
Abstract
This study investigated the antiobesity effects of black ginger extract (BGE) in high-fat diet (HFD)-induced obese mice. Mice were divided into six groups: normal diet control (NC, AIN-93G normal diet), 60% HFD control (HFD), HFD containing metformin at 250 mg/kg b.w. (Met, positive control), and HFD containing BGE at 5, 10, or 20 mg/kg b.w. for 15 weeks. BGE administration significantly prevented HFD-induced increases in weight gain, organ weight, and adipose tissue mass. Furthermore, it resulted in decreased adipogenesis and lipogenesis-related factors, including phosphorylated mitogen-activated protein kinase, peroxisome proliferator-activated receptor gamma, CCAAT/enhancer-binding proteins, sterol regulatory element-binding protein 1, phosphorylated cAMP response element-binding protein, glucose-6-phosphate dehydrogenase, fatty acid synthase, dephosphorylated ATP-citrate lyase, dephosphorylated acetyl-CoA carboxylase, and lipoprotein lipase, in white adipose tissues. Moreover, BGE administration enhanced lipolysis in white adipose tissue, as evidenced by elevated levels of adipose triglyceride lipase, phosphorylated hormone-sensitive lipase, and protein kinase A, along with reduced levels of perilipin and phosphodiesterase 3B. BGE induced thermogenesis in brown adipose tissues, as reflected by the increased expression of AMP-activated protein kinase, uncoupling protein 1, and carnitine palmitoyltransferase 1 and decreased levels of fatty acid-binding protein 4. In conclusion, this study provides comprehensive evidence supporting the antiobesity effects of BGE, elucidating the underlying molecular mechanisms involved in preventing weight gain, suppressing adipogenesis, promoting lipolysis, and stimulating thermogenesis. These findings suggest the potential therapeutic utility of BGE in combating obesity and associated metabolic disorders (KHGASP-2023-034).
INTRODUCTION
Obesity, a multifaceted health challenge affecting populations worldwide, has emerged as a critical public health concern. 1 Characterized by excessive fat accumulation, obesity is intricately linked to an array of deleterious health outcomes, spanning cardiovascular diseases, metabolic disorders, and an increased susceptibility to various chronic conditions. 2 –4 Its prevalence has witnessed an alarming surge over recent decades, evolving into a global epidemic that transcends geographical, socioeconomic, and cultural boundaries. 5 –7 Beyond its overt manifestation as a physical condition, obesity represents a complex interplay of genetic, environmental, and lifestyle factors. 8 The fundamental principle underlying changes in body fat is the equilibrium between the calories ingested from food and drinks (energy intake) and the calories expended through the basal metabolic rate, physical exercise, and other processes (energy expenditure). 9,10 The consistent intake of a high-calorie-density food has been linked to an increasing prevalence of metabolic syndrome and obesity. 11,12 The escalating prevalence of high-calorie-density diet consumption poses a significant and pressing global health concern.
As the world grapples with the ramifications of obesity, the pursuit of effective interventions has become paramount. Traditional approaches, often centered around dietary modifications and exercise regimens, have encountered limited success, necessitating a more comprehensive exploration of alternative avenues. 13,14 Natural products derived from plants and herbal sources have surfaced as potential agents in the fight against obesity. 15,16 Investigating their antiobesity efficacy not only holds promise in providing novel therapeutic strategies but also aligns with the growing global inclination toward integrative and holistic healthcare.
Black ginger (Kaempferia parviflora), a plant native to Thailand, is known for its traditional use in Thai folk medicine. 17 It has gained attention in modern research owing to its potential health benefits, including its anti-inflammatory effects and ability to prevent metabolic syndrome. 17,18 Toda et al. discovered that black ginger extract (BGE) contains compounds such as 5-hydroxy-7-methoxyflavone, 5,7-dimethoxyflavone, and 5-hydroxy-3,7,4′-trimethoxyflavone. 19 These polymethoxy flavonoids have been shown to boost energy metabolism in myocytes, suggesting that BGE has the potential to ameliorate metabolic syndrome. In Yoshino et al.’s study, they conducted a randomized, double-blind, placebo-controlled clinical trial with 76 individuals 20–65 years of age and a body mass index of 24–29 kg/m2. Participants were administered 150 mg/day of BGE for a duration of 12 weeks. After the 12-week period, Yoshino et al. confirmed that the abdominal fat area and triglyceride levels showed a significant reduction in the BGE treatment group compared to the placebo group. In addition, they found that none of the groups showed adverse effects or clinically relevant abnormal changes. 20 Therefore, we validated the antiobesity effects of BGE by examining the adipose tissue lipid metabolism of high-fat diet (HFD)-induced obese mice.
