
Abstract
Osteoporotic fractures seriously affect the quality of life of the elderly. Panax notoginseng saponins (PNS) have the potential function of preventing osteoporosis. The Phosphatidylinositol 3-kinase (PI3K)/protein kinase (AKT)/mammalian target of rapamycin (mTOR) pathway is involved in the regulation of osteoporosis and has been proven to be related to VEGF secretion and angiogenesis. Therefore, this study aimed to explore the effects of PNS on ovariectomized rats with osteoporotic fracture through the PI3K/AKT/mTOR pathway and angiogenesis-related factors. Female Sprague–Dawley rats were randomly divided into normal control, fracture model, ovariectomized fracture model, low-dose PNS (100 mg/kg/d), and high-dose PNS (200 mg/kg/d). The ovariectomized rat fracture model was established. In low and high dose groups, PNS was administered intraperitoneally. The vascularization of fracture ends was detected in vitro by micro-CT on the 7th, 14th, and 21st day after modeling, and the area and number of blood vessels in the unit field of vision of the callus healing plane were seen by hematoxylin-eosin staining. The expression levels of PI3K, AKT1, mTOR, hypoxia inducible factor-1; VEGF: vascular endothelial growth factor (HIF-1), VEGF, Ang-1, VEGFR2, and angiopoietin like 2 Gene (ANGPTL2) were determined using Western blotting. In the PNS treatment group, the area of cortical bone increased, the area of callus decreased, and the number and area of blood vessels increased significantly when compared with the ovariectomized fracture model group. PNS regulates the PI3K/AKT/mTOR signaling pathway and promotes the expression of vascular-related cytokines (VEGF, Ang-1, VEGFR2, and ANGPTL2) in osteoporotic fractures. PNS may regulate the expression of vascular-related factors through the PI3K/AKT/mTOR pathway and promote the healing of osteoporotic fractures in ovariectomized rats.
INTRODUCTION
The incidence of age-related diseases rises progressively as the population ages. Osteoporosis is a common orthopedic illness whose incidence has lately increased. 1 Among the many problems of osteoporosis, osteoporotic fracture has the most significant impact on the elderly’s quality of life. The etiology of osteoporosis is that osteoblasts’ osteogenic capacity is impaired while osteoclasts are active, resulting in decreased bone mass, decreased bone strength, and increased brittleness. The fracture is produced by slight trauma or nontraumatic elements in everyday living. Hence, it is sometimes called low-energy fracture or brittle fracture. 2 Osteoporotic fracture healing is distinguished by poor bone quality, prolonged fracture healing time, and a high rate of refracture. 3
Panax notoginseng saponins (PNS), flavonoids, volatile oil, amino acids, carbohydrates, and a variety of trace elements. Panax notoginseng’s principal active ingredient is its total saponins, which include a range of monomer saponins. PNS can dilate blood vessels, drop arterial blood pressure, reduce myocardial oxygen consumption, suppress platelet aggregation, lengthen coagulation time, lower blood cholesterol levels, eliminate free radicals, act as an anti-inflammatory and antioxidant, and increase coronary blood flow. 4 It is primarily used to treat cerebral infarction, transient ischemic attack, cerebrovascular disease sequelae, cerebral arteriosclerosis, 5,6 and ischemic heart disease. 4 A previous study revealed that PNS could help prevent osteoporosis. 7
Phosphatidylinositol 3-kinase (PI3K)/AKT/mammalian target of rapamycin (mTOR) is a crucial intracellular signal pathway that regulates the cell cycle and is directly linked to cell dormancy, proliferation, carcinogenesis, and longevity. 8 The activation of PI3K leads to the production of second messenger hosphatidyl-inositol-3,4,5-trisphosphate (PIP3) on the plasma membrane. PIP3 binds to AKT and Recombinant Pyruvate Dehydrogenase Kinase Isozyme 1 (PDK)1, which contain the PH domain in the cell, and enhances PDK1 to phosphorylate Thr308 on AKT protein, resulting in AKT activation. Activated AKT modulates cell function through phosphorylation of various downstream factors, including enzymes, kinases, and transcription factors. 9 mTOR is an important serine threonine protein kinase downstream of PI3K/AKT, which can regulate cell proliferation and survival through ribosome kinase activation. 10 Previous studies indicated that the PI3K/AKT cell signaling pathway participates in osteoporosis regulation, 11,12 and mTOR is a therapeutic target in osteoporosis. 13 The PI3K/AKT/mTOR pathway has been shown to be related to VEGF secretion and angiogenesis, 14 and there are studies supporting the influence of PNS on the PI3K/AKT/mToR signaling pathway. 15,16 However, the mechanism of panax notoginseng saponins on the PI3K/AKT/mTOR pathway and angiogenesis in osteoporotic fractures has not been reported.
Therefore, in this study, the osteoporotic fracture model of ovariectomized rats was used to explore the effect of PNS on fracture healing in rats and its regulatory mechanism on the PI3K/AKT/mTOR signaling pathway and angiogenesis-related cytokines. It is expected to provide theoretical and experimental basis for the clinical use of PNS in the treatment of osteoporotic fractures.
