
Abstract
Ulcerative colitis (UC) is a subtype of inflammatory bowel disease affecting the colon with idiopathic origin. Melinjo endosperm extract (MeE) contains polyphenolic compounds that have antioxidative and anticancer properties. We examined the effect of MeE on inflammation and mucin expression in the colons of UC of mice treated with dextran sulfate sodium (DSS). C57BL/6J male mice were assigned into four categories: control, DSS + 0% MeE, DSS + 0.1% MeE, and DSS + 0.5% MeE. The control group was provided distilled water and a standard chow diet for 4 weeks. In DSS + 0% MeE, DSS + 0.1% MeE, and DSS + 0.5% MeE groups, the mice were treated with MeE for 3 weeks followed by MeE diets and drinking water containing 3% DSS for a week. Macrophage count, the mucus area stained by Alcian blue (AB), the levels of adenosine monophosphate-activated protein kinase (AMPK), nuclear factor-κB (NFκB) p65, and silent information regulator (Sirt) 1 protein expression, as well as proinflammatory mediators and Mucin 2 mRNA expression were assessed. In the DSS + 0% MeE group, the AB-stained areas and Mucin 2 mRNA expression levels were observed to be lower than those of controls. However, the levels in the +0.5% MeE group were significantly increased. Compared with the control group, the macrophage number, the expression of IL-1β mRNA, and NFκB p65 protein in the DSS + 0% MeE group showed a significant increase. Conversely, these levels were significantly decreased in the +0.5% MeE group. The phosphorylated AMPK and Sirt1 protein levels were upregulated in the +0.5% MeE group. In conclusion, MeE may alleviate UC injury by reducing macrophage infiltration and regulating the AMPK/NFκB/Sirt1 pathway.
INTRODUCTION
Inflammatory bowel disease (IBD) is a chronic condition characterized by inflammation in the gastrointestinal tract (GIT), including the colon’s mucosa. 1 Ulcerative colitis (UC), a subtype of IBD, presents as a widespread disorder marked by persistent inflammation of the colonic mucosa, accompanied by chronic diarrhea, abdominal pain, rectal bleeding, and colonic shortening. 2,3 While the pathogenesis of UC remains uncertain, it is thought to arise from intricate interactions involving environmental factors, immune dysregulation, and alterations in the gut microbiome. 4
The mucosal layer serves as a protective barrier on the surface of the GIT. Intestinal goblet cells, situated within the epithelial layer, secrete Mucin 2, which plays a vital role in protecting epithelial cells against contact and triggering by intestinal bacteria. Thus, dysfunction of the mucus barrier is inextricably linked to the onset and progression of UC. 5
Adenosine monophosphate-activated protein kinase (AMPK) has been reported to regulate the metabolisms of carbohydrate and fat across different systems including the hepatic and striated muscle systems. 6 Recent studies suggests that AMPK signaling is pivotal in suppressing inflammatory responses, including those observed in colitis 7 and pneumonia. 8 For instance, metformin, an antidiabetic drug, significantly attenuated the severity of colitis through AMPK activation in the intestinal epithelial cells and suppressed the development of colon cancer in mice. 9 In addition, curcumin was reported to inhibit the invasion of colon cancer cell through AMPK activation and subsequent inhibition of the expression of the p65 subunit of nuclear factor-κB (NFκB). 10
Silent information regulator (Sirt) 1, known as a nicotinamide adenine dinucleotide-dependent histone deacetylase, controls physiological processes such as inflammation, oxidative stress, apoptosis, and autophagy. 11 SRT1720, a Sirt1 activator, decreased interleukin (IL)−1β and tumor necrosis factor (TNF)-α mRNA levels and the histological score in the colon of a mouse colitis model. 12
Melinjo (Gnetum gnemon L.) is a plant species native to Southeast Asia frequently used in Indonesian cuisines. Ethanol extract obtained from the endosperm of melinjo comprises stilbenoids, including gnetin C (a resveratrol dimer), gnemonosides A and C (resveratrol dimer glycosides), and trans-resveratrol (a resveratrol monomer). 13 Melinjo resveratrol intake upregulated pathways of Sirt1/AMPK and nuclear factor erythroid-2-related factor (Nrf)-2 mediated antioxidant defense in inflamed gingival tissues. 14 Melinjo endosperm extract (MeE) alleviated hyperglycemia and hyperinsulinemia and inflammatory conditions in adipose tissue of mice treated with high-fat diet. 15 In a previous study, we showed that hepatic AMPK activity was elevated in offspring of pregnant rats administered a high-fructose diet when the mothers consumed melinjo extract during lactation. 16 However, few studies have investigated the action of MeE on inflammation within the colons of individuals diagnosed with UC.
The present study was conducted to assess whether MeE affects inflammation and mucin expression in the colons of mice with dextran sulfate sodium (DSS)-induced colitis. In addition, whether MeE could modulate inflammation through AMPK/NFκB/Sirt1 pathway was investigated in the colons.
MATERIAL AND METHODS
Melinjo seed extract powder
Melinjo seed endosperm powder (Gnetoron®) was provided by Hosoda SHC Co., (Fukui, Japan). Based on the details provided by the supplier, the powder was manufactured with reference to a previous study 13 with slight modifications. In brief, the dried melinjo endosperms were extracted with ethanol solution. The filtrate was evaporated in vacuo, and the concentrate was freeze-dried with emulsifier and starch to obtain the MeE powder. The obtained MeE powder contained 16.8% gnetin C, 1.1% gnemonoside A, 4.4% gnemonoside D, and 0.3% trans-resveratrol.