MATERIALS AND METHODS
Preparation and standardization of BGE
BGE was provided by Nutrione. The 5,7-dimethoxyflavone content of BGE was quantitatively determined using high-performance liquid chromatography (HPLC). The 5,7-dimethoxyflavone in BGE was extracted using methanol and 1% dimethyl sulfoxide, and it was subsequently filtered before being loaded into the HPLC system. For separation, we used an Agilent Eclipse Plus C18 Column. We determined the amount of 5,7-dimethoxyflavone in BGE using a calibration curve generated based on the concentration of the 5,7-dimethoxyflavone standard against the peak area. The BGE was standardized to 10% 5,7-dimethoxyflavone (Fig. 1).
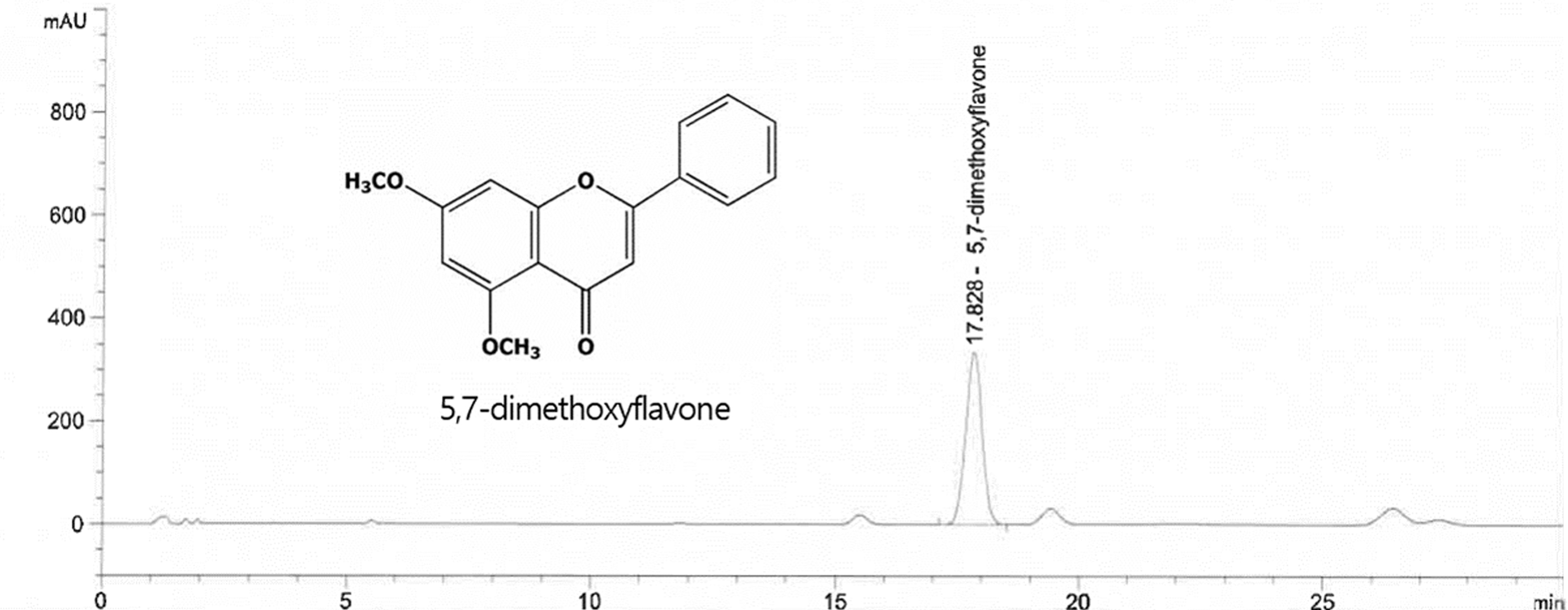
High-performance liquid chromatography analysis of 5,7-dimethoxyflavone in BGE. BGE, black ginger extract.
Animals
Four-week-old C57BL/6 male mice were obtained from Saeron Bio. The mice were maintained in a facility with a 12-h light/dark cycle at 22 ± 2°C and 55 ± 5% relative humidity. All mice were subjected to 1 week of pre-experimental acclimatization, fed standard chow, and provided with fresh water for ad libitum consumption. A total of 48 mice were divided into six groups, randomly assigned: normal diet control (NC, AIN-93G normal diet), 60% HFD control (Con, HFD), HFD containing metformin at 250 mg/kg b.w. (Met, positive control), and HFD containing BGE at 5 mg/kg b.w. (5), 10 mg/kg b.w. (10), or 20 mg/kg b.w. (20) for 15 weeks. Mice were managed in accordance with relevant guidelines and kept under the pre-approved protocols of the University Animal Care and Use Committee (KHGASP-2023-034).
