
Abstract
Background:
Imbalances in gut microbiota and subsequent destabilization of intestinal barrier equilibrium have been related to the evolution of metabolic disorders. Goji berries (Lycium barbarum; GB) and their fermented counterpart (FGB) have been identified for their prebiotic capacity in managing intestinal barrier functions and inflammatory profiles Consequently, this research was designed to investigate the effects of supplementing GB and FGB on intestinal integrity, inflammation, and changes in the composition of gut microbiota in high-fat (HF)-fed rats.
Materials and Methods:
Thirty-two male Sprague–Dawley rats (6 weeks old, 8 per group) were divided into four categories based on their weight and provided with either respective diets over a 6-week period: low-fat (LF; 10% of calories from fat), HF (45% of calories from fat), and HF diets supplemented with either GB or FGB at a 2% (w/w).
Results:
Supplementation of GB and FGB resulted in compositional changes in the gut microbiota, denoted by a distinct abundance of Faecalibacterium prausnitzii with GB and Akkermansia muciniphila species with FGB, which have been linked to ameliorated obesity phenotypes and metabolic parameters. These alterations were correlated with enhancements in gut barrier integrity, thereby protecting against local and systemic inflammation induced by a HF diet. Supplementation with GB and FGB also mitigated lipopolysaccharide-induced inflammation through inhibition of its downstream pathway.
Conclusion:
These findings indicate that both GB and FGB supplementation can improve gut barrier function and inflammatory profiles in HF-fed rats via modulation of the microbial composition of the gut, supporting the potential application of GB and FGB in improving gut barrier function and managing inflammation amid metabolic challenges.
INTRODUCTION
The gastrointestinal tract stands out as the most densely colonized organ in the human body, harboring around 100 trillion microbes, about 10 times the total number of human cells. 1 Typically, the gut microbiome engenders a symbiotic relationship with its host, orchestrating favorable impacts on both host physiological and behavioral modalities through endocrine, immune, and neural channels that, in turn, modulate cerebral activities. 2 Notwithstanding its regulatory role in sustaining host well-being, alterations to the gut microbiota—termed “dysbiosis”—and subsequent destabilization of gut barrier homeostasis are implicated in the genesis of metabolic disruptions, suggesting a potentially causative role in various pathologies. 3 A high-fat (HF) diet-induced gut dysbiosis, in particular, is correlated with both intestinal inflammation and a compromised gut barrier integrity, resulting in the circulatory influx of endotoxic lipopolysaccharide (LPS) and ensuing metabolic endotoxemia. 4,5 Research indicates that extended exposure to LPS can intensify intestinal permeability, trigger macrophage infiltration, and stimulate inflammation in both fat and liver tissues, underscoring a plausible causal link between microbiota-propelled inflammation and metabolic disturbance. 5,6 Consequently, substantial focus has been directed toward modulating the composition of the gut microbiota as a strategy to circumvent or ameliorate such metabolic aberrations. 3,7 –9 Particularly, noninvasive dietary approaches such as prebiotics have been shown to correct impaired gut barrier function, reduce inflammation both at the local and at the systemic levels, and ultimately alleviate diet-induced hyperphagic conditions by modifying the microbial composition of the host’s gut microbiota. 3,7,8,10
Considerable focus has been placed on the contribution of berries to the prevention of obesity and its associated metabolic issues. Goji berries (Lycium barbarum; GB), rich in polyphenolic compounds, are recognized for their benefits in boosting gut microbiota activity, potentially serving as prebiotics. They are involved not only in enhancing immunity and addressing obesity, hyperlipidemia, and systemic inflammation caused by oxidative stress but also in significantly influencing the regulation of the intestinal microenvironment and enhancing overall host health and specific intestinal responses through their biological effects, including impacts on gut microbiota and metabolites. 11 –16 Furthermore, fermented GB by probiotics (Lactobacillus and/or Streptococcus sp.) has been demonstrated to increase cellular antioxidant and inflammatory activities, preserve intestinal integrity, and modulate gut microbiota. 14,17 –19
We recently demonstrated that supplementation with GB and fermented GB (FGB) exerts positive metabolic impacts in rats subjected to a HF diet, improving hyperphagic conditions, lipid levels, and the antioxidant capacities. 20 However, we did not examine whether these benefits are linked to alterations in gut barrier function or inflammatory responses, along with shifts in the composition of gut microbiota. This study, therefore, aims to evaluate the effects of GB supplementation on the integrity of the intestinal barrier and inflammation, in connection with changes in the gut microbiota of rats fed an HF diet. Furthermore, we seek to identify any differences in metabolic outcomes when GB is fermented with Lactobacillus pentosus compared with its nonfermented form. Our hypothesis posited that supplementation with GB and FGB would modify gut microbiota composition, which would correlate with improvements in intestinal barrier functions and inflammatory profiles, and that there would be discernible differences in metabolic effects between GB and FGB by fermentation.