Mouse colitis model and experimental design
In this study, after approval from the Animal Research Committee of the Aomori University of Health and Welfare, all experimental procedures were carried out complying with Institutional Guidelines for Animal Experimentation (Permission number: 22001). Male C57BL/6J mice were purchased from CLEA Japan Inc. (Tokyo, Japan) and used in this study. All mice were allowed ad libitum access to standard laboratory chow diet and distilled water. The mice were housed at a constant temperature of 23 ± 1°C with a 12-hour light/dark cycle. After one week of adaptation, the mice, weighing between 21 and 24 g, were assigned to four groups in a random manner: control (n = 8), DSS + 0% MeE (n = 8), DSS + 0.1% MeE (n = 8), and DSS + 0.5% MeE (n = 8) (Fig. 1A). The control group was given distilled water and standard chow diet for 4 weeks. In DSS + 0% MeE, DSS + 0.1% MeE, and DSS + 0.5% MeE groups, MeE was treated for 3 weeks followed by MeE diets and water supplemented with 3% DSS solution (molecular weight ranging from 36 to 50 kDa, 160110, MP Biochemicals, Santa Ana, CA, USA) for a week. We chose to administer diets containing 0.1% and 0.5% MeE according to the findings from an assessment for subchronic toxicity in rats, where the level of no observed adverse effect was equivalent to a preparation of 0.5% MeE (1000 mg/kg daily) in this study. 17 Throughout the experiments, the bedding materials in all the cages were changed simultaneously to mitigate the effects of environmental factors. All mice were weighed daily and observed for diarrhea and/or bloody stools on day 6. On day 8 after the DSS-treatments, all mice were weighed, and colons and livers were collected under anesthesia using a combination anesthetic prepared with medetomidine, midazolam, and butorphanol, intraperitoneally. 18 The colon lengths were promptly measured and then subsequently immersed in 4% paraformaldehyde in phosphate-buffered solution for immunohistochemistry. The portion of the colons were immediately frozen in liquid nitrogen and stored at –80°C before measurement.
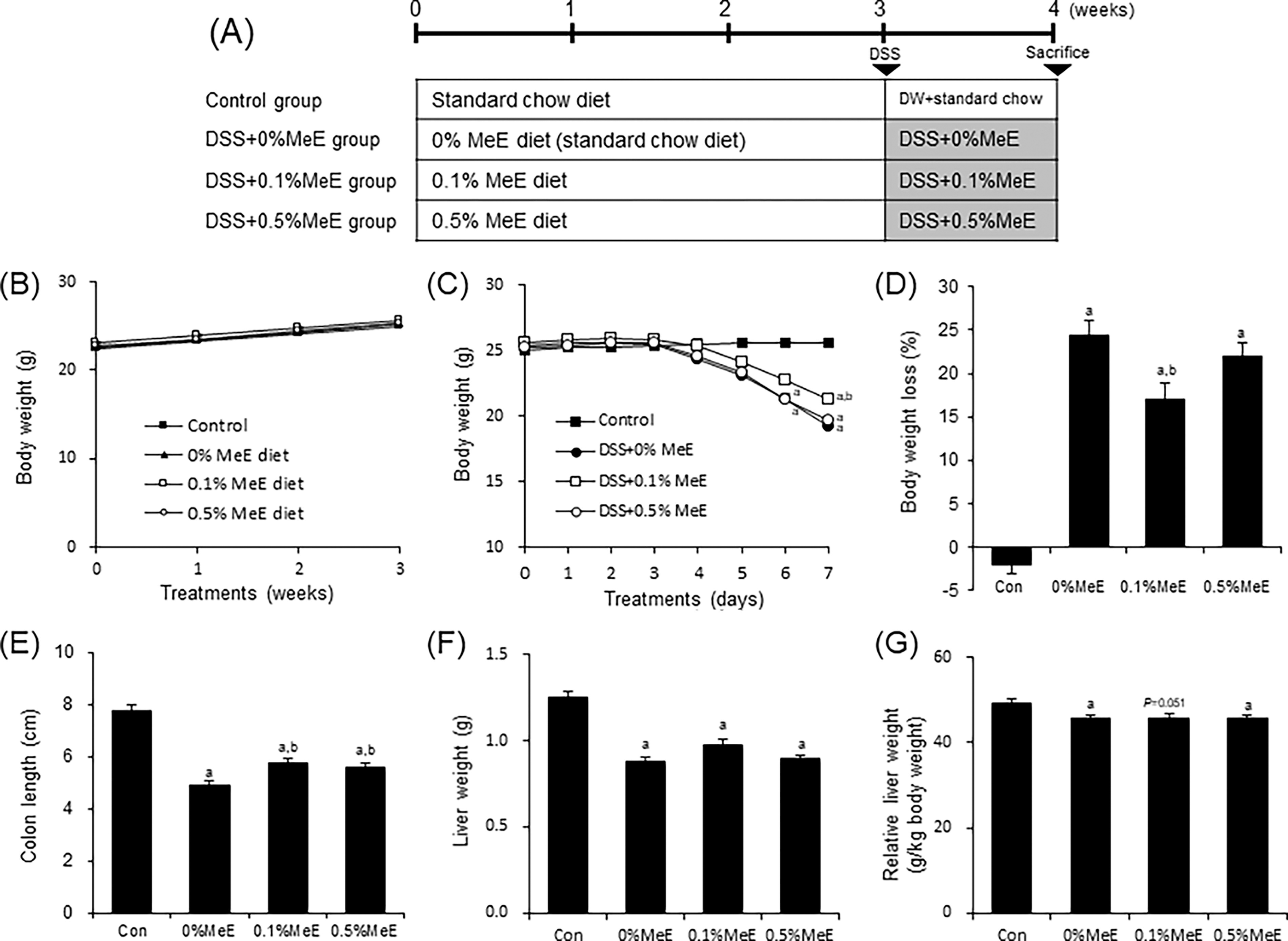
Effect of melinjo extract (MeE) on DSS-induced colitis in mice.