Biochemical analysis
Serum or fecal analyses were performed using a triglyceride quantification kit, cholesterol assay kit, aspartate transaminase (AST) activity kit, and alanine aminotransferase (ALT) activity kit according to the manufacturer’s protocols (Biomax).
Micro-computed tomography
Tomographic scans of the entire body and abdomen were conducted on anesthetized mice under optimal conditions (Voxel size of 150 μm, 45 kVp energy, 110 μA intensity, 79.8 mm field of view/diameter, and 160 ms integration) using micro-computed tomography (micro-CT). Data were analyzed using the micro-CT Evaluation Program V6.6.
Hematoxylin and eosin (H&E) staining
The isolated and fixed tissues from mice were encased in paraffin, then sliced into 5-μm sections from the resulting paraffin blocks, and stained with hematoxylin and eosin. To examine and visualize the sections, an optical microscope was used.
Western blot analysis
Proteins were extracted using a lysis reagent, and 50 μg of each protein sample was loaded into 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis. After being electrophoretically transferred onto membranes, the membrane was blocked using 5% skim milk in Tris-Buffered Saline + Tween 20 (TBST). Afterward, the membranes were incubated while gently shaken with primary antibodies, followed by attachment of the secondary antibody (Table 1). The intensity of western blot bands was quantified using the CS Analyzer 3.0 and then normalized with β-actin.
Antibodies Used for Western Blot Analysis
ACC, acetyl-CoA carboxylase; ACL, ATP-citrate lyase; AMPK, AMP-activated protein kinase; ATGL, adipose triglyceride lipase; C/EBP, CCAAT/enhancer-binding protein; CPT1A, carnitine palmitoyltransferase 1; CREB, cAMP response element-binding protein; FABP4, fatty acid-binding protein 4; FAS, fatty acid synthase; HSL, hormone-sensitive lipase; MAPK, mitogen-activated protein kinase; PDE3B, phosphodiesterase 3B; PKA, protein kinase A; PPARγ, peroxisome proliferator-activated receptor gamma; SREBP1c, sterol regulatory element-binding protein; UCP1, uncoupling protein 1.
Statistical analysis
All data were stated as means ± standard deviations. We assessed the significance of treatment effects using analysis of variance with Tukey’s Honest Significant Difference test after one-way analysis of variance using SPSS software. Statistical significance was set at P < .05.
RESULTS
BGE administration prevented obesity in HFD-fed mice
HFD feeding induced increases in weight gain, food efficiency ratio, and organ weight compared with normal-diet feeding in mice. However, dietary supplementation with BGE significantly reduced weight gain and organ weight, including that of adipose tissue. No significant difference in the food efficiency ratio was noted between the HFD control and BGE groups (Table 2). These findings indicate that supplementation with BGE in the diet can prevent obesity during HFD intake.
Effects of Black Ginger Extract on Body Weight, Weight Gain, and Organ and Adipose Tissue Weight in Mice with High-Fat Diet-Induced Obesity
Normal diet control (NC, AIN93G normal diet), 60% high-fat diet (HFD) control (Con, 60% HFD), HFD containing metformin at 250 mg/kg b.w. (Met, positive control), HFD containing black ginger extract (BGE) at 5 mg/kg b.w. (5), 10 mg/kg b.w. (10), or 20 mg/kg b.w. (20) for 15 weeks. Values are presented as means ± standard deviations (n = 8). Different letters (a > b > c > d > e) indicate significant differences with P < .05, as determined using Tukey’s honest significant difference test.
The effect of BGE on the serum and fecal lipid profiles of HFD-fed mice is shown in Table 3. The HFD control group showed a significant increase in triglyceride, total cholesterol, low-density lipoprotein cholesterol, and high-density lipoprotein cholesterol levels compared with the NC group. Conversely, the lipid profile levels decreased significantly in HFD-fed mice supplemented with BGE. Moreover, administration of BGE to the HFD group led to an increase in fecal triglyceride and total cholesterol levels of HFD-fed mice compared with those of the other HFD groups (Table 3) (P < .05), indicating that BGE supplementation effectively suppresses triglyceride and total cholesterol absorption.
Effects of BGE on Lipid Profiles and ALT and AST Levels in Mice with HFD-Induced Obesity
NC (AIN93G normal diet), 60% HFD control (Con, 60% HFD), HFD containing metformin at 250 mg/kg b.w. (Met, positive control), HFD containing BGE at 5 mg/kg b.w. (5), 10 mg/kg b.w. (10), or 20 mg/kg b.w. (20) for 15 weeks. Values are presented as means ± standard deviations (n = 8). Different letters (a > b > c > d > e) indicate significant differences with P < .05, as determined using Tukey’s Honest Significant Difference test.