MATERIALS AND METHODS
Animals and treatment regime
All in vivo experiments were performed according to procedures approved by the Institutional Animal Care and Use Committee of Sun Moon University (SM-2020-02-01). Thirty-two male Sprague–Dawley rats (6 weeks old from Samtako, Osan, Korea) were single-housed in a temperature-controlled room with 12-h light–dark cycle. After the 1-week acclimation period, the rats were divided into four groups (n = 8/group) and fed diets with low fat (LF; 10% kcal from fat), HF (45% kcal from fat), or HF with GB with (HF/FGB) or without fermentation (HF/GB) at the concentration of 2% (w/w) over a 6-week period; the FGB was produced by fermenting enzyme-treated GB with L. plantarum CB3, as previously described. 20 The decision to use a 2% concentration of GB was guided by findings from previous research studies. 11,21 –23 Following 6 weeks on their assigned diets, the animals were subjected to overnight fasting and subsequently euthanized through inhalation of carbon dioxide. Blood specimens were collected in a sterile container and subsequently centrifuged at 1000 g for a duration of 10 min at a temperature of 4°C, facilitating the collection of serum. Liver, cecum (including contents), colon, and retroperitoneal and epididymal fat pads were dissected and weighed. All samples were snap-frozen and stored at −80°C until future analysis.
Quantitative reverse transcription–polymerase chain reaction
Total RNA was extracted using a commercialized kit (Qiagen, Hilden, Germany) followed by reverse transcription of mRNAs using TOPscript™ RT DryMIX (Enzynomix, Daejeon, Korea). Subsequently, quantitative reverse transcription–polymerase chain reaction (qRT-PCR) was performed using CFX96 RT-PCR system (Bio-Rad, Hercules, CA, USA) and low ROX TOPreal™ qPCR 2X PreMIX (Enzynomics). β-actin (colon tissues) or Gapdh (liver tissues) served as the reference genes. Data were analyzed based on the 2−ΔΔCT method. 24 The sequences of the primers used are listed in Supplementary Table S1.
Biochemical analysis
The concentration of LPS-binding protein (LBP) in serum was determined with sandwich-type rat ELISA kits from Elabscience Biotechnology (E-EL-R0589; Houston, TX, USA). The results were expressed as nanograms per milliliter.
Microbiota DNA sequencing
DNA was isolated from the cecal contents using the ZR Fecal DNA MiniPrep per the manufacturer’s protocol (Zymo Research, Irvine, CA, USA). To generate sequencing libraries, the variable V4 region of the 16S ribosomal RNA gene was targeted, following the (Earth Microbiome Project) EMP protocol. 25 Amplicon sequencing was executed on the Illumina MiSeq system, using V2 chemistry (SeqMatic LLC, Fremont, CA, USA). The MiSeq Reporter Software System was used to accomplish the demultiplexing of pooled samples. Raw sequence datasets were then analyzed using Kraken2. METAGENassist platform 26 facilitated the multivariate statistical analysis. Linear discriminant analysis effect size (LEfSe) was calculated using the online Galaxy platform to identify representative taxa for each designated group. 27
Statistical analysis
The statistical analysis was conducted using GraphPad Prism software (version 9.1.1; La Jolla, CA, USA). A one-way analysis of variance was used for the examination of the majority of the parameters. Fisher’s least-significant-difference test was used for a post hoc analysis. Nonparametric Spearman correlation was used to ascertain correlations between the abundance of the cecal microbiome and inflammatory indicators. A significant difference was determined with a P value <.05. The data were expressed as mean ± standard error of mean.
