
Abstract
Probiotics are well-known to be directly or indirectly involved in the host immune system. In this study, we analyzed the immune-boosting effects of lactic acid bacteria, including Limosilactobacillus and Lactococcus, in immunocompetent C57BL/6J mice. Three different lactic acid bacteria strains were orally administered to C57BL/6J mice for 8 weeks. Then, liver, spleen, and whole blood were harvested after sacrificing the animals. There were no significant changes in whole-body weight, weight of organs, or complete blood cell count by oral administration of lactic acid bacteria. The frequencies of CD3+, CD4+, and CD8+ T cells were significantly increased in the Limosilactobacillus reuteri MG5462 group compared to control. The frequency of NK1.1+ cells was significantly increased in the Lactococcus lactis MG5474 group compared to control. On the other hand, splenocyte proliferations and natural killer cytotoxicity did not differ between groups. In addition, the MG5462 group had a significant increase in the production of TNF-α compared to the control, which is consistent with the upregulation of T cells in the MG5462 group. Therefore, Limosilactobacillus reuteri could be a functional food additive to boost immunity by positively affecting T cell populations.
INTRODUCTION
Microbiome-based therapeutics include several approaches, including prebiotics, probiotics, synbiotics, postbiotics, and fecal microbiota transplantation. 1 Probiotics are live microorganisms that are beneficial to the health of the host when consumed in adequate amounts. 2
Limosilactobacillus, Bifidobacterium, and Lactococcus are considered inherently safe probiotics because they have been used as ingredients in foods or dietary supplements for long periods of time and in large populations without specific problems. 3 The beneficial effects of probiotics, such as production of antimicrobial compounds, competitive exclusion of pathogens, and improvement of intestinal barrier function, are important properties that contribute to the strengthening of the host immune system. 4 However, one limitation is that most probiotics cannot colonize the gut, so their beneficial effects are temporary and require ongoing supplementation. 1
The immune system defends the host against pathogens through innate and adaptive immunity. Innate immunity, 5 the initial defense, comprises myeloid cells like macrophages, monocytes, neutrophils, dendritic cells, and lymphoid cells such as natural killer (NK) cells. 6 However, the innate system’s limited capacity to handle diverse pathogens is supplemented by the adaptive immune system. T lymphocytes and B lymphocytes play roles in adaptive immunity, with T cells handling cellular immune responses and B cells producing antibodies. 7 Recent studies have reported the effects of probiotics on immune-enhancing or immunomodulatory functions. Chen et al. investigated the effect of Lactobacillus fermentum ZS40 on dextran sulfate sodium-induced ulcerative colitis mice. In that study, serum levels of pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α) were downregulated, and inflammatory cell infiltration was remarkably improved in the Limosilactobacillus fermentum ZS40-administered group. 8 In addition, another in vitro study demonstrated the immunostimulatory activity of Lactococcus lactis LM1185 on RAW 264.7 cells. The co-culture of RAW 264.7 cells and Lactococcus lactis LM1185 activated the nuclear factor-kappa B (NF-κB)/inducible nitric oxide synthase (iNOS)/prostaglandin endoperoxide synthase 2 (COX-2) signaling pathway. 9
Previously, we identified the immune-enhancing effects of Limosilactobacillus fermentum MG4538, MG5091, MG5159, and their mixture in cyclophosphamide-induced immunosuppressed mouse model. 10 We also confirmed that Limosilactobacillus fermentum MG4263, Limosilactobacillus reuteri MG5462 and Lactococcus lactis MG5474 stimulated phagocytosis of macrophage cells and enhanced the expression of NO, TNF-α, IL-6, iNOS, and COX-2 by activating the NF-κB and MAPK signaling pathways in vitro. 11
Therefore, the aim of this study was to investigate the immune-boosting effects of Limosilactobacillus fermentum MG4263, Limosilactobacillus reuteri MG5462, Lactococcus lactis MG5474 in the immunocompetent C57BL/6J mouse model to confirm their potential as useful new probiotic candidate for enhancing immune activity.
MATERIALS AND METHODS
Bacterial sample preparation
The following 3 bacterial strains were supplied by Mediogen Co., Ltd. (Jecheon, Korea): Limosilactobacillus fermentum MG4263, Limosilactobacillus reuteri MG5462, Lactococcus lactis MG5474. Each strain was activated by culturing in MRS broth (Sigma Aldrich, USA). After harvesting the bacterial pellets, these were mixed with the cryoprotectant mixture, lyophilized, and then stored at 4°C for further use.