Histopathology and immunohistochemical staining
Hematoxylin-eosin (HE) staining was performed on the 4-μm thick cross sections of the colons.
The histological damage the colons was scored in a grading scale unbiasedly to analyze the histological anomalies or lesions across the entire colon, as previously outlined. 19
To assess the degree of mucin production, the area was stained with Alcian blue (AB). The AB-stained area was analyzed in 10 randomly selected fields using CellSensDimension software (Olympus Corporation, Tokyo, Japan). The area stained with AB was normalized to the total area of the cross-section, and the results were presented as a percentage.
To assess the distribution of macrophages in the colon, a cross section of the colon was stained for macrophages following the protocol established by Sato et al. 20 In brief, the cross sections were pretreated with pepsin solution and incubated with anti-F4/80 antibody (Bio-Rad, CA, USA), followed by the incubation with the reagents of Histofine® Simple Stain Mouse MAX PO (Nichirei Biosciences Inc., Tokyo, Japan). The positive reactions were detected using a solution of 3,3′-diaminobenzidine tetrahydrochloride. Ten fields were randomly selected to analyze the F4/80-positive cells in the colon. The quantification was conducted utilizing CellSensDimension software (Olympus Corporation).
Western blotting
Colon tissues were homogenized in RIPA buffer supplemented with 1 mM phenylmethylsulfonyl fluoride and a protease inhibitor cocktail (Roche Applied Science, Indianapolis, IN, USA). After centrifugation, the supernatants were harvested for analysis of protein concentration. The proteins were separated using electrophoresis on 10% sodium dodecyl sulfate-polyacrylamide gels and subsequently transferred onto PVDF membranes.
Following transfer, the PVDF membranes were incubated with antibodies specific to targeted proteins, such as AMPKα, phospho-AMPKα-Thr172 polyclonal antibody (1:500), mTOR (1:500), phospho-mTOR-Ser2448 (1:500), NFκB p65 (1:500, Cell Signaling Technology, Danvers, MA, USA), Toll-like receptor (TLR) 4 (1:500, Santa Cruz Biotechnology, Santa Cruz, CA, USA), and Sirt1 (1:500, Abcam, Cambridge, UK). The proteins bands were visualized using enhanced chemiluminescence reagents for western blot detection, and the density of specific bands was subsequently quantified by a densitometry software (ATTO Corp., Tokyo, Japan). The levels of targeted protein were normalized to the levels of β-actin in each respective sample.
Real-time PCR
Total RNA was extracted from the colon tissue following the manufacturer’s instructions of Direct-zol RNA MicroPrep Kit (Zymo Research, CA, USA). Complementary DNA was synthesized using a High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher Scientific, Vilnius, Lithuania). The levels of TNF-α, IL-1β, and β-actin mRNA were quantified using specific primers designed for each target gene as follows: forward: GCCTATGTCTCAGCCTCTTCT and reverse: TTGTGAGTGTGAGGGTCTGG for TNF-α; forward: AGGCAGTATCACTCATTGTGG and reverse: ACGAGGCTTTTTTGTTGTTC for IL-1β; forward: GCTGACGAGTGGTTGGTGAATG and reverse: GATGAGGTGGCAGACAGGAGAC for MUC2; and forward: CACGATGGAGGGGCCGGACTCATC and reverse: TAAAGACCTCTATGCCAACACAGT for β-actin. Real-time PCR was conducted using a StepOnePlus™ instrument (Applied Biosystems, Foster city, CA, USA) and the PowerUp™ SYBR™ Green Master Mix (Thermo Fisher Scientific) following the manufacturer’s instructions. The levels of gene expression were expressed relative to those of β-actin levels. The fold differences were determined using the delta–delta CT method with the StepOne™ software.
Statistical analysis
The data are presented as means ± standard errors of the means (SEM), and statistical analyses were conducted using one-way analysis of variance followed by Tukey’s post hoc test. Statistical significance was defined as a P value less than .05 in all cases.