RESULTS
Effect of GB and FGB supplementation on the composition of the gut microbiota
Gut microbiota were assessed across all phylogenetic levels with an abundance >1% represented (Fig. 1). Both GB and FGB supplementation led to compositional changes in the gut microbiota. The gut microbiota of the HF/GB and HF/FGB groups were observed to aggregate together through principal coordinates analysis, thereby differentiating from both the LF and the HF groups throughout all taxonomic ranks (Fig. 1A). Specifically, GB supplementation led to a significant increase in Aerofaciens species, leading to differential abundance (P < .05) of Collinsella, Coriobacteriaceae, Coriobacteriales, Coriobacteriia, and Actinobacteria at the genus, family, order, class, and phylum levels, respectively, as well as to significant increase in abundance of Dorea, Faecalibacterium, and Sharpea at the genus level. The composition of the gut microbiota in HF/FGB rats was distinguished by Muciniphila species (P < .05), which belongs to Akkermansia, Verrucomicrobiaceae, Verrucomicrobiales, Verrucomicrobiae, and Verrucomicrobia at the genus, family, order, class, and phylum levels, respectively.
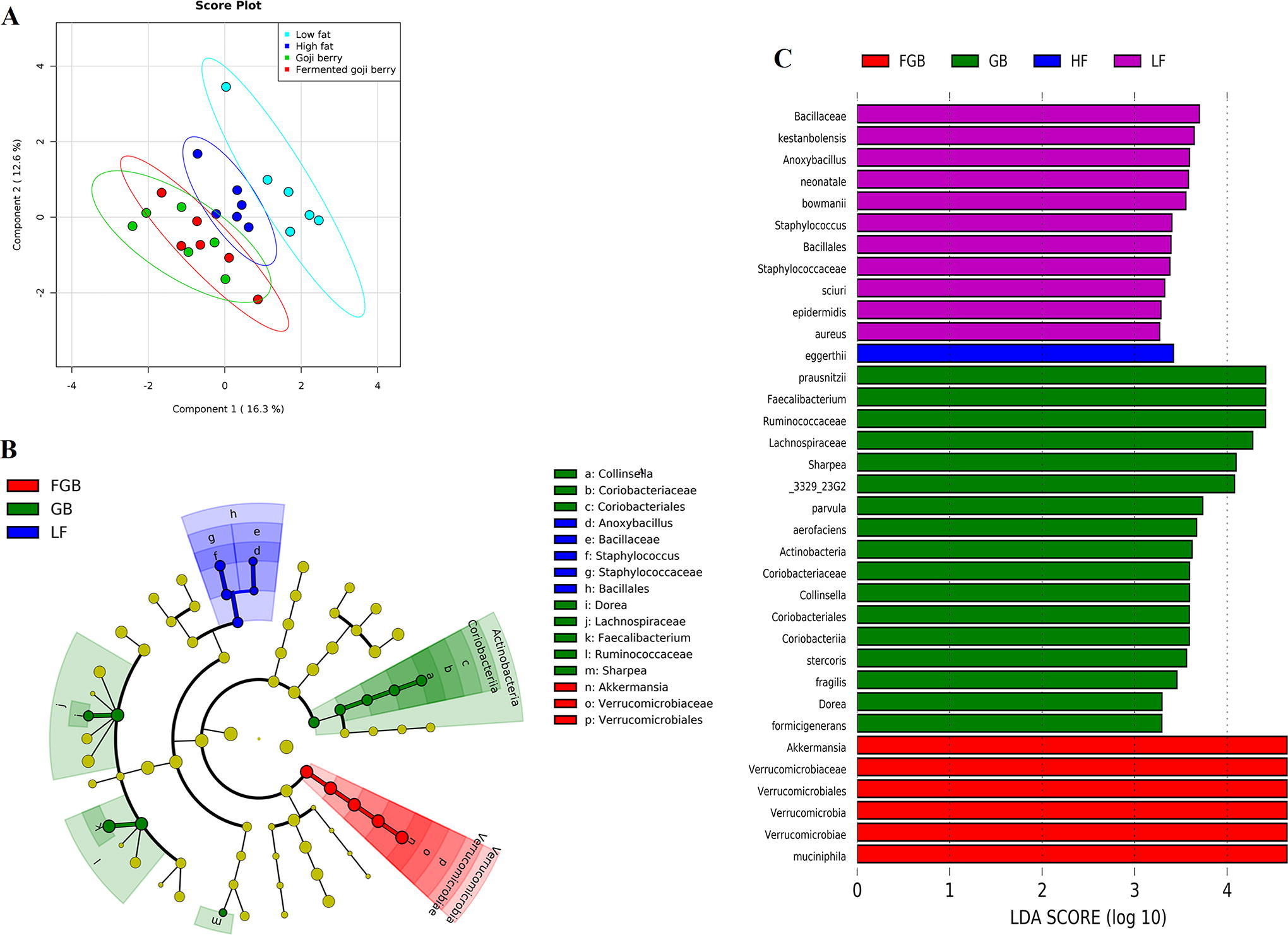
Effect of supplementation of non-fermented and fermented goji berry on the composition of the gut microbiota. Principal component analyses on all taxonomic levels
Effect of GB and FGB supplementation on the intestinal barrier integrity
The permeability of intestinal tight junctions was further examined in the colon (Fig. 2A–C). Gene expression of Cldn-1 and Ocln, as representative tight junction proteins, was significantly upregulated by GB and/or FGB compared with the HF group (Cldn-1: GB vs. HF; Ocln: HF/GB vs. HF at P < .05). Also, the gene expression of zonula occludens-1 (ZO-1) was significantly downregulated by HF feeding (LF vs. HF at P < .05), which was ameliorated by both GB and FGB supplementation.