HDL, high-density lipoprotein; LDL, low-density lipoprotein.
BGE administration decreased adipose tissue mass and adipocyte size in HFD-fed mice
Adipose tissue mass, especially that of abdominal adipose tissue, significantly increased in the HFD control group compared with the NC group. However, supplementation with BGE in the diet significantly reduced adipose tissue mass compared with the HFD control group. Moreover, lipid size significantly decreased in the BGE groups (Fig. 2). Therefore, BGE administration potentially decreases body weight gain by reducing both adipose tissue mass and adipocyte size.
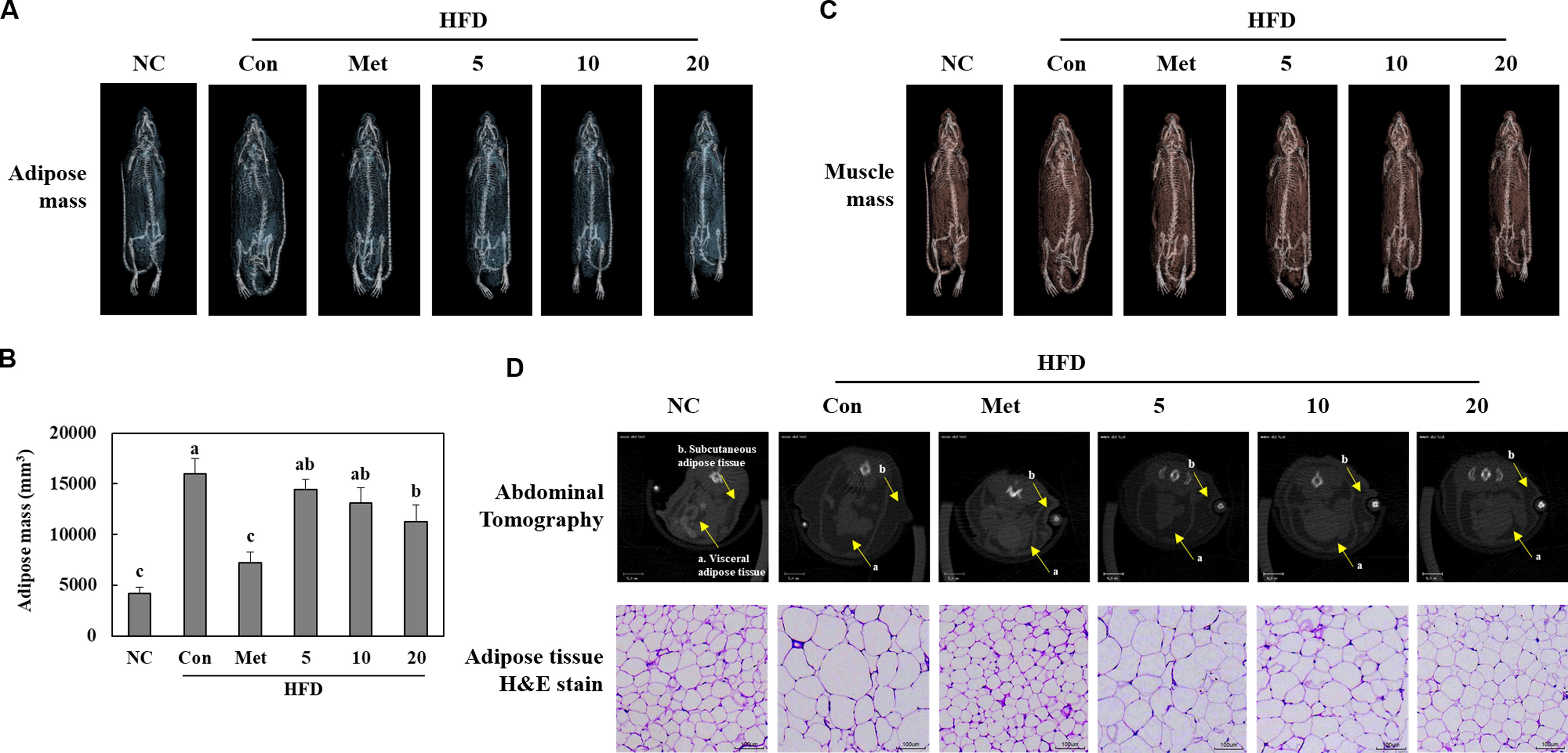
Effects of BGE on adipose tissue mass
BGE administration suppressed adipogenesis in the white adipose tissues of HFD-fed mice
The HFD control group showed higher protein levels of phosphorylated mitogen-activated protein kinase (MAPK) and adipogenic transcription factors compared with the NC group. Moreover, leptin and phosphorylated cAMP response element-binding protein (CREB) protein levels were significantly elevated in the HFD control group compared with those in the NC group. However, administration of BGE led to a reduction in the expression of these adipogenic factors in HFD mice. Interestingly, it increased adiponectin protein expression in the white adipose tissues of HFD-fed mice (P < .05) (Fig. 3).