RESULTS
Body weight changes and colon length
Before DSS treatment, the body weights were not significantly different among the MeE-untreated, the 0.1% MeE, and the 0.5% MeE groups for three weeks (Fig. 1B). Following DSS treatment, mice in the DSS-treated groups showed significantly lower body weights compared with the control mice by day 7 (Fig. 1C). Conversely, in the DSS + 0.1% MeE group, the body weights were significantly heavier compared with the +0% MeE group. The body weight loss in mice of the DSS + 0% MeE group was higher than that in mice of the control group (Fig. 1D). The weight loss of mice in the +0.1% MeE group was significantly reduced compared with that of mice in the +0% MeE group, but not in the +0.5% MeE group. Incidences of diarrhea and/or bloody stools were observed on day 6, with varying occurrences across groups: control (0/8), DSS (3/8), DSS + 0.1% (3/8), and DSS + 0.5% (5/8) (observations/all animals). The colon lengths of mice in the DSS + 0% MeE mice were significantly shortened compared with those in the control mice. Conversely, the colon lengths of mice in the +0.1% MeE and +0.5% MeE groups were significantly longer compared with the 0% MeE mice (Fig. 1E). Liver weights (absolute and relative) of mice in the DSS-treated groups were significantly lower than those of the control group. However, the weights of the livers were not significantly different among the +0%, +0.1%, and +0.5% MeE groups (Fig. 1F and G).
Effect of MeE on Histopathological Score, AB-Stained Areas, and MUC2 mRNA
In the HE-stained sections, the histopathological scores in the colons of the +0% MeE group were significantly elevated compared with the control group, confirming that DSS treatment caused severe colon injury characterized by crypt structure and goblet cell damage, muscular thinning, and intense infiltration of inflammatory cells (Fig. 2), consistent with typical UC symptoms. In contrast, DSS + 0.1% MeE and +0.5% MeE groups showed significantly lower scores compared with those in the +0% MeE group.
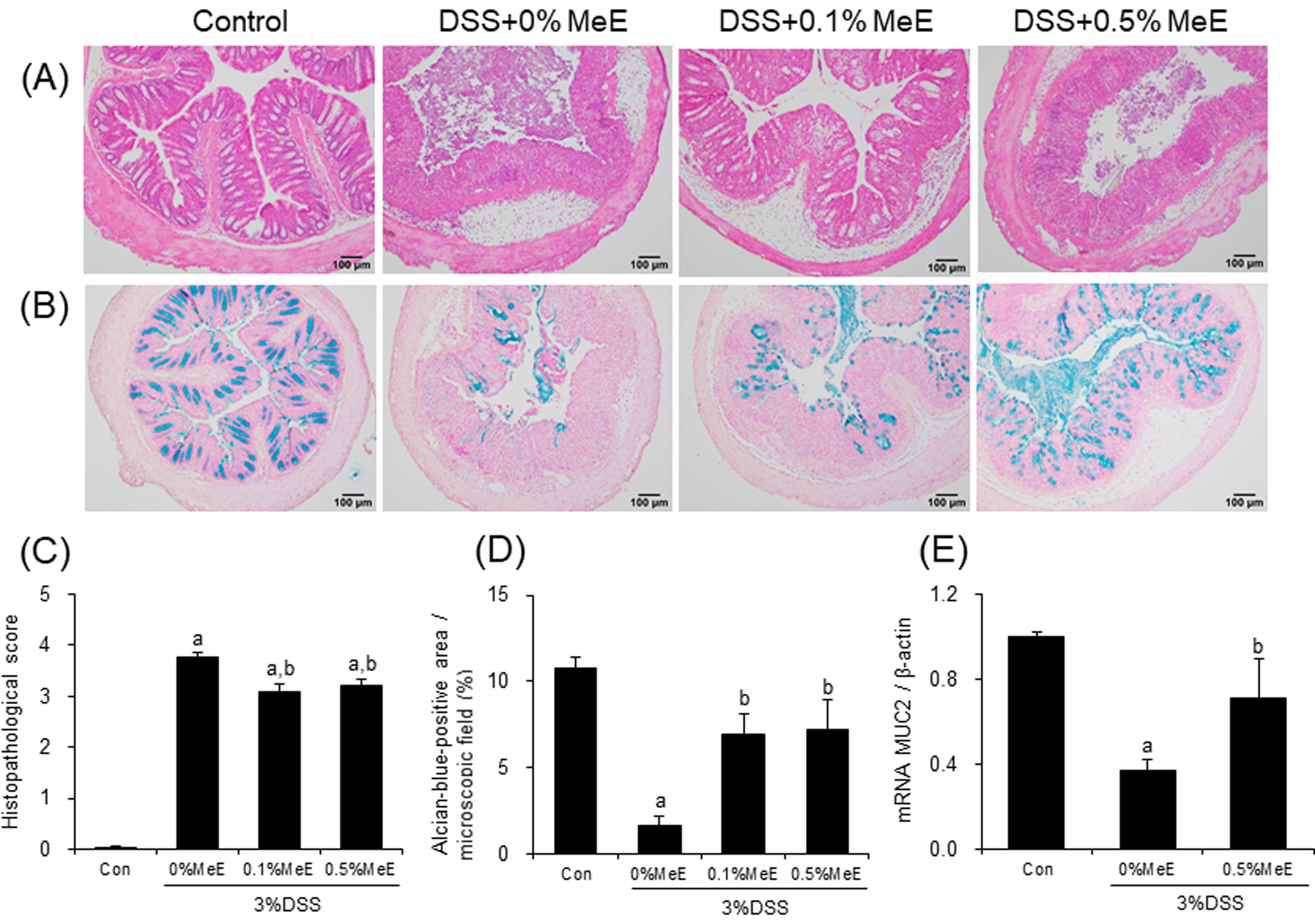
Histopathological staining for hematoxylin-eosin (HE) and Alcian Blue (AB), and mRNA expression of MUC2 in the colonic tissues. Representative photomicrographs for
The AB-stained positive areas in the colon of the DSS + 0% MeE group were significantly decreased compared with the control group (Fig. 2). In contrast, in the DSS + 0.1% MeE and +0.5% MeE groups, the positive areas were significantly increased when compared with those in the +0% MeE group. However, the AB-stained areas were not significantly changed between the DSS + 0.1% MeE and +0.5% MeE groups.