Effect of supplementing with nonfermented and fermented goji berry on the intestinal barrier integrity. Gene expression of tight junction proteins (Cldn-1, Ocln-1, and Zo-1;
The enhancing impact of GB and FGB supplementation on intestinal barrier function was further validated by notably elevated gene expression levels of mucin 2 (Muc2), Il-22, and regenerating islet-derived 3 gamma (Reg3γ) in comparison to rats fed either the LF or the HF diets, confirming their roles in critical regulation of intestinal homeostasis (Muc2: HF/GB vs. LF or HF; Il-22: HF/FGB vs. HF; Reg3γ: HF/FGB vs. HF at P < .05; Fig. 2D–F).
Effect of GB and FGB supplementation on inflammatory profiles
In the colon, the administration of a HF diet notably elevated the gene expression of Mcp-1 as well as that of pro-inflammatory cytokines, Il-b, Il-6, and Tnfα, compared with LF-fed rats (LF vs. HF at P < .05; Fig. 3A–D). This increase was mitigated by supplementation with either GB or FGB (Mcp-1, Il-b, or Il-6: HF vs. HF/GB or HF/FGB at P < .05; Tnfα: HF vs. HF/GB at P < .05 or HF/FGB at P < .01).

Effect of supplementation of nonfermented and fermented goji berry on HF-induced inflammation at the local and systemic level. Gene expression of inflammatory parameters (Mcp-1, Il-b, Il-6, and Tnfα) in the colon
The HF-induced increase in the circulating level of LBP was significantly reduced by HF/GB and prevented by HF/FGB (HF vs. HF/GB or HF/FGB at P < .05; Fig. 3E).
Effect of GB and FGB supplementation on LPS pathways and subsequent inflammation
In the liver, the gene expressions of TLR4, Cd14, and Myd88, components of the LPS receptor complex, were assessed (Fig. 4A–C). TLR4, Cd14, and MyD88 were significantly upregulated by HF feeding at the gene level, which was suppressed by GB and/or FGB (TLR4: HF vs. LF at P < .05; Cd14: HF vs. LF or HF/FGB at P < .0001 or HF/GB at P = .0001; MyD88: HF vs. LF at P = .0001 or HF/GB or HF/FGB at P < .0001). HF/FGB also downregulated Myd88 gene expression (LF vs. HF/FGB at P < .05).