MATERIALS AND METHODS
Animals
Three months old female Sprague–Dawley (SD) rats were obtained from Changsha Tianqin Biotechnology Co., Ltd. [license number: SCXK (Huang) 2019-0014] and Shanghai Xipu Bikai Experimental Animal Co., Ltd. [license number: SCXK (Shanghai) 2018-0006]. The rats were given normal feed and water ad libitum and raised in cages under controlled temperature, 20–26°C, and humidity, 40%–70%, conditions.
This study was approved by the Animal Care and Use Committee of the authors’ institution and conducted according to the Principles of Laboratory Animal Care (National Society for Medical Research).
Main materials
PNS (F90699a1, Harbin Zhenbao Pharmaceutical Co., Ltd., Heilongjiang, P.R. China); Kirschner wire (A20316, Yancheng Aisikang Medical Equipment Co., Ltd., Jiangsu, P.R. China); internal control primary antibody: mouse monoclonal anti-GAPDH (1/2000, TA-08) and secondary antibody: goat anti-mouse IgG (H + L) horseradish peroxidase conjugate (1/2000, ZB-2305) (ZSGB-Bio, Beijing, P.R. China); target primary antibody: rabbit anti-PI3K (1/1000, ab191606) and mouse anti-AKT1 (1/500, ab8805) (Abcam PLC, Cambridge, UK), rabbit anti-mTOR (1/1000, 2972S, CST, Danvers, MA, USA), rabbit anti-HIF-1α (1/500, PB0245) and rabbit anti-VEGF (1/500, BA0407) (Boster Biological Technology, Pleasanton, CA, USA), and secondary antibody: goat anti-rabbit IgG (H + L) horseradish peroxidase (1/2000, ZB-2301, ZSGB-Bio, Beijing, P.R. China); Super ECL Plus detection reagent (RJ239676, Thermo Fisher Scientific, Waltham, MA, USA).
Experimental grouping
The rats were randomly assigned into normal control, fracture model control, ovariectomized fracture model control, ovariectomized fracture model + low-dose PNS (100 mg/kg/d) (low-dose PNS), and ovariectomized fracture model + high-dose PNS (200 mg/kg/d) (high-dose PNS) groups.
A total of 90 rats were used. Each group had 18 rats. The sample size was determined based on the preliminary study. The number of rats required in the present study was kept to the minimum. A computer-generated random number table was used to perform the randomization.
Rat model and PNS administration
The rats were anesthetized and fixed laterally. An incision of 1–2 cm was made in the lateral abdomen. The skin, fascia, and muscle were cut in turn or separated bluntly with scissors, and the white and moist fat mass exposed. By separating the fold of fat mass, the pink and bright cauliflower-like ovary can be seen. The junction of the uterus and fallopian tube were ligated, and the ovary and part of fallopian tube were removed. The adipose tissue was placed into the abdominal cavity after ligation, and penicillin was used to prevent infection and the incision was sutured. One month later, the fracture model was established. 17 After administration of anesthesia, the fracture was fixed on the side and disinfected with iodophor. The skin was cut open at 1.5–2 cm at the junction of the middle and upper 1/3 of the anterolateral tibial shaft to expose the tibial shaft. The scalpel was transversely cut off, resulting in fracture. The tibial shaft was fixed by retrograde upward driving with a 0.8 mm Kirschner needle (approximately 17 mm in length) and then disinfected and sewed. After fracture modeling, each rat in the low-dose and high-dose PNS group was given intraperitoneal injections of 100 mg/kg/day and 200 mg/kg/day PNS, respectively. PNS was diluted to 25 mg/mL and 50 mg/mL for low-dose and high-dose solution, respectively. After ovariectomy and fracture modeling, 6 rats per group were euthanized at each time point (7th, 14th, and 21st day). Three rats per group were randomly allocated for micro-CT detection, and 3 rats per group were randomly allocated for hematoxylin-eosin (HE) staining and Western blot for each time point. The tibia samples were obtained under aseptic conditions.
The investigators performing the subsequent steps of experiments and data analysis were blinded to the group allocation.
Micro-CT detection
The tibia samples were fixed in 4% paraformaldehyde solution and kept in a refrigerator at 4°C for computed tomography (CT) detection. The Scanner software of SkyScan1174 Micro-CT was used to scan the samples at 800 μA 50 kV, scan resolution 12 μm, and field size 1304 × 1024. The fracture area with thickness of 6 mm was set as three-dimensional (3D) reconstruction of interest area (ROI), the N-Recon software was used to reconstruct the 3D image, the CT-AN software was used for 3D analysis, and the tissue mineral density (TMD) of the cortical bone in the ROI was measured.