To assess the effect of MeE on mucin production in the colon, the mRNA levels of MUC2, a secreted form of mucin, were evaluated. MUC2 mRNA levels in the colon of the DSS + 0% MeE group were significantly decreased compared with the control group. Conversely, the levels of MUC2 mRNA were significantly elevated in the +0.5% MeE group (Fig. 2).
Effect of MeE on macrophage infiltration and mRNA levels of proinflammatory cytokines
Immunohistochemical analysis revealed the presence of macrophages that reacted positively for F4/80 in the inflamed mucosal lamina propria of the colon in DSS-treated mice (Fig. 3). The quantity of F4/80-positive macrophages in the colons of the DSS + 0% MeE group was significantly increased, when compared with the control group (Fig. 3). In contrast, in the DSS + 0.1% MeE and +0.5% MeE groups, the numbers of macrophages were significantly decreased compared with the +0% MeE group.
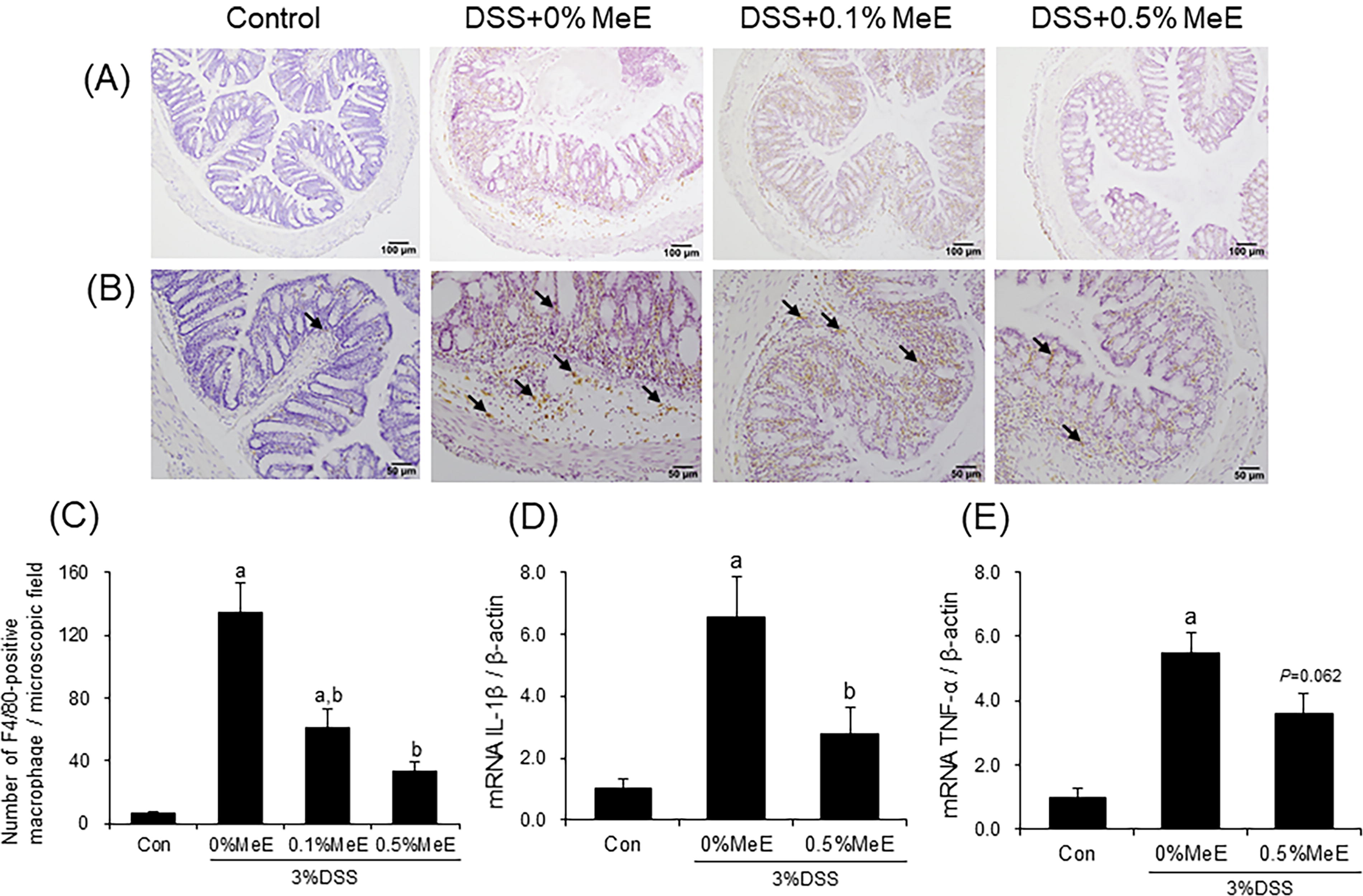
Immunohistochemical staining for F8/40-positive macrophages and mRNA expression of IL-6 and TNF-α in the colonic tissues. Representative photomicrographs of the colonic tissues
To assess the impact of MeE on the production of proinflammatory cytokines in the colon, the expression levels of IL-1β and TNF-α mRNA were examined. The levels of IL-1β and TNF-α mRNA expression in the colons of the +0% MeE group were significantly increased, when compared with the control group (Fig. 3). In contrast, levels of the IL-1β mRNA in the DSS + 0.5% MeE group were significantly decreased, when compared with the +0% MeE group. The DSS + 0.5% MeE group showed a tendency toward decreased expression levels of TNF-α mRNA compared with the +0% MeE group (P = .062).