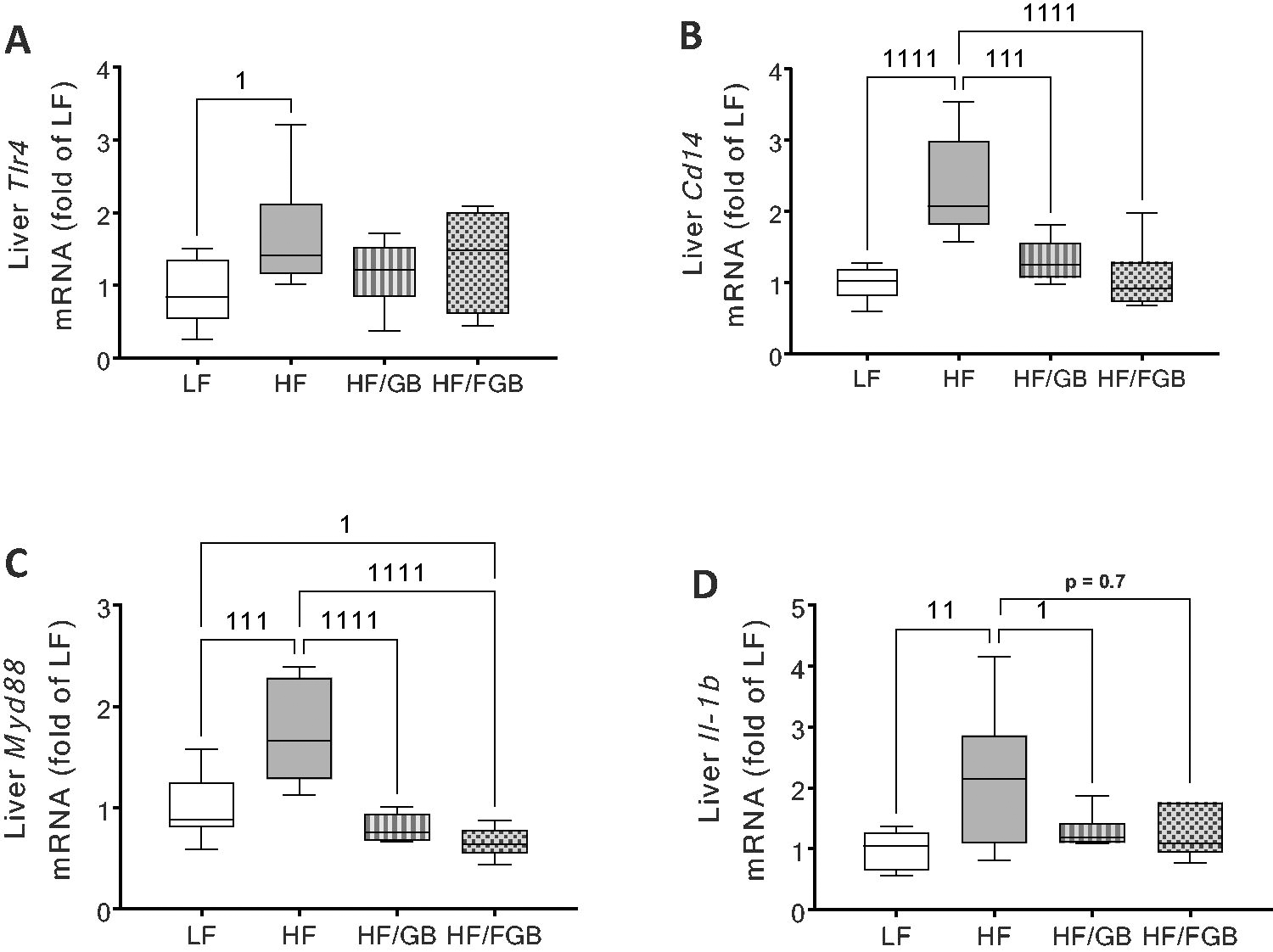
Effect of supplementing with nonfermented and fermented goji berry on LPS receptor complex and downstream inflammation. Gene expression of LPS receptor complex (Tlr4, Cd14, and Myd88;
Similarly, GB and FGB supplementation suppressed HF-induced upregulation of the Il-b gene expression in the liver (LF vs. HF at P < .01, HF vs. HF/GB at P < .05 or HF/FGB at P = .7; Fig. 4D).