HE staining
Tissues were washed with running water, and 5% nitrate was used for decalcification for 3 days. The samples were dehydrated with the solutions of 70%, 80%, and 90% ethanol and mixture of pure alcohol and xylene for 15 min. They were treated with xylene I and II for 15 min each (until clear), paraffin embedded, and cut into slices. The slices were baked, dewaxed, and hydrated. They were subjected to hematoxylin staining for 3 min, hydrochloric acid ethanol differentiation for 15 s, and bluing for 15 s. After washing, they were stained with eosin for 3 min. Then, they were washed with running water, dehydrated, cleared, mounted, and examined under a microscope. The numbers of blood vessels and vascular areas were calculated using “ImageJ” software. This was performed by randomly selecting a sufficient number of fields of view, calculating the number of microvessels in a unit area and the area of blood vessels in different fields of view. Image binarization was performed to distinguish the image contour and establish an irregular selection area to measure or mark the contour of the selection area. For detailed methods, please refer to the following link: https://www.163.com/dy/article/G7970I9E0516W2BQ.html
Western blot
The tissues were ground into powder in liquid nitrogen. Lysis buffer was added, and the sample was ground again. Then, the tissue homogenate was placed into an Eppendorf (EP) tube and lysed on ice for 30 min. Then, they were centrifuged at 4°C (10,000 ×g) for 15 min. The supernatant was obtained, and the proteins were extracted. The proteins (50 μg/lane) were loaded for sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred to the PVDF membrane. The membrane was washed, and blocking was performed with a buffer. The primary antibodies were added, and incubation was performed at 4°C overnight. The membrane was rinsed, the secondary antibody was added, and incubation was performed at room temperature for 1 h. Immunoblotting was conducted using an enhanced chemiluminescence kit (PerkinElmer Inc.). Quantification was performed with Imagequant LAS4000 (GE Healthcare, Japan). The vascular-related cytokines including VEGF, Ang-1, VEGFR2, and ANGPTL2 were determined.
Statistical analysis
The statistical analysis was conducted using GraphPad Prism 7 software. The differences among groups were compared using the Bonferroni test, with P < 0.05 considered statistically significant.
RESULTS
Effect of PNS on bone area of fractured rats by microscopic CT scanning
The micro-CT scanning results are shown in Figure 1. It was expected to allocate 3 rats per group for micro-CT detection for each time point, but a problem that occurred during the sampling process resulted in a few samples that could not be detected, so only 2 rats per group were used in micro-CT detection. Compared with the control group, the cortical bone area of Bone Volume (BV) in the ovariectomized fracture model was decreased, whereas the callus area of BV was increased. Compared with the ovariectomized fracture model, the cortical bone area of BV decreased first and then increased, and the callus area of BV was remarkedly decreased in the low- and the high-dose PNS groups on the 7th, 14th, and 21st day. However, there was no obvious change in the cortical TMD in each group.
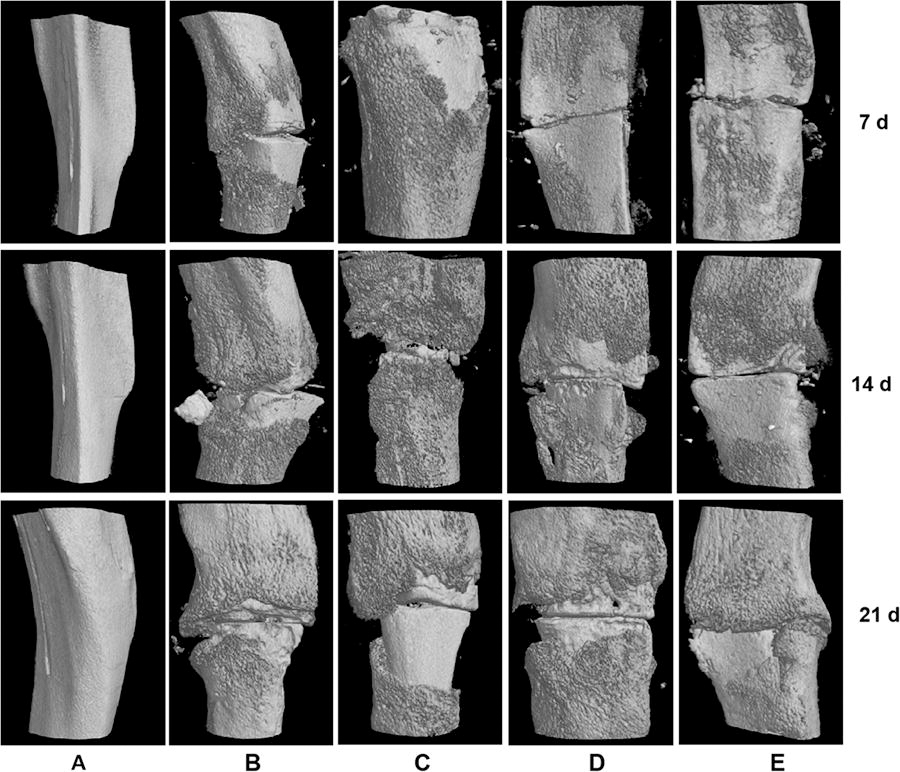
Micro-CT detection: the cortical bone area and the callus area of BV and cortical tissue mineral density of the rat samples in each group on the 7th, 14th, and 21st day. n = 2 per group for each time point. The data were presented as means ± SD (
Effect of PNS on the number of blood vessels after fracture modeling
The results of HE staining are shown in Figure 2. Compared with the fracture model and ovariectomized fracture model groups, the number of blood vessels in the low- and the high-dose PNS groups increased significantly, and it was more significant in the high-dose PNS group on the 7th day. Compared with the normal control group, the vascular area in the fracture model control and the ovariectomized fracture model control groups showed no significant differences, but it was significantly increased in the low- and the high-dose PNS groups compared with the ovariectomized fracture model group.
