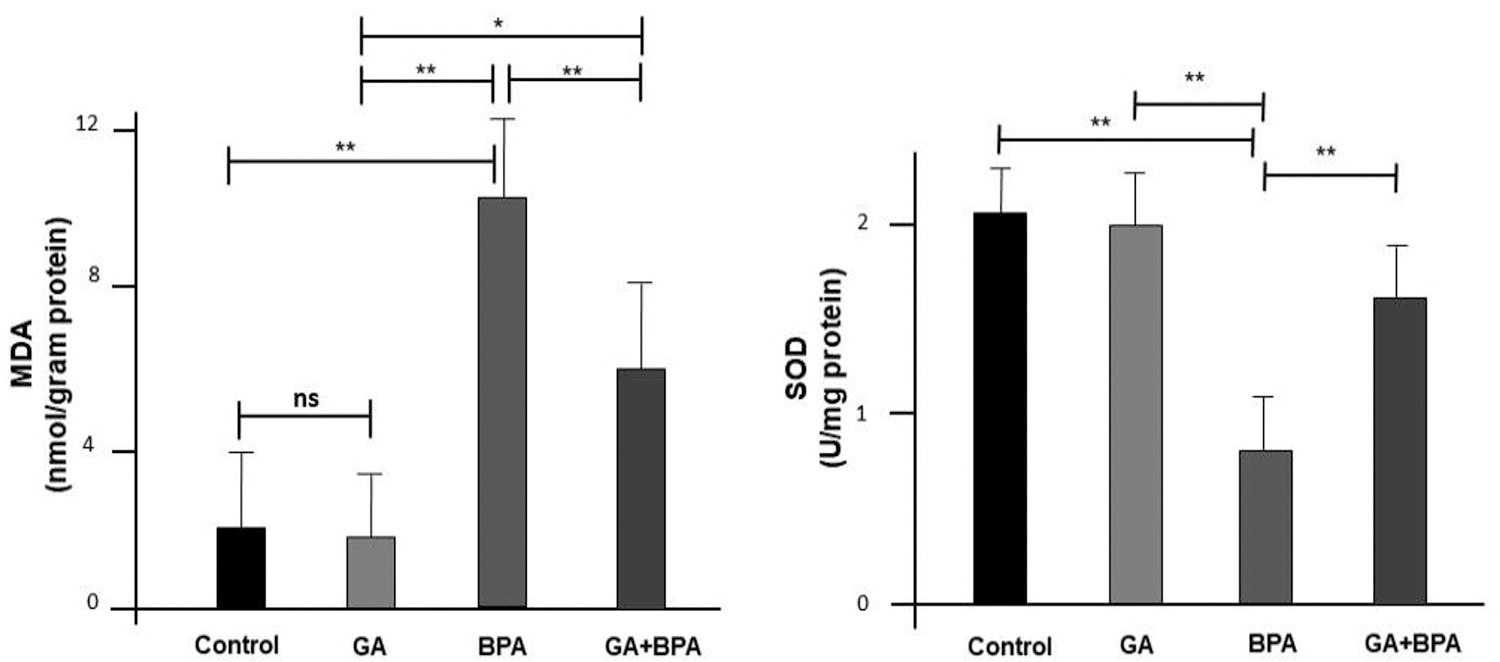
Oxidative stress markers in ovarian tissues across the groups (n = 8), GA: Gallik Asit (20 mg/) group, BA: bisphenol A (100 mg/kg) group. Across the biomarkers, the “ns” notation indicates no significant difference between the groups being compared at those points, while the ‘*’ denote significant differences where the treatment effects are evident. *P < 0.05, **P < 0.01. MDA, malondialdehyde; SOD, superoxide dismutase.
The MDA levels, indicative of lipid peroxidation, were significantly different across all pairwise comparisons except between the control and GA groups. This suggests that GA does not contribute to lipid peroxidation in a manner that is statistically distinguishable from the control. In contrast, the BPA group exhibited a dramatic increase in MDA levels, which was significantly mitigated in the GA+BPA group, though still elevated compared with the control and GA groups.