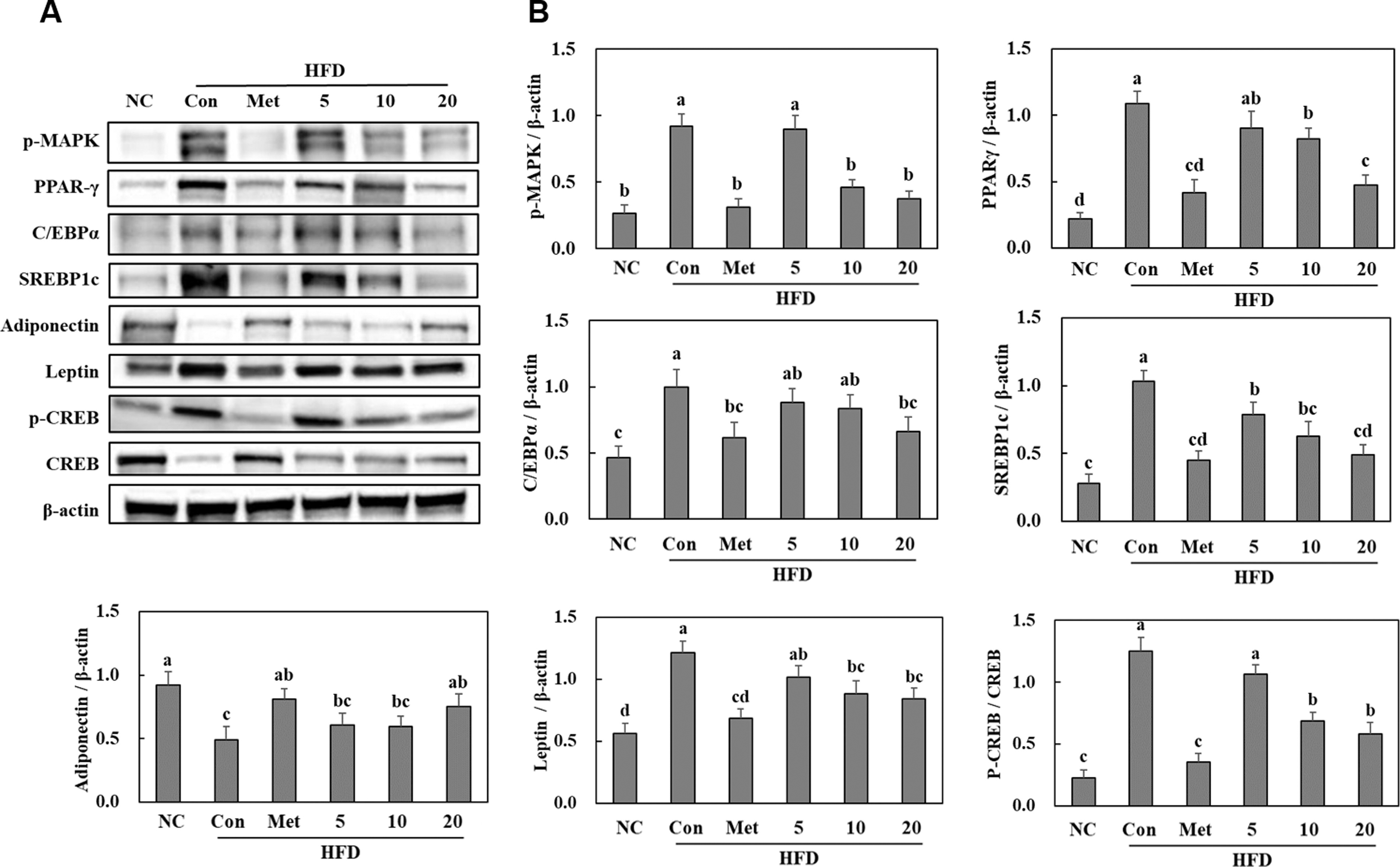
Effects of BGE on the adipogenic pathways (
BGE administration suppressed lipogenesis in the white adipose tissues of HFD-fed mice
The protein expression of glucose-6-phosphate dehydrogenase, which is required for fatty acid biosynthesis via nicotinamide adenine dinucleotide phosphate production, significantly increased in the HFD control group compared with that in the NC group. HFD feeding significantly increased the protein expression levels of fatty acid synthase (FAS), dephosphorylated ATP-citrate lyase (ACL), dephosphorylated acetyl-CoA carboxylase (ACC), and lipoprotein lipase (LPL). However, supplementation with BGE significantly increased the protein expression levels of these lipogenic factors in the white adipose tissues of HFD-fed mice (Fig. 4). These results suggest that BGE administration suppresses fat accumulation by inhibiting adipogenesis and lipogenesis.