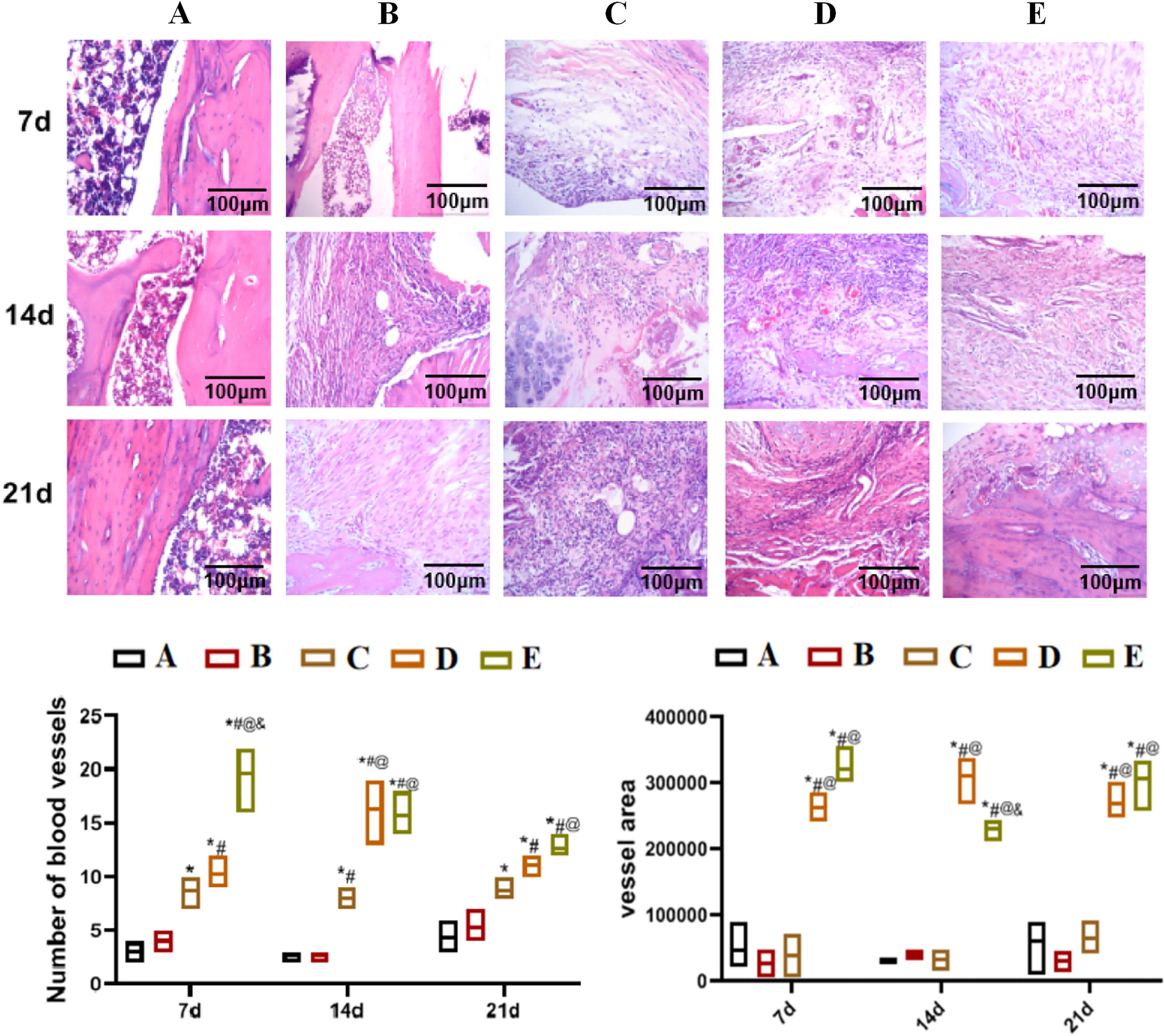
Hematoxylin-eosin staining detection of pathological changes in the knee joint tissues of rats in each group on the 7th, 14th, and 21st day. The significance of differences between the two groups was analyzed by one-way and two-way ANOVA, with P < 0.05 indicating a significant difference.