Effect of MeE on protein expression and phosphorylation of AMPK and mTOR
Although the expression of AMPK protein did not show a significant difference among the colon samples from the experimental groups, the phosphorylated levels of AMPK were significantly increased in the DSS + 0.5% MeE group compared with the +0% MeE group (Fig. 4). Moreover, in the +0.5% MeE group, the phosphorylated to total AMPK (p-AMPK/t-AMPK) ratio was significantly increased. The phosphorylated mTOR levels were increased in the colons of DSS-treated mice; however, significant changes in phosphorylated mTOR levels were not observed among the +0%, +0.1%, and +0.5% MeE groups (Fig. 4). Regarding the expression of total mTOR protein, it was significantly increased in the +0% MeE group compared with the control group. In contrast, the expression of total mTOR in the +0.5% MeE group tended to be lower than that in the control (P = .151) (Fig. 4). The phosphorylated to total mTOR (p-mTOR/t-mTOR) ratios were not statistically related in the colons of the +0%, +0.1%, and +0.5% MeE groups, suggesting that mTOR activation in the colons of DSS-treated mice may be unaffected by MeE treatments.
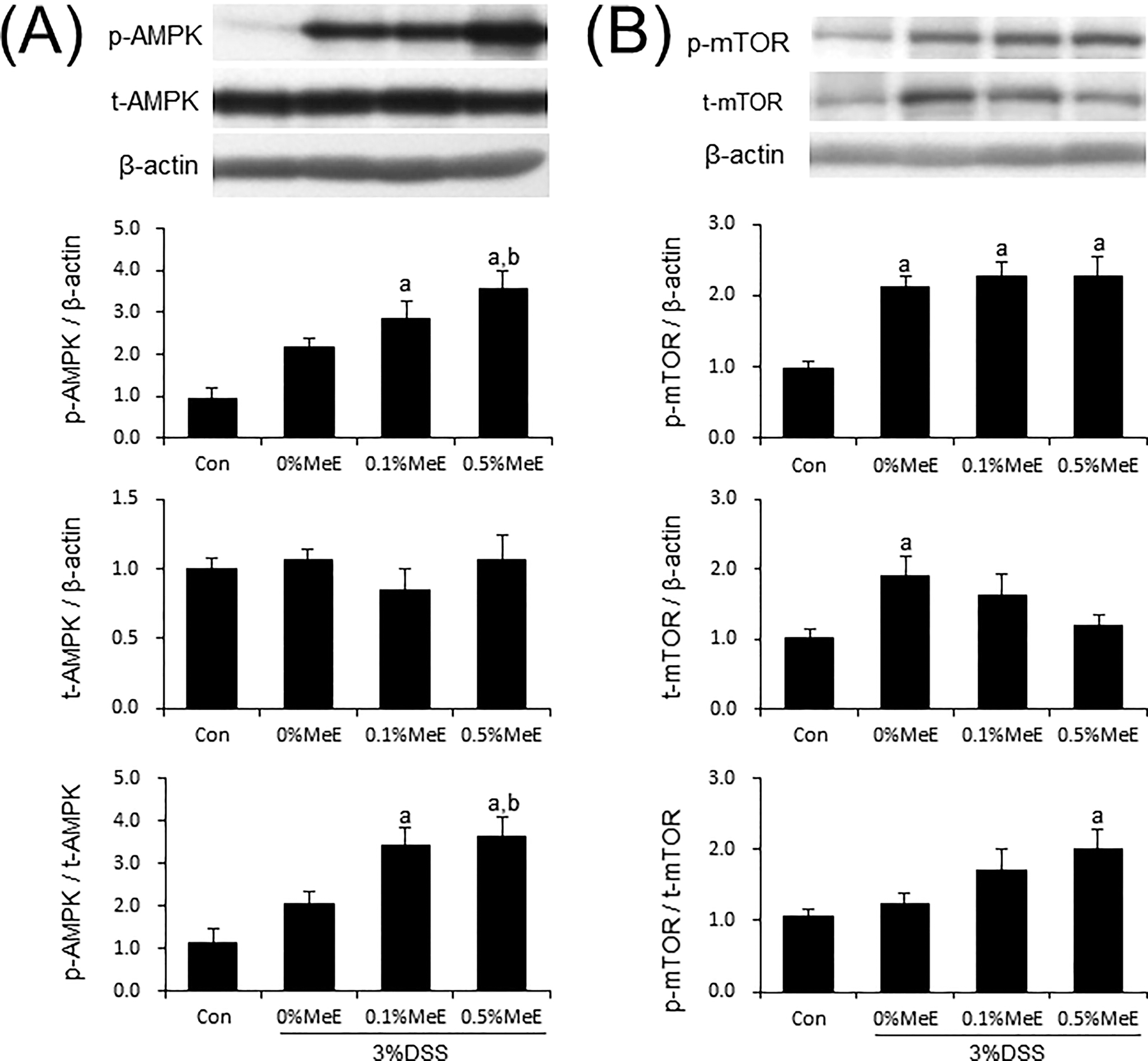
Effect of Melinjo endosperm extract (MeE) on expression and phosphorylation of
Effect of MeE on NFκB p65, Sirt1, and TLR4 protein expression
Compared with the control group, the NFκB p65 expression levels in the colon of the DSS + 0% MeE group were significantly increased (Fig. 5A). Conversely, in the +0.5% MeE group, the expression levels were significantly decreased, when compared with the +0% MeE group. Sirt1 expression levels were significantly decreased in the colon of the +0% MeE group compared with the control group. Conversely, the expression levels in the +0.5% MeE group were notably elevated (Fig. 5B). Colon TLR4 expression levels in the colon were not significantly different among the experimental groups.