Animal experimental procedures
Animal experiments were conducted following the guidelines outlined in the National Research Council’s Guide for the Care and Use of Laboratory Animals. Six-week-old male C57BL/6J mice were sourced from RaonBio Co. Ltd. (Seoul, Korea) and housed in groups of four per cage under controlled conditions: temperature (23 ± 1.0°C), humidity (45 ± 5%), and a 12-h light/dark cycle. Institutional Animal Care and Use Committee approval was obtained from Duksung Women’s University (No. 2023-001-013). The mice were randomly divided into four groups (n = 8 each): (1) Control (CON) group received drinking water; (2) MG4263 group received Limosilactobacillus fermentum MG4263 (1 × 109 CFU/mouse); (3) MG5462 group received Limosilactobacillus reuteri MG5462 (1 × 109 CFU/mouse); (4) MG5474 group received Lactococcus lactis MG5474 (1 × 109 CFU/mouse). Each bacterial strain was dissolved in distilled water and orally administered once daily at 1 × 109 CFU/mouse for 56 days. The CON group received distilled water instead of bacterial strains. Mice were sacrificed after 56 days, and whole blood, liver, and spleen samples were collected. Body weights were measured every 3 days throughout the experiment.
Complete blood cell count (CBC)
Blood samples were collected via cardiac puncture and immediately placed into EDTA-coated tubes (Greiner bio-one). A hematology analyzer (XN-100, Sysmex, Japan) was used to measure white blood cell count, differential white cell count, red blood cell count, hemoglobin concentration, platelet count, mean corpuscular volume, mean corpuscular hemoglobin concentration, and mean corpuscular hemoglobin.
Flow cytometry
The impact of lactic acid bacterial strains on the frequencies of CD3+, CD4+, CD8+ T cells, NK1.1+ cells, and B cells was assessed in splenocytes isolated from mice. Splenocytes were plated at a density of 1 × 10^6 cells per well in a 96-well plate and labeled with anti-mouse CD3-PE, anti-mouse CD4-PE, anti-mouse CD8-APC, anti-mouse NK1.1-APC, and anti-mouse B220-PE (BD Biosciences) at 4°C for 30 min. Following staining, flow cytometry (Novocyte Flow Cytometer) was employed to analyze the cells.
Splenocyte proliferation assay
The effects of lactic acid bacterial strains on splenocyte proliferation were assessed using a CCK-8 kit (Dojindo Laboratories). The primary splenocytes were seeded at a density of 1 × 105 cells/well in a 96-well plate and incubated for 72 h with or without 10 μg/mL of Concanavalin A (Con A). CCK-8 solution was added to each well and incubated for 3 h. Absorbance was measured at 450 nm using a microplate reader (BMG Labtech, Ortenberg, Germany).
NK cytotoxicity assay
Following dissection, primary splenocytes were promptly harvested, and the cytotoxic activity of NK cells against YAC-1 cells (KCLB) was evaluated using a CytoTox96 Non-radioactive Cytotoxicity Assay Kit (Promega). YAC-1 cells were cultured in RPMI 1640 supplemented with 10% inactivated fetal bovine serum (FBS, Gibco), and 1% penicillin-streptomycin (Gibco). They were seeded at a density of 1 × 105 cells per well in a 96-well plate and co-cultured with splenocytes (effector cells) at an E/T ratio of 5:1 for 4 h. After centrifugation of the 96-well plate at 1,600 rpm for 4 min, supernatants were transferred to a fresh plate. CytoTox96 reagent was then added to each well and incubated for 30 min at room temperature in the dark. Finally, a stop solution was added, and absorbance was measured at 490 nm using a microplate reader (BMG Labtech, Ortenberg, Germany).
Cytokine and antibody production
Lactic acid bacterial strains’ impact on cytokine and antibody production by splenocytes was assessed using enzyme-linked immunosorbent assay (ELISA). Splenocytes were plated at a density of 1 × 106 cells per well in a 6-well plate and incubated for 72 h. Culture supernatants were then collected to quantify cytokine (TNF-α and IFN-γ) and antibody (IgM and total IgG) production using a mouse ELISA kit (BD Biosciences, Invitrogen). Absorbance readings were obtained at 450 nm using a microplate reader.