Effect of PNS on the expression of related proteins in fracture rats
The protein expressions on the 7th day after modeling are presented in Figure 3. Compared with the normal control group, the AKT1, HIF-1α, mTOR, VEGF, and Vascular Endothelial Growth Factor Receptor; Ang-1: Angiopoietin (VEGFR-2) protein expression levels in the ovariectomized fracture model group were significantly increased. Compared with the ovariectomized fracture model group, the AKT1, Ang-1, ANGPTL2, and VEGFR-2 protein expression levels in the low-dose PNS group were significantly increased; all protein expression levels in the high-dose PNS group were significantly increased.

Western blot detection of the protein expression in callus tissues of rats in each group on the 7th day after modeling.
The protein expressions on the 14th day after modeling are shown in Figure 4. Compared with the normal control group, the AKT1, ANGPTL2, HIF-1α, PI3K, VEGF, and VEGFR-2 protein expression levels in the ovariectomized fracture model group were significantly increased. Compared with the ovariectomized fracture model group, all protein expression levels in the low-dose PNS and the AKT1, Ang-1, ANGPTL2, HIF-1α, mTOR, PI3K, and VEGF protein expression levels in the high-dose PNS groups were significantly upregulated.
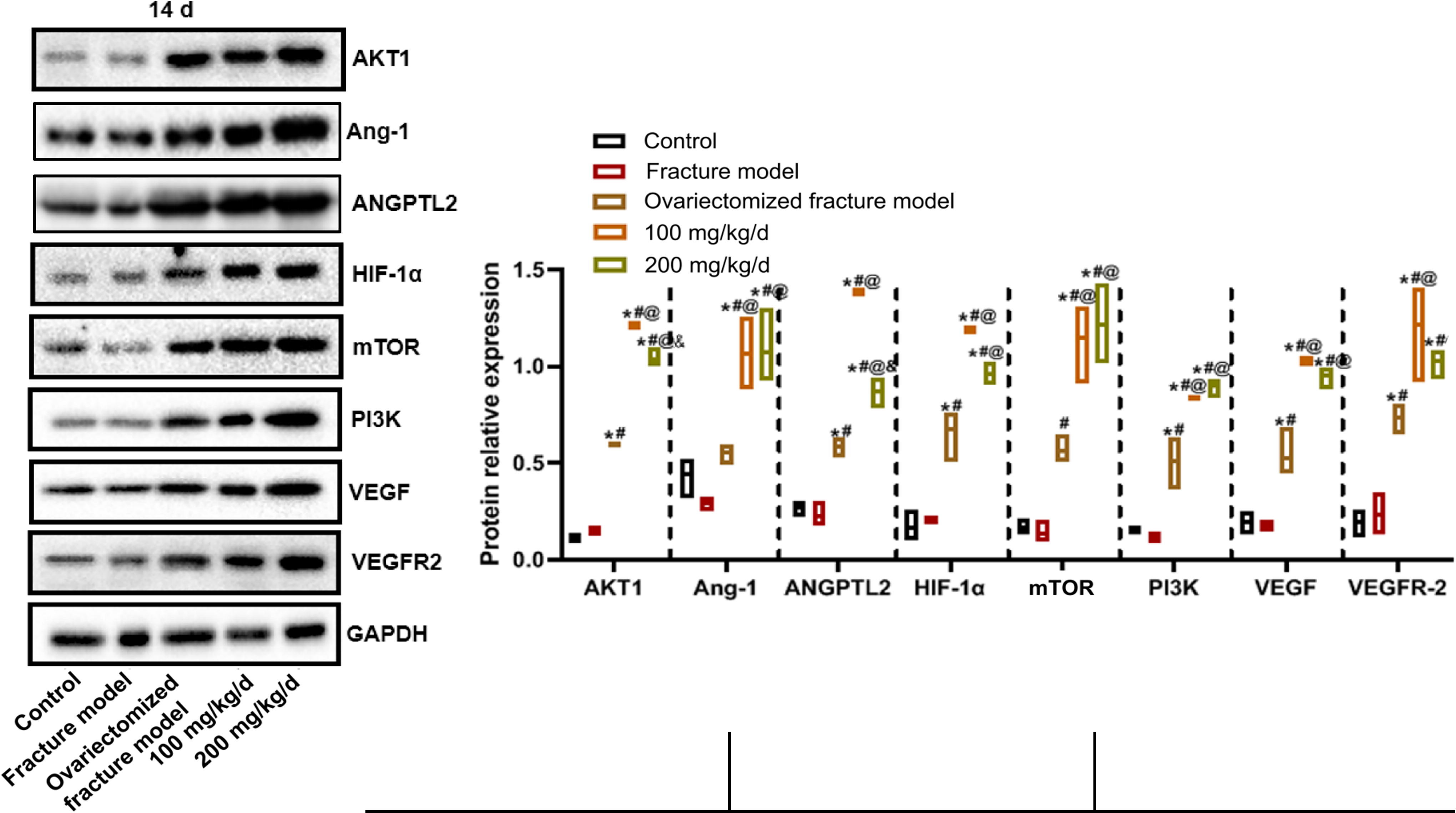
Western blot detection of the protein expression in callus tissue of rats in each group on the 14th day after modeling.