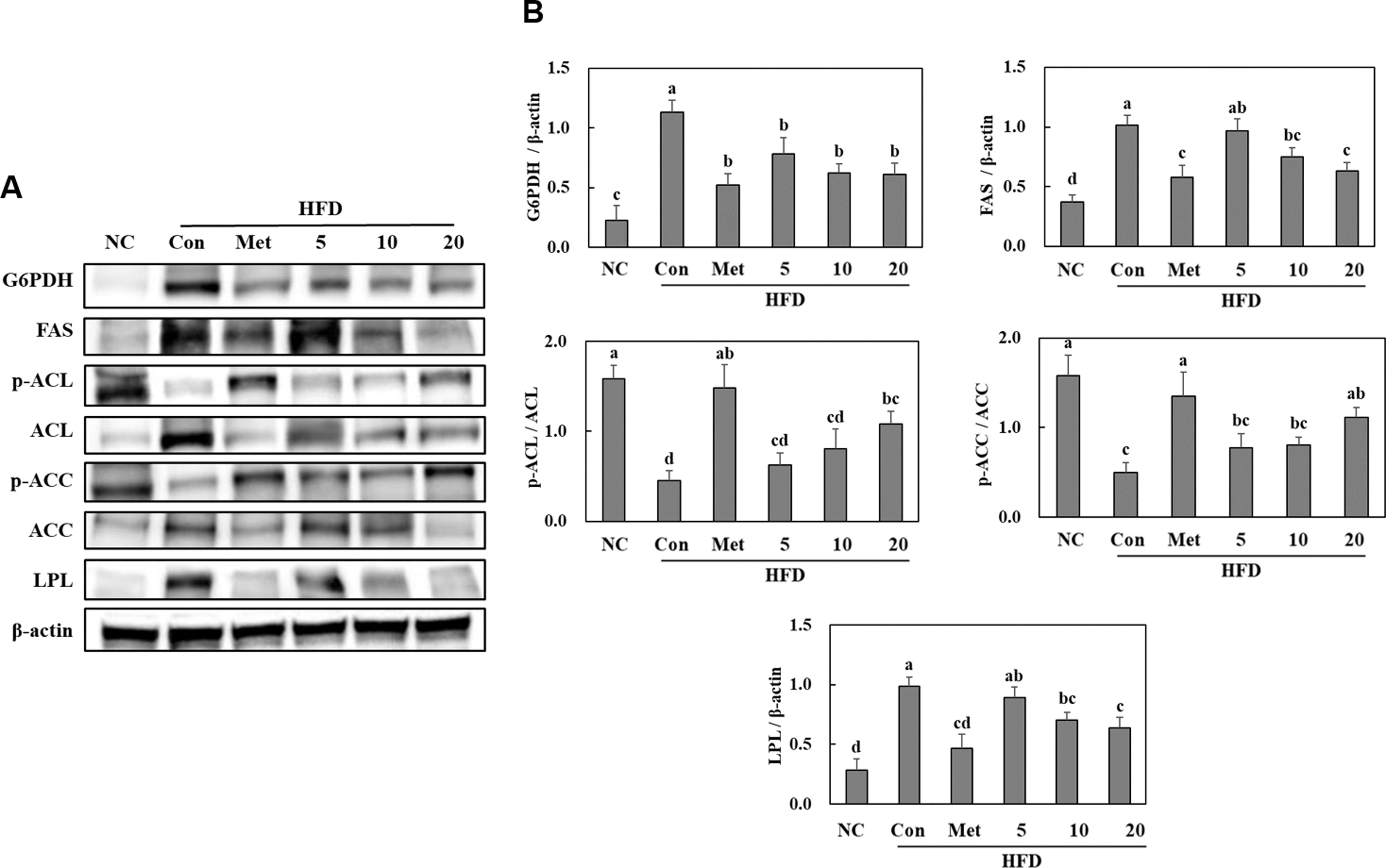
Effects of BGE on the lipogenic pathways (
BGE administration stimulated lipolysis in the white adipose tissues of HFD-fed mice
In the HFD control group, the protein expression levels of adipose triglyceride lipase (ATGL), phosphorylated hormone-sensitive lipase (HSL), and protein kinase A (PKA) decreased, while those of perilipin and phosphodiesterase 3B (PDE3B) increased compared with those in the NC group. However, administration of BGE to the HFD group led to increased protein expression levels of ATGL, phosphorylated HSL, and PKA and decreased those of perilipin and PDE3B in the white adipose tissue (P < .05) (Fig. 5).