Similarly, SOD activity, a measure of antioxidant defense, differed significantly between groups, with the exception of the control and GA groups, and the control and GA+BPA groups. The BPA group showed a marked decrease in SOD activity, indicating a reduction in antioxidant defense mechanisms. The GA+BPA group, while showing improved SOD activity compared with the BPA group, did not reach the levels observed in the control group, suggesting partial recovery of antioxidant capacity with GA treatment.
Hormonal profile alterations in ovarian tissues
The assays measuring hormone levels, specifically FSH, LH, estrogen, and progesterone in ovarian tissues, demonstrated notable variances between the study groups (Table 2) (P < 0.05 for all comparisons).
FSH and estrogen levels were significantly different when comparing the control to the BPA group and the GA group to the BPA group. This indicates that BPA exposure significantly impacts the synthesis or regulation of these hormones. The LH and progesterone levels showed significant differences between the control and BPA groups, the GA and BPA groups, and the GA and GA+BPA groups. Notably, progesterone levels were significantly different between the BPA and GA+BPA groups, suggesting that GA may influence the regulation of progesterone in the presence of BPA (P < 0.05 for all comparisons).
DISCUSSION
This study investigated GA’s protective effects against BPA-induced ovarian toxicity, focusing on histopathological changes, immunohistochemical markers, oxidative stress parameters, and hormone levels.
BPA-induced inflammation can damage ovarian tissue, impacting essential processes such as folliculogenesis and steroidogenesis, and lead to disorders including ovarian cysts, endometriosis, and reduced ovarian reserve. 9,17 Our study’s immunohistochemical results highlight the inflammatory effects of BPA on ovarian tissue and GA’s potential to counteract these effects. The increased levels of TNF-α, COX-2, and IL-1β observed in BPA-exposed tissues signal an inflammatory response, with these markers critical to inflammation development. TNF-α is a key cytokine in acute inflammation, COX-2 produces proinflammatory prostaglandins, and IL-1β is crucial in the cellular response to harm and infection. 18,19 Administering GA with BPA reduces inflammation, as seen by lower TNF-α, COX-2, and IL-1β levels, indicating GA’s anti-inflammatory properties. GA may suppress the TNFα and pathway, crucial for inflammatory gene control, and enhance antioxidant defenses, indirectly reducing inflammation by lowering oxidative stress.
The investigation into the roles of 8-OHdG and caspase 3 in BPA-induced ovarian toxicity and the protective effect of GA provides critical insights into the molecular mechanisms of cellular damage and defense. 8-OHdG, a marker of oxidative DNA damage, 20,21 and caspase 3, a key executor in the apoptotic pathway, serve as pivotal indicators of the physiological stress and cellular death mechanisms activated in response to BPA exposure. 22 Oxidative stress, a consequence of BPA exposure, leads to increased 8-OHdG levels, reflecting the damage incurred by DNA through ROS interaction. 4 This damage is a crucial step toward cell malfunction and apoptosis, marked by the activation of caspase 3. Activation of caspase 3 is a critical phase in apoptosis, denoting an irreversible commitment to cell death. 23 Altındağ et al. have demonstrated that GA mitigates cisplatin-induced testicular toxicity by preserving spermatogonia, leydig, and sertoli cells, alongside improving testicular volume and serum testosterone levels. 24 Similarly, Demir et al. reported that GA protects against cisplatin-induced ovarian toxicity by reducing oxidative stress markers and enhancing antioxidant status, alongside restoring structural and functional integrity of the ovaries. 21 In another study, Gelen et al. found that GA ameliorates cadmium-induced hepatotoxicity through a decrease in liver enzyme levels, oxidative stress markers, and inflammation, while enhancing the antioxidant defense system. 25 Chuang et al. further identified that GA induces apoptosis in lung fibroblasts through ROS-dependent ATM/p53 signaling, showcasing its potential in fibrotic and tumor cell lines. 26 Our study found that GA notably decreased 8-OHdG and caspase 3 levels in BPA-exposed ovarian tissue, signaling reduced oxidative DNA damage and apoptosis. This supports previous findings of GA’s protective roles, affirming its antioxidative, anti-inflammatory, and anti-apoptotic effects in various toxicity models.