The protein expressions on the 21st day after modeling are illustrated in Figure 5. Compared with the normal control group, the AKT1, ANGPTL2, HIF-1α, mTOR, PI3K, VEGF, and VEGFR-2 protein expression levels in the ovariectomized fracture model group were significantly increased. Compared with the ovariectomized fracture model group, the AKT1, Ang1, HIF-1α, mTOR, and PI3K protein expression levels in the low-dose PNS group were significantly upregulated, while the AKT1, Ang-1, HIF-1α, and PI3K expression levels in the high-dose PNS group were significantly upregulated.
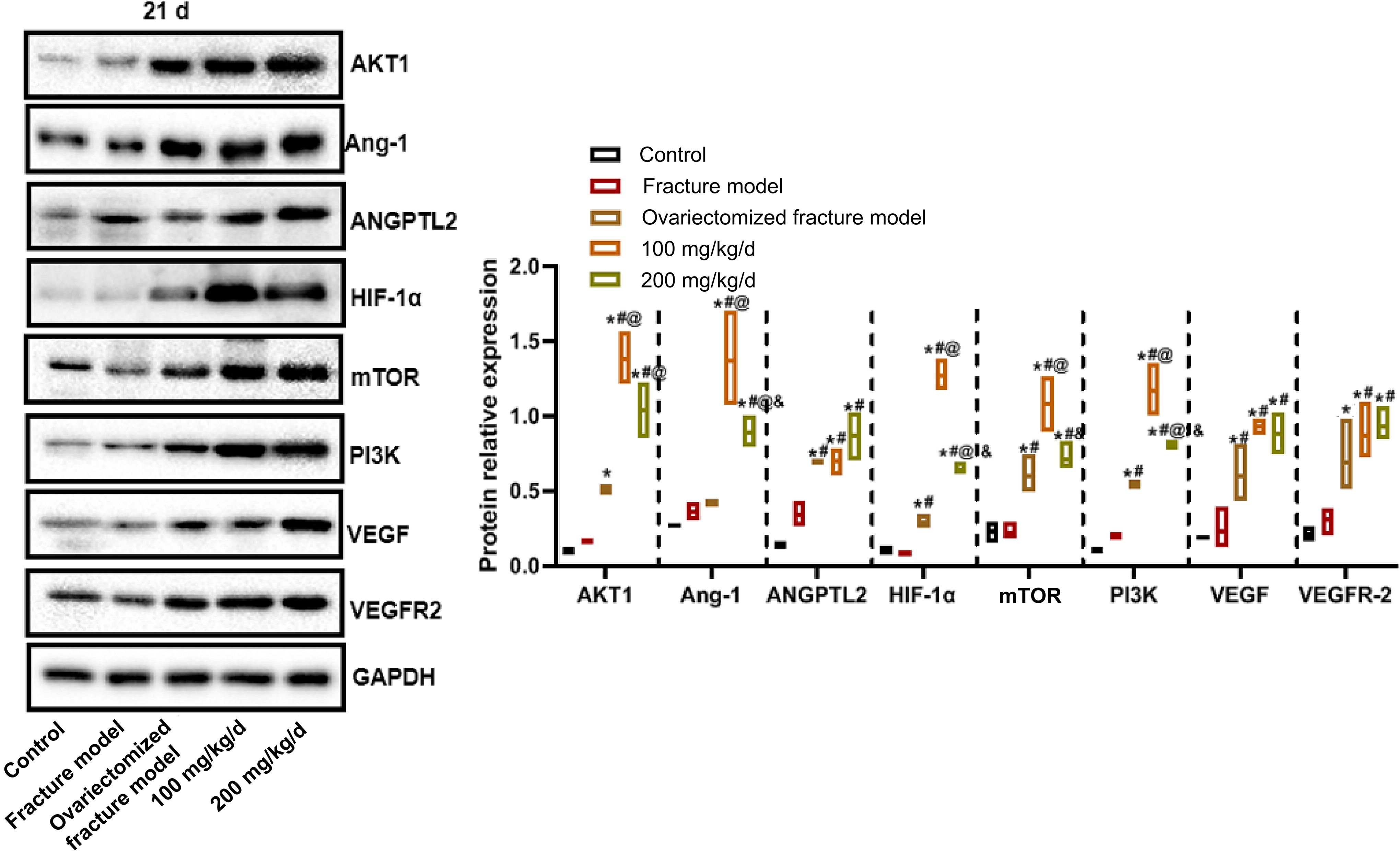
Western blot detection of the protein expressions in callus tissue of rats in each group on the 21st day after modeling.