Statistical analysis
Data are expressed as means ± standard deviations. Statistical analyses were conducted using SPSS version 20 (IBM Corporation). One-way analysis of variance (ANOVA) was employed to compare mean values among groups. In cases where significance was noted at P < 0.05, between-group differences were assessed using the Tukey-HSD test.
RESULTS
Effects of lactic acid bacterial (LAB) strains on the weight changes of body and organs in immunocompetent C57BL/6J mice
Throughout the experiment, the impact of LAB strain oral administration on body weight was continuously monitored. Figure 1A illustrates a consistent upward trend in whole-body weight across all groups, suggesting no adverse effects from the animal diet. Upon sacrifice, liver and spleen weights were measured, revealing no significant differences between the groups (Fig. 1B and C).
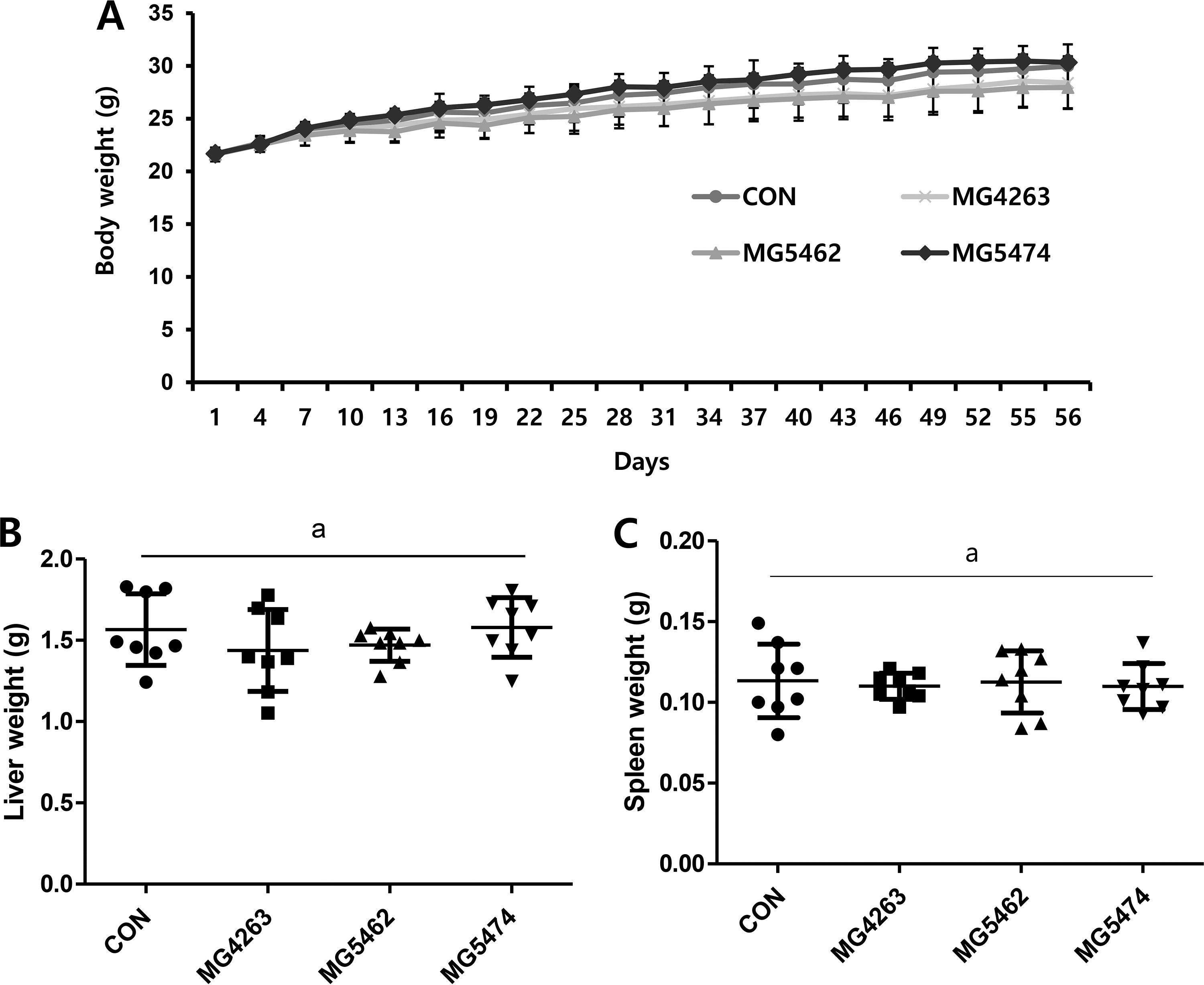
Effects of LAB strains on the weight changes of body and organs in immunocompetent C57BL/6J mice. Mice were randomly divided into four groups (n = 8): CON—drinking water; MG4263 group—Limosilactobacillus fermentum MG4263; MG5462 group—Limosilactobacillus reuteri MG5462; MG5474 group—Lactococcus lactis MG5474, and mice were orally administered once a day at 1 × 109 CFU/mouse for 56 days. The body weights were measured every 3 days. Mice were sacrificed after 56 days of experiment and the whole blood, liver and spleen were collected.