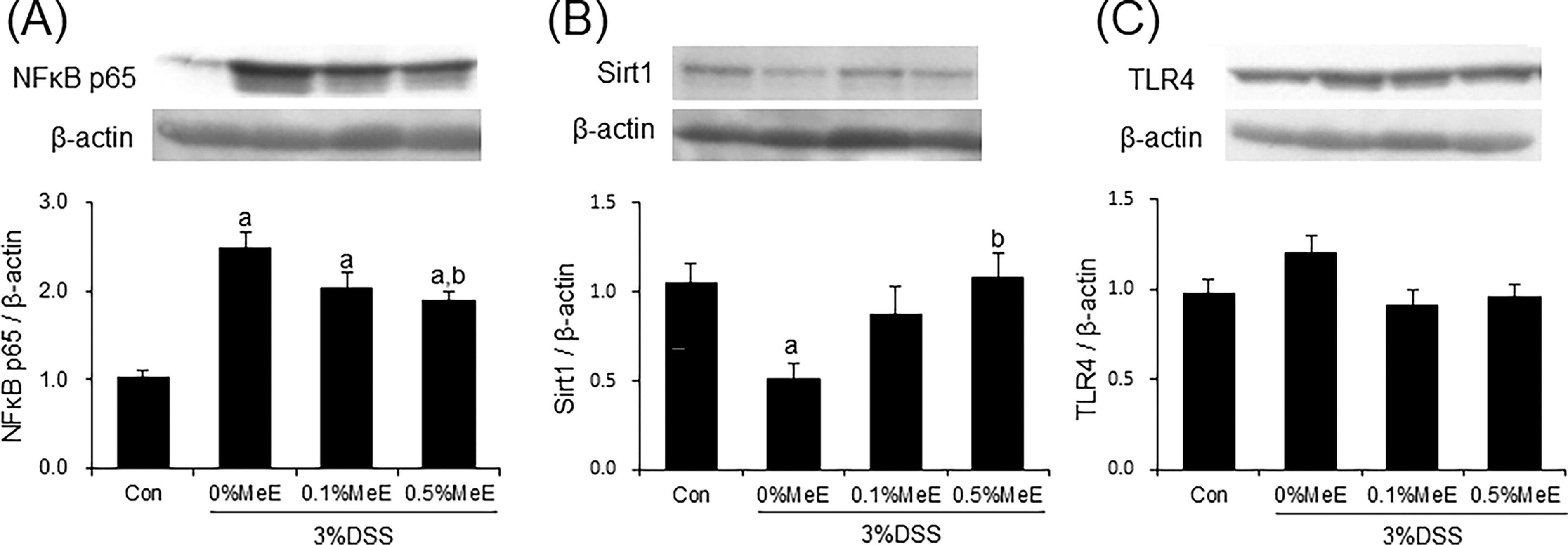
Effect of Melinjo endosperm extract (MeE) on
DISCUSSION
The key outcomes in our study revealed that when mice with DSS-induced colitis were fed diets containing MeE, several significant changes occurred: (1) AB-stained areas and mRNA levels of MUC2 were increased, (2) macrophage count and TNF-α and IL-1β mRNA levels were decreased, (3) AMPK phosphorylation was upregulated, and (4) NFκB p65 protein was downregulated, whereas Sirt1 protein was upregulated in the colon compared with the findings in the 0% MeE group.
In this study, we found that the AB-stained areas and the mRNA levels of MUC2 were elevated in the DSS + 0.5% MeE group compared with the +0% MeE group. The intestinal mucus plays a crucial function in shielding epithelial surfaces from pathogens. The dysregulation of intestinal mucus and goblet cells that produce mucus is associated with infectious and inflammatory conditions. 21 Natural polyphenols from plants protect the intestinal epithelial barrier. For instance, thyme (Thymus vulgaris L.) polyphenols have been reported to mitigate intestinal barrier damage in DSS-triggered colitis in mice. 22 In addition, resveratrol promotes MUC2 synthesis preventing mucosal barrier dysfunction. 23 Therefore, our results suggest that MeE attenuated mucosal damage through the restoration of MUC2 synthesis.