DISCUSSION
Age, gender, race, and other factors are closely related to the prevalence of osteoporotic fractures. Osteoporotic fractures usually occur in the thoracolumbar spine, hip, proximal humerus and distal radius, and ulna. 18 Among them, hip fracture is the most serious complication of osteoporosis because of its high disability rate and high mortality rate. Vertebral fracture can lead to kyphosis deformity and chronic low back pain; severe vertebral fracture can cause respiratory, digestive, and circulatory function decline. 19 Although these serious complications have caused enormous harm to elderly patients, a large amount of medical resources are consumed for the treatment of osteoporotic fractures in our country each year, which also pose a serious impact on the entire social economy. 20 In this study, a fracture model was established in rats. The female rat ovariectomies caused systemic bone osteoporosis. The osteoporosis model could be established 1 month after ovariectomy. The fracture was induced by external force. Osteoporotic fracture is one of the most common and serious clinical complications, and its repair is difficult; thus, we explored it in this study. A study by Inoue et al. described an establishment of femoral bone defect model in ovariectomized rats, 17 which is consistent with the idea of our composite model, to explore the treatment of fracture after osteoporosis. In this study, the cortical bone area of BV in the ovariectomized fracture model was decreased, and the callus area was increased, suggesting that ovariectomy-induced bone loss and fractures that mimics those in osteoporosis, and stimulated callus formation, which was improved by PNS treatment.
PNS can improve the cerebral blood circulation of the cerebral vascular system to prevent cerebral hemorrhage caused by cerebral embolism. 21 It facilitates hemostasis; activates blood circulation; induces platelets to release Adenosine diphosphate (ADP), platelet factor, Ca2+, and other hemostatic active substances; and ultimately shows procoagulant effects. It also enhances the reduction of blood viscosity and improves the “hypercoagulable state” of the blood. It also has a strong blood activating effect. It can proliferate hematopoietic cells and increase their binding activity with the promoter and enhancer of the upstream regulatory region to regulate the gene expression related to the hematopoietic cell proliferation and differentiation. PNS also has anti-inflammatory, antipulmonary, antiliver and renal interstitial fibrosis, antiaging, and antitumor effects and promotes osteogenesis. It can improve bone mineral density, osteoblast activity, and promote fracture repair. 15,22 –24 In the present study, the number of blood vessels and vascular area in the low- and the high-dose PNS groups increased significantly compared with the ovariectomized fracture model group. PNS promoted fracture healing and increased the number of blood vessels and vascular area. Therefore, PNS promoted angiogenesis. Few studies reported on the toxicity of PNS. Wang et al. showed that by affecting the tricarboxylic acid cycle, energy metabolism, and arachidonic acid metabolism, PNS can effectively reduce periplocin-induced cardiotoxicity, indicating that PNS is nontoxic. 25 In this study, the dose was determined based on a previous study. 26
VEGF is a multifunctional cytokine produced and secreted by megakaryocytes, platelets, endothelial cells, osteoblasts, or tumor cells. It is a very specific vascular endothelial cell mitogen, which can promote endothelial cell proliferation, angiogenesis, increase vascular permeability, and vascular maintenance. 27 The VEGF signaling pathway is essential for the entire process of angiogenesis, and it influences the proliferation, migration, and chemotaxis of vascular endothelial cells in the bone, lung, kidney, brain, and tumor. 28 Under the pathological state, hypoxia is the most important factor that promotes the synthesis of VEGF. Under the hypoxia state, the synthesis of VEGF in cells increases by 12 times. Cell hypoxia causes the release of HIF-1 and then promotes transcription of the VEGF gene. 29 VEGF is expressed at different stages of fracture healing. 30 In the early stage of fracture, many active VEGFs are observed in the fracture hematoma and are significantly higher than those in the systemic blood circulation. 31 During fracture healing, VEGF plays a key role in angiogenesis and bone formation. 32 VEGF has a high affinity with its receptor VEGFR-2, and their combination promotes angiogenesis. 33 In this study, the results showed that VEGF and VEGFR-2 expression increased at different times after the fracture, indicating that VEGF and VEGFR-2 expression was promoted after the fracture. Different doses of PNS further increased VEGF and VEGFR-2 expression in ovariectomized rat fracture model, indicated that PNS may play a role in angiogenesis and bone formation by increasing VEGF and VEGFR-2 expression.
HIF is a nuclear transcription factor with high activity in hypoxia, a key mediator specifically regulating oxygen homeostasis, and a key factor in response to hypoxia stress. 34 Increased in HIF-1 expression is the first adaptive response at the molecular level in the early stage of hypoxia, and it can regulate various downstream target gene expressions. 35 HIF-1α plays a key role in the occurrence of early fracture healing. 36 Different hypoxic environments contribute to the process of bone development, metabolism, and fracture healing, which can activate HIF-1α. Through signal transduction, VEGF, one of direct target genes of HIF-1α, regulates angiogenesis and bone formation. HIF-1α promotes angiogenesis mainly by promoting the expression of VEGF protein under hypoxia. 37 Javaheri B found that HIF-1α participated in cell differentiation with tissue specificity. 38 In this study, HIF-1α expression increased significantly at different times after the fractures in ovariectomized rats, indicating that HIF-1α promoted neovascularization formation by upregulating VEGF content during fracture healing. In addition, different doses of PNS further increased the HIF-1α expression in the ovariectomized rat fracture model, indicating that PNS may upregulate VEGF expression by increasing HIF-1α expression, so as to promote the formation of new blood vessels and chondrocytes, and osteoblasts differentiation, and thus improve fracture healing.