BPA disrupts oxidative balance by upregulating oxidative mediators, reducing antioxidant defenses, causing mitochondrial dysfunction, altering cell signaling, and inducing apoptosis. 27 –29 Research has shown that BPA exposure reduces antioxidants like SOD, catalase, GR, and GSH-Px and increases hydrogen peroxide and lipid peroxidation, harming liver, sperm quality, and organ development in animals. 30 –33 ROS generation from BPA exposure varies by cell type, BPA concentration, exposure duration, and detection methods used. 8,34,35 Moreover, human studies show a link between BPA and inflammation, with higher urinary BPA levels correlating with oxidative stress and systemic inflammation markers, affecting white blood cell counts. 36 This gender and hormonal status-dependent effect of BPA may be attributed to the differential expression of estrogen receptors (ERs), which vary with gender and age. 37 In postmenopausal women, the lower hormone levels could facilitate a greater binding affinity of BPA to ERs, leading to adverse cellular responses linked to oxidative stress and inflammation. 4,37 Our study confirms GA’s protective role against BPA’s oxidative damage, as shown by reduced MDA levels and increased SOD activity in the GA+BPA group, suggesting GA enhances antioxidant defenses and mitochondrial integrity, countering BPA-induced oxidative stress. GA’s antioxidant properties, which neutralize free radicals, chelate metals, modulate enzyme activities, and mitigate BPA’s effects on ovarian tissue. 38
BPA exposure alters FSH, LH, estrogen, and progesterone levels, reflecting its endocrine-disrupting effects, particularly on ovarian function. BPA can mimic or block hormones, interfere with receptor signaling, and change hormone production and breakdown. 39,40 GA treatment moderates BPA-induced hormonal disruptions, notably normalizing progesterone, highlighting polyphenols' protective role against toxins. GA’s mitigation of endocrine dysfunction is linked to its antioxidant properties, combating BPA’s oxidative stress and supporting steroidogenesis. 41,42 GA’s improvement of progesterone levels in the GA+BPA group indicates it may protect the corpus luteum, often affected by BPA, essential for reproduction and embryogenesis. 30 GA may enhance progesterone by influencing steroidogenesis enzymes like cytochrome P450scc, which converts cholesterol to pregnenolone and acts as a selective estrogen receptor modulator, blocking BPA’s estrogen receptor binding to minimize hormonal imbalances. 43,44 The findings of this study are in line with those of, where GA was shown to protect against BPA-induced alterations in hormone levels in a rodent model. 45 Similarly, numerous studies have demonstrated that antioxidants can prevent BPA-induced changes in reproductive hormone levels, further supporting the hypothesis that oxidative stress plays a role in the endocrine-disrupting mechanisms of BPA. 8,46 –48
CONCLUSION
The study conclusively demonstrates that BPA induces significant ovarian toxicity, as evidenced by the degeneration of secondary follicles and altered levels of critical reproductive hormones (Graphical Abstract). GA, when co-administered with BPA, appears to exert a protective effect, ameliorating both the histopathological damage to ovarian tissues and the disruption of hormonal balance. The immunohistochemical and oxidative stress markers further substantiate GA’s role in mitigating the inflammatory response and oxidative stress induced by BPA exposure. These findings suggest that GA has potential therapeutic value in shielding ovarian tissues from BPA-induced damage, highlighting its antioxidant and anti-inflammatory properties. The study underscores the need for further research into the use of GA as a protective agent against environmental toxins that disrupt endocrine and reproductive health.
Footnotes
DATA AVAILABILITY
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
AUTHOR DISCLOSURE STATEMENT
The authors have no conflict of interest.
FUNDING INFORMATION
No funding was received for this article.