In this study, we demonstrated a decreased macrophage number and lower mRNA levels of IL-1β and NFκB p65 expressions in the MeE-diet fed mice with DSS-induced colitis. Because the activation of NFκB increases the proinflammatory cytokine production and immune cell stimulation in the lamina propria, its transcription factor is essential for stimulating the inflammation cascade. 24 Resveratrol exerts anti-inflammatory activity by decreasing the activation of NFκB. For instance, resveratrol attenuated IL-1β-induced inflammation via the suppression of TLR4/NFкB pathways in cultured human cells. 25 Furthermore, resveratrol treatment reduced the production levels of IL-1β and TNF-α in mice with chronic colitis induced by DSS. 26 Administration of gnetin C (a resveratrol dimer) found in the MeE decreased in IL-1β levels within periodontal tissues in a mouse model of periodontitis. 27 It remains unclear whether the polyphenolic compounds, gnetin C or resveratrol, exhibit more effective actions. Prior studies demonstrated that the activities of gnetin C and resveratrol were nearly equivalent against tyrosinase activity and melanin biosynthesis, 28 suggesting that gnetin C and resveratrol might have a similar effect. Therefore, we assumed that the MeE used in this study may reduce the proinflammatory cytokine synthesis including IL-1β and TNF-α by inhibiting NFκB activation.
Notably an increase in AMPK phosphorylation was detected in the colons of mice treated with DSS + 0.5% MeE. AMPK signaling is a vital role in carbohydrate and fat metabolism across different systems including the hepatic and striated muscle system, and it contributes to protect the integrity of the intestinal epithelial barrier. Metformin upregulates AMPK activity in intestinal epithelial cells and suppresses colon cancer progression in mice. 9 In addition, polyphenolic compounds from natural sources upregulate the AMPK activity. For instance, resveratrol ameliorated intestinal barrier dysfunction through the AMPK pathway in mice with colitis. 23 A resveratrol dimer such as ε-Viniferin has shown elevated levels of AMPK phosphorylation in the livers of rats with diabetes. 29 Thus, in this study, we postulated that MeE activates AMPK and may contribute to maintaining the stability of the epithelial lining of the intestines of mice with DSS-induced colitis.
The reason for MeE attenuating inflammation in DSS-induced colitis is presently speculative. We demonstrated that MeE increased AMPK phosphorylation levels and decreased NFкB p65 protein expression levels in the colons. AMPK regulates the polarization of macrophages toward pro- and anti-inflammatory phenotypes. In lipopolysaccharide (LPS) stimulated RAW264.7 cells, LPS promoted the polarization of M1 macrophages through the NFκB pathway and expression of proinflammatory mediators such as TNF-α and inducible nitric oxide synthase. 30 LPS promoted the polarization of macrophage toward the M1 phenotypes through the NFκB pathway in mouse RAW264.7 cells. 30 In addition, extracellular matrix protein 1 facilitated the polarization of M1 macrophages, which is crucial for managing inflammation and tissue repair in the intestine during IBD in response to LPS stimulation. 31 AMPK is involved in promoting the anti-inflammatory characteristics of macrophages in metabolic disorders such as diabetes and cardiovascular diseases. 32 For instance, metformin has been reported to regulate the AMPK pathway to inhibit NOD-like receptor family pyrin domain-containing 3 (NLRP3) inflammasome activation, thereby promoting M2 macrophage polarization. 33 Resveratrol reduced inflammation in skeletal muscle induced by obesity by decreasing the polarization of M1 macrophages through increased AMPK phosphorylation. 34 Thus, MeE may contribute to the attenuation of inflammation in DSS-induced colitis via regulating macrophage polarization with the activation of AMPK; however, further experiments are required to elucidate the detailed mechanism involved in macrophage polarization. Moreover, AMPK activation suppresses NFκB activation through the increased Sirt1 and cAMP response element binding protein expression, consequently, inhibiting the polarization of M1 macrophages. 35 In this study, MeE-mediated upregulation of Sirt1 expression and the subsequent activation of AMPK may be involved in the mitigation of DSS-induced colitis. Collectively, our findings indicate that MeE may regulate the AMPK/NFκB/Sirt1 pathway to reduce colonic inflammation in mice with colitis induced by DSS.
In conclusion, we show that MeE suppresses the colonic macrophage infiltration and the NFκB p65 expression while enhancing AMPK phosphorylation and Sirt1 expression of mice with DSS-triggered colitis. Furthermore, MeE restored the areas of AB-stain and the mRNA levels of MUC2 in the colon. Therefore, we suppose that MeE may exert beneficial effects on the improvement of UC injury by the regulation of the AMPK/NFκB/Sirt1 pathway as well as the protection of the intestinal epithelial barrier function. Although more detailed studies are necessary to further elucidate the physiological action, MeE may be beneficial for the management of UC if the current results can be replicated in humans.
Footnotes
ACKNOWLEDGMENT
The authors thank Keiko Tamakuma for her technical assistance.
AUTHORS’ CONTRIBUTIONS
S.K.: Conceived and designed the analysis, collected the data, performed the analysis, and mainly wrote the paper. A.K.: Conceived and designed the analysis and collected the data. S.S.: Conceived and designed the analysis, drafting the work and reviewing it critically, and wrote the paper partially.
AUTHOR DISCLOSURE STATEMENT
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
FUNDING INFORMATION
This work was supported by the Aomori University of Health and Welfare under Grant of Aomori University of Health Sciences Senryaku-kata No. Senryaku-kenkyu 7.