Effect of LAB strains on CBC in immunocompetent C57BL/6J mice
Table 1 displays hematological markers in immunocompetent C57BL/6J mice administered with LAB strains. While there was a slight elevation in platelet count observed in the Limosilactobacillus reuteri MG5462 and Lactococcus lactis MG5474 groups compared to others, these differences were not statistically significant. Moreover, no significant variances were noted between the groups in other parameters (Table 1).
Hematological Indices Following Lab Oral Administration In Immunocompetent C57bl/6j Mice
BASO, basophil; EOS, eosinophil; HCT, hematocrit; HGB, hemoglobin concentration; LYM, lymphocyte; MCH, mean corpuscular hemoglobin; MCHC, mean corpuscular hemoglobin concentration; MCV, mean corpuscular volume; MONO, monocyte; MPV, mean platelet volume; NEU, neutrophil; PLT, platelet count; RBC, red blood cell; RDW, red blood cell distribution; WBC, white blood cell. a letter indicates no significant difference between groups.
Effects of LAB strains on the frequency of immune cells in immunocompetent C57BL/6J mice
The Limosilactobacillus reuteri MG5462 group exhibited a significant increase in the percentage of CD3+ T cells (35.89 ± 1.3) compared to the CON group (30.94 ± 1.9) (Fig. 2A). This elevation was also evident in CD4+ and CD8+ cells from the Limosilactobacillus reuteri MG5462 group (20.86 ± 2.4, 12.20 ± 1.5, respectively) compared to the CON group (16.63 ± 0.8, 10.51 ± 0.3%, respectively) (Fig. 2B and C). Regarding NK1.1+ cells, there was a significant increase in percentage observed in the Lactococcus lactis MG5474 group (2.92 ± 0.6%) compared to the CON group (Fig. 2D). Additionally, the frequency of B220+ cells was notably reduced in all treatment groups (MG4263 58.68 ± 3.2%, MG5462 56.39 ± 5.1%, MG5474 57.89 ± 4.5%) compared to the CON group (64.00 ± 3.4%) (Fig. 2E), suggesting a compensatory mechanism wherein the increased frequency of T cells correlated with a decrease in B cells.

Effects of LAB strains on the frequency of CD3+, CD4+, CD8+ T cells, NK1.1+ cells and B220+ cells in C57BL/6J mice. Splenocytes were harvested directly after dissection, then frequencies of
Effects of LAB strains on the splenocyte proliferation and the cytotoxicity of NK cells in immunocompetent C57BL/6J mice
Splenocytes were harvested immediately after dissection and were incubated with or without Con A (10 μg/mL) for 72 h. The proliferation of splenocytes was not different in the treatment groups (without ConA; MG4263 99.4 ± 5.1% MG5462 97.3 ± 3.8%, MG5474 97.4 ± 3.2%, with Con A; MG4263, 84.9 ± 25.6%, MG5462, 70.6 ± 18.9% MG 5474 76.9 ± 19.5%, respectively) compared to CON group (100 ± 7.5%, 100 ± 40.9%) (Fig. 3A and B). The cytotoxicity of NK cells against YAC-1 cells appeared to be reduced in the Limosilactobacillus reuteri MG5462 group (2.79 ± 0.5%), but was not statistically significant compared to all other group (3.35 ± 1.4%) (Fig. 3C).