DISCUSSION
Here, we explored the effect of GB supplementation on the gut barrier functions and inflammatory markers, in conjunction with changes in gut microbiota composition in rats subjected to a HF diet. In addition, we aimed to discern potential disparities in the pharmacological and metabolic outcomes elicited by FGB with L. plantarum CB3 relative to nonfermented GB. We hypothesized that both GB and FGB supplementation would induce compositional alterations the gut microbiota, correlating with enhancements in intestinal barrier function and inflammation markers. We further anticipated varying magnitudes of pharmacological and metabolic responses between GB and FGB supplementation. Supplementation of GB and FGB resulted in compositional modifications in the gut microbiota, denoted by a distinct presence of Faecalibacterium prausnitzii species with GB and Akkermansia muciniphila species with FGB, all of which have been linked to ameliorated obesity phenotypes and metabolic parameters. 28,29 These alterations were correlated with enhancements in the integrity of the gut barrier, thereby protecting against local (colon, liver) and systemic inflammation induced by an HF diet. Supplementation with both GB and FGB also mitigated inflammation induced by LPS through inhibition of expression of CD14 and MyD88 as its downstream pathway. In summary, these data suggest that supplementation with both GB and FGB contributed to improved intestinal integrity and inflammatory profiles in rats under an HF diet, and these enhancements were related to the compositional changes observed in the gut microbiota.
The primary health benefits of prebiotics were investigated through characterizing the alterations in gut microbiota composition in rats. A marked shift in the composition of the gut microbiota across all taxonomic ranks was noted in rats subjected to HF feeding in comparison with the LF group. Notably, an increased abundance of Bacteroides eggerthii, a species identified as pro-inflammatory, was evident in rats on a HF diet. 30 In addition, supplementation with GB and FGB resulted in notable and distinct modifications in gut microbiota composition compared with both LF and HF diet groups. Notably, GB group exhibited a differential increase in F. prausnitzii species, known for its metabolic benefits. This species is recognized for preserving intestinal mucosal integrity and mitigating gut inflammation. 28,29 A study by Martín et al. indicated that administering F. prausnitzii lessened inflammation severity by augmenting intestinal barrier function and influencing paracellular permeability in a murine model. 28 In addition, Yang et al. showcased that strains of F. prausnitzii could mitigate low-grade inflammation, restore intestinal integrity, reduce liver damage and insulin resistance, adjust hormone release, control appetite, and suppress fat synthesis by influencing the gut–brain axis. 29 On the contrary, FGB supplementation induced a differential abundance of the A. muciniphila species. Specifically, the muciniphila species from the Akkermansia genus, which exhibited differential abundance in the FGB group, has been well recognized for its beneficial role in modulating host metabolism and in addressing obesity and associated metabolic disorders. This is achieved through suppressing systemic inflammation, reducing the process of lipid overload associated with the low-density lipoprotein receptor, enhancing the production of short-chain fatty acids, and/or adjusting intestinal permeability and gut barrier integrity. 31
Consumption of HF diet has been known to induce intestinal inflammation, resulting in compromised gut barrier function and heightened gut permeability. This, in turn, enables the translocation of pro-inflammatory products derived from bacteria, such as LPS, into the circulatory system, as has been documented previously. 4 It was postulated that the alterations in gut microbiota composition driven by GB and FGB could be correlated with enhancements in intestinal inflammation, thereby safeguarding gut epithelial integrity and mitigating systemic inflammation. In alignment with the hypothesis, GB and FGB supplementation normalized the HF-induced escalation of inflammatory responses in the colon. This underscores their potential protective role against localized inflammation, a finding that is in harmony with prior studies. 11,22
Inflammation compromises the functionality of the intestinal epithelial barrier. 4 Consequently, the amelioration of gut inflammation observed in this study, resulting from GB and FGB supplementation, might have conferred protection against the HF-induced deterioration of intestinal barrier integrity. Claudin, occludin, and ZO-1, as the core components of tight junctions, play a vital role in maintaining intestinal barrier function and ensuring the stability of tight junctions. 32 Our study revealed that HF feeding disrupted the permeability of intestinal tight junctions, particularly affecting Zo-1. This disruption was effectively mitigated by GB and FGB supplementation, with GB proving to be especially potent, consistent with previous studies. 11,14 The fortifying influence of GB and FGB on gut barrier function was additionally substantiated by the elevated level of Muc2, Il-22, and Reg3γ genes in the colon, recognized as pivotal regulators of intestinal homeostasis. 33 Cytokines are integral to orchestrating immune responses to microbial and environmental challenges, especially at epithelial barriers. 34 IL-22 stands out as essential for gastrointestinal health, functioning as a primary cytokine in preserving epithelial homeostasis. 33 It’s pertinent to mention that previous studies have linked the release of mucin and the induction of Reg3γ to IL-22. 35 Specifically, the binding of IL-22 to its receptor has the ability to increase mucin production, establishing a physicochemical barrier to prevent pathogen intrusion, and to boost the production of intestinal Reg3γ, which acts as an antimicrobial peptide. 34,35 Therefore, the study suggests that the improvements in intestinal barrier integrity, facilitated by GB and/or FGB supplementation, could be ascribed to the synergistic actions of MUC2 and Reg3γ, which was mediated by IL-22.