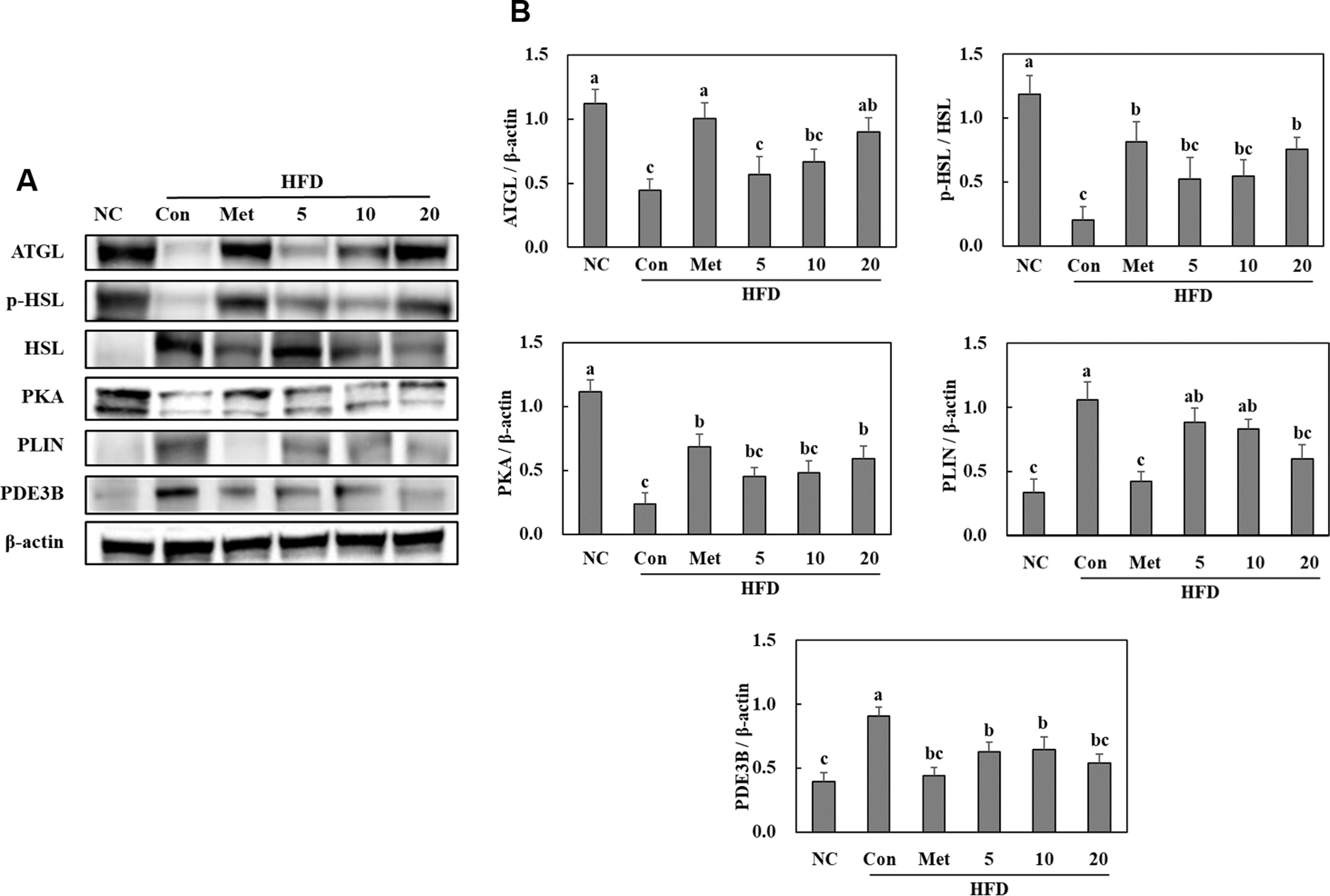
Effects of BGE on the lipolytic pathways (
BGE administration stimulated thermogenesis in the brown adipose tissues of HFD-fed mice
The thermogenesis pathway-related protein levels in the brown adipose tissues of the HFD control group were significantly decreased compared with those of the NC group. Moreover, the fatty acid-binding protein 4 (FABP4) expression was significantly increased compared with the NC group. However, BGE administration increased the thermogenesis pathway-related protein levels and decreased those of FABP4 in the brown adipose tissues of HFD-fed mice (P < .05) (Fig. 6).