Ang1 is produced and secreted by Sertoli cells such as pericytes and smooth muscle cells around the vascular endothelial cells. Synovial fibroblasts synthesize and secrete Ang1 in rheumatoid arthritis. 39 –41 Ang1 is involved in physiological and pathological angiogenesis in adults. 42 The Ang1 signaling system promoted the process of vascular regeneration induced by VEGF. 43 ANGPTL2 is a secretory glycoprotein. 44 In rat embryos, ANGPTL2 was highly expressed in blood vessels and skeletal muscle, and it affected the function of vascular endothelial cells in the form of autocrine or paracrine resoponses. 45 In this study, the Ang1 and ANGPTL2 expression increased significantly at different time after ovariectomized fracture, indicating that the formation of neovascularization was promoted by upregulating the content of Ang1 and ANGPTL2 during fracture healing. Different doses of PNS further increased the Ang1 and ANGPTL2 expression levels in the ovariectomized fracture model, indicating that PNS may promote the formation of neovascularization by upregulating the expression levels of Ang1 and ANGPTL2.
The PI3K protein family participates in the modulation of cell proliferation, differentiation, apoptosis, and glucose transport and is a key signal transduction molecule in cells. 46 AKT is a serine/threonine-specific protein kinase, which is essential for the growth of various cells, including glucose metabolism, apoptosis, cell proliferation, transcription, and cell migration. 47 The P13K/AKT signaling pathway plays a key function in cell survival, proliferation, and differentiation. 48 AKT is the central link of P13K/AKT pathway and an important downstream molecule of PI3K. 49 mTOR, also known as FKBP rapamycin-associated protein (FRAP), is a member of the phosphoinositide kinase 3-related kinases family and a downstream substrate of PI3K/AKT. 50 PI3K/AKT/mTOR may contribute to the regulation of HIF-1 α expression and activity during hypoxia, but there are some differences in the cell types and experimental conditions. 51 The PI3K/AKT pathway can activate mTOR expression under hypoxia, which leads to an increase in the HIF and VEGF expression. VEGF can directly act on the endothelial cells, promote angiogenesis, and increase vascular permeability. 52,53 In this study, the PI3K/AKT/mTOR expression increased significantly at different time points after the fracture, indicating that the PI3K/AKT/mTOR signaling pathway may involve in the regulation of the healing process of ovariectomized osteoporotic fracture. Different doses of PNS further increased the PI3K/AKT/mTOR expression in the ovariectomized rat fracture model, indicating that in the healing process of ovariectomized rat fractures, PNS may participate in the regulation of the expression and activity of the vascular-related factors via the PI3K/AKT/mTOR signaling pathway. In addition, our results show that the expression of each protein varies from day to day, and we guess it is because the fracture healing process is very complicated and involves many mechanisms. Fracture healing consists of three stages: hematoma inflammation, original callus reaction, and bone growth and remodeling. With the change of time, the expressions of proteins in each stage are different.
Nevertheless, there were some limitations to this study. One of the limitations was that the angiogenic cytokines such as interleukin (IL)-1β, IL-6, IL-8, and IL-10 were not assessed in this study. These should be further explored in subsequent studies. Adding an agonist group or activation of the PI3K/Akt/mTOR pathway can help to confirm the influence of PNS on osteoarthritis in many aspects, and this will also be further explored in the subsequent experimental design.
CONCLUSION
In conclusion, after treating ovariectomized rats with osteoporotic fractures, PNS significantly increased cortical bone area and blood vessels and decreased callus area. The expression of PI3K/AKT/mTOR pathway and angiogenesis-related cytokines further increased. In osteoporotic fracture, PNS can increase the expression of angiogenesis-related cytokines by regulating the PI3K/AKT/mTOR signal pathway, thus promoting the fracture healing of osteoporotic fractures in ovariectomized rats, which shows that PNS is in line with expectations in the treatment of osteoporotic fracture.
Footnotes
ACKNOWLEDGMENT
The authors are grateful to the study participants for their valuable contribution to this research.
AUTHORS’ CONTRIBUTIONS
T.J.: conceptualization, data curation, writing –original draft, formal analysis, writing—review and editing, and visualization. G.H.: Formal analysis, writing—original draft, and visualization. R.Y.: Conceptualization and data curation. Z.G.: Conceptualization, methodology, writing—review and editing.
DATA AVAILABILITY
The data underlying this article will be shared on reasonable request to the corresponding author.
AUTHOR DISCLOSURE STATEMENT
The authors declare no conflicts of interest.
FUNDING INFORMATION
This work was supported by