Compromised integrity of the intestinal barrier has been known to be linked with systemic inflammation, a process driven by the translocation of LPS across the intestinal epithelial barrier into the bloodstream, thereby triggering widespread inflammation. 4 In correlation with the enhancement in both gut inflammation and barrier integrity, our findings indicate a trend where the increase in serum LBP, as an indicator of circulating LPS resulting from HF feeding, was countered by supplementation with either GB or FGB. In addition, the increased presence of LPS in circulation interacts with its receptor complex (CD14/TLR4/MD2), sparking TLR4-mediated MyD88-dependent signaling, which, in turn, activates a cascade of inflammatory cytokines, including IL-6 and TNFα, in various organs. 36 –38 Significantly, the absence of CD14, a requisite TLR4 adapter protein, in animals prevents the onset of diet-induced inflammation. 5 In the current research, supplementation of GB and FGB demonstrated a mitigating effect on the LPS-induced overexpression of Tlr4 to a certain extent and significantly for Cd14 and Myd88 in the liver. In addition, the related upsurge in the gene expression of the emblematic pro-inflammatory cytokine, Il-b, was attenuated by the supplementation of GB and FGB, thereby substantiating their protective role against systemic inflammation. Supporting this, Ávila et al. have previously documented that the oral intake of GB extract reduced LPS-induced inflammation through the TLR4-dependent pathway in an LPS-challenged rat model. 39
It is worth noting that although GB and FGB supplementation demonstrated metabolic benefits on gut barrier integrity and inflammatory profiles, the HF/FGB group exhibited particularly notable beneficial effects. Research has previously highlighted that the metabolic impacts of prebiotics can vary significantly in both intensity and underlying biological mechanisms when subjected to fermentation with probiotics. 40 –42 Such studies have often shown that fermentation can either amplify the metabolic advantages or introduce novel actions distinct from those of the unfermented prebiotics. This suggests the involvement of alternative biological pathways in the observed metabolic improvements within the HF/FGB group. Therefore, a deeper exploration of various metabolic pathways could unveil the specific actions underlying FGB’s metabolic advantages. In addition, considering the central role of gut microbiota as the primary beneficiary of prebiotics and probiotics’ metabolic effects, using metagenomic and metabolomic approaches could elucidate the differential metabolic mechanisms of GB and FGB.
In conclusion, the current findings demonstrate that supplementation with GB and FGB has the potential to enhance gut barrier functionality and reduce inflammation responses at the local and systemic level in rats fed an HF diet. These impacts were associated with shifts in the composition of gut microbiota, supporting the variations observed in their metabolic advantages as illustrated in this study. Consequently, these observations lend support to the prospective use of GB and FGB in modulating intestinal barrier integrity and the inflammatory profile under metabolic challenges.
Footnotes
AUTHORS’ CONTRIBUTIONS
Conceptualization, methodology, validation, formal analysis, and investigation: E.J., S.E., S.C., and S.L.; data curation: S.E. and S.L.; writing—original draft preparation: E.J.; writing—reviewing and editing: E.J., S.E., S.C., and S.L.; supervision: S.L. All authors have read and agreed to the published version of the article.
DATA AVAILABILITY STATEMENT
Data are available from the corresponding author upon request.
AUTHOR DISCLOSURE STATEMENT
The authors declare no conflict of interest.
FUNDING INFORMATION
This work was supported by the
SUPPLEMENTARY MATERIAL
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.