Effects of BGE on the thermogenic pathways (
DISCUSSION
The biochemical mechanism underlying adipogenesis involves an exceedingly orchestrated series of molecular events. It commences with mesenchymal stem cells committing to the adipocyte lineage, triggered by various signaling pathways, including the insulin-signaling cascade. 21,22 Key transcription factors, including peroxisome proliferator-activated receptor gamma, CCAAT/enhancer-binding proteins, and sterol regulatory element-binding protein, play roles in lipid metabolism and the maturation of adipocytes. 23,24 They promote differentiation of preadipocytes into mature adipocytes and facilitate lipid storage. 25,26 As the adipocytes mature, they undergo morphological changes, accumulating lipid droplets and acquiring the ability to secrete adipokines, which play essential roles in metabolic regulation and homeostasis. In this study, HFD feeding elevated the levels of transcription factors and phosphorylated MAPK and CREB in white adipose tissues; conversely, BGE administration reduced these adipogenic factors. Upon differentiation, adipocytes accumulate fat. LPL internalizes fatty acids from triglycerides in circulating lipoproteins into adipocytes. 27 Both fatty acids from the circulating lipoproteins and those newly synthesized through lipogenesis, which is facilitated by acetyl-CoA via ACL and ACC dephosphorylation and FAS activation, are stored as triglycerides. 28 Notably, our study revealed that BGE administration reduced the expression of proteins associated with the lipogenic pathways in the white adipose tissues of HFD-fed mice. These findings suggest that BGE administration potentially diminishes HFD-induced adipose tissue weight gain by inhibiting adipogenesis- and lipogenesis-related protein activation.
Adiponectin and leptin are adipokines, bioactive substances secreted by adipose tissue, which serve crucial roles in obesity: adiponectin, with its anti-inflammatory prowess, fosters insulin sensitivity and orchestrates lipid metabolism, whereas leptin, the “master” of appetite regulation, is involved in energy balance and fat storage modulation. 29 –32 Dysfunctions of these adipokines lead to complex metabolic imbalances and the onset of obesity-related complications. This study confirmed that the HFD-induced imbalance in the protein expression levels of adiponectin and leptin was improved by BGE administration, indicating a reduction in this imbalance by decreasing adipocyte size via BGE administration.
Lipolysis is a biochemical process involving triglyceride decomposition into glycerol and free fatty acids and is primarily facilitated by enzymes such as ATGL and HSL. 33,34 In terms of its antiobesity effects, increased lipolysis contributes to adipose tissue reduction by releasing stored energy in the form of fatty acids. In contrast, thermogenesis is the process of heat production via AMPK stimulation. It is closely associated with brown adipose tissue, which contains a high density of mitochondria and is involved in dissipating energy as heat. 35 Uncoupling protein 1 in brown adipose tissue plays a critical role in uncoupling oxidative phosphorylation, leading to heat generation rather than adenosine triphosphate production. 36 Enhanced thermogenesis potentially contributes to weight loss by utilizing excess energy for heat production. The interplay between lipolysis and thermogenesis in the context of antiobesity effects involves the lipolytic mobilization of fatty acids, which subsequently serve as substrates for thermogenic processes. The efficient utilization of stored fat for energy expenditure, especially via thermogenic mechanisms, potentially contributes to weight loss and improved metabolic health. 37,38 This study demonstrated that BGE administration stimulates lipolysis- and thermogenesis-related factors in the adipose tissues of HFD-fed mice. Therefore, these finding suggest that BGE administration is effective in reducing fat accumulation and preventing obesity by increasing energy expenditure.
CONCLUSION
This study’s findings suggest potential antiobesity effects of BGE, offering a perspective on the molecular landscape of lipid metabolism regulation in adipose tissue. Furthermore, this research paves the way for further investigations and emphasizes the potential of BGE use as a multifaceted approach toward addressing the complex challenges posed by obesity.
Footnotes
AUTHORS’ CONTRIBUTIONS
Conceptualization: S.P.K., I.J., and O.-K.K. Methodology: S.P.K., I.J., and O.-K.K. Data curation: S.P.K., I.J., and O.-K.K. Investigation: S.P.K., I.J., N.K., M.K., and O.-K.K. Writing—original draft: S.P.K., I.J., and O.-K.K. All authors have read and agreed to the published version of the article.
AUTHOR DISCLOSURE STATEMENT
The authors have no conflicts of interest to declare.
FUNDING INFORMATION
No funding was received for this article.