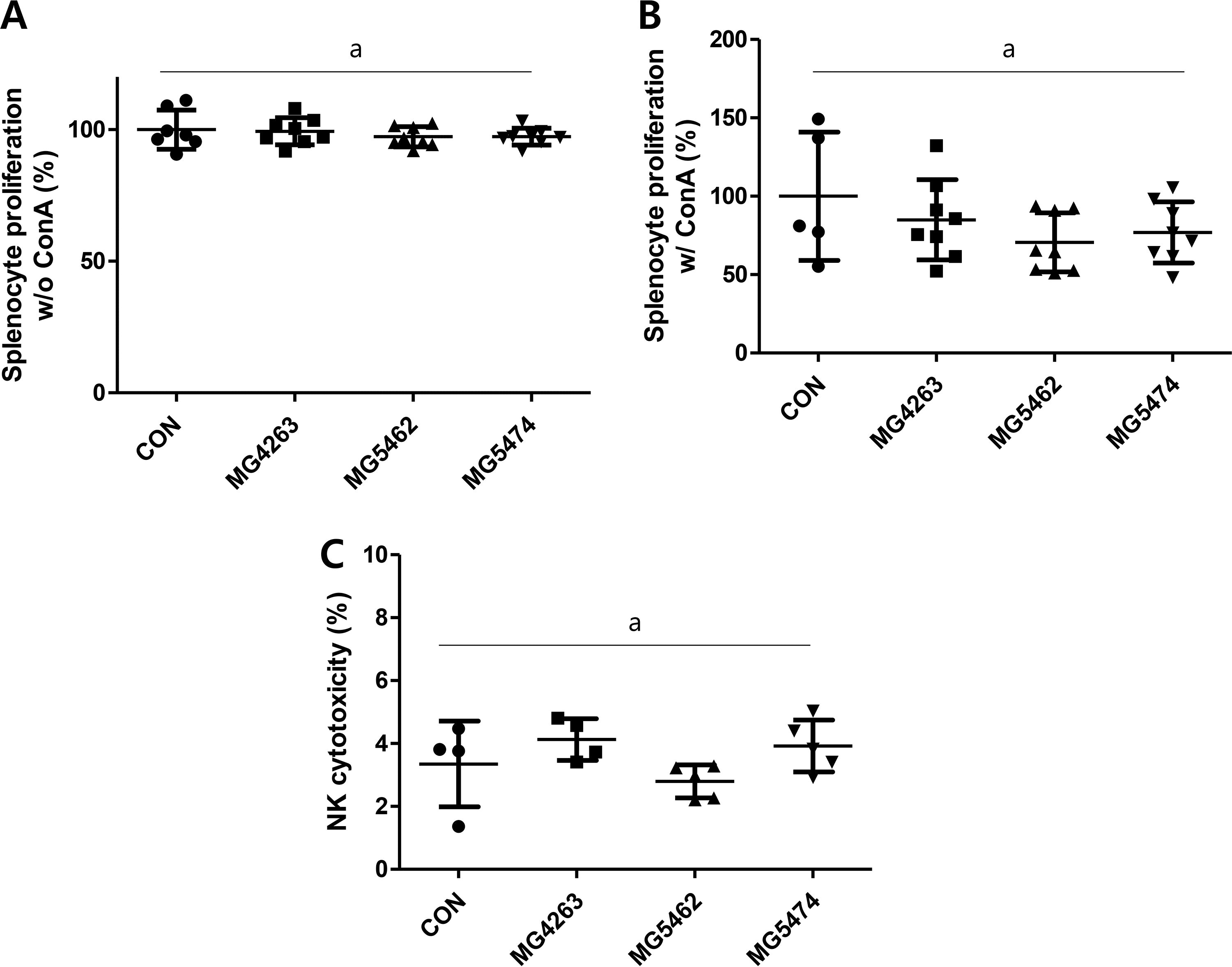
Effects of LAB strains on the splenocyte proliferation and cytotoxicity of NK cells in C57BL/6J mice. Splenocytes were harvested directly after dissection and were incubated for 72 h with or without Con A (10 μg/mL) then
Effects of LAB strains on the production of cytokines (TNF-α, IFN-γ) and antibodies (IgM, total IgG) in immunocompetent C57BL/6J mice
The effect of LAB strains on cytokine and antibody productions in primary splenocytes from immunocompetent mice was investigated using cell culture supernatants. There was a significant increase in the production of TNF-α (80.3 ± 55.9 pg/mL) from the Limosilactobacillus reuteri MG5462 group compared to CON group groups (20.5 ± 9.7 pg/mL) whereas the production of IFN-γ was not statistically different between (Fig. 4A and B). Among antibodies, IgM production was significantly reduced in the Lactococcus lactis MG5474 group (14500 ± 6200 pg/mL) compared to the MG5462 group (32100 ± 7800 pg/mL), but there was little difference in total IgG production between groups (Fig. 4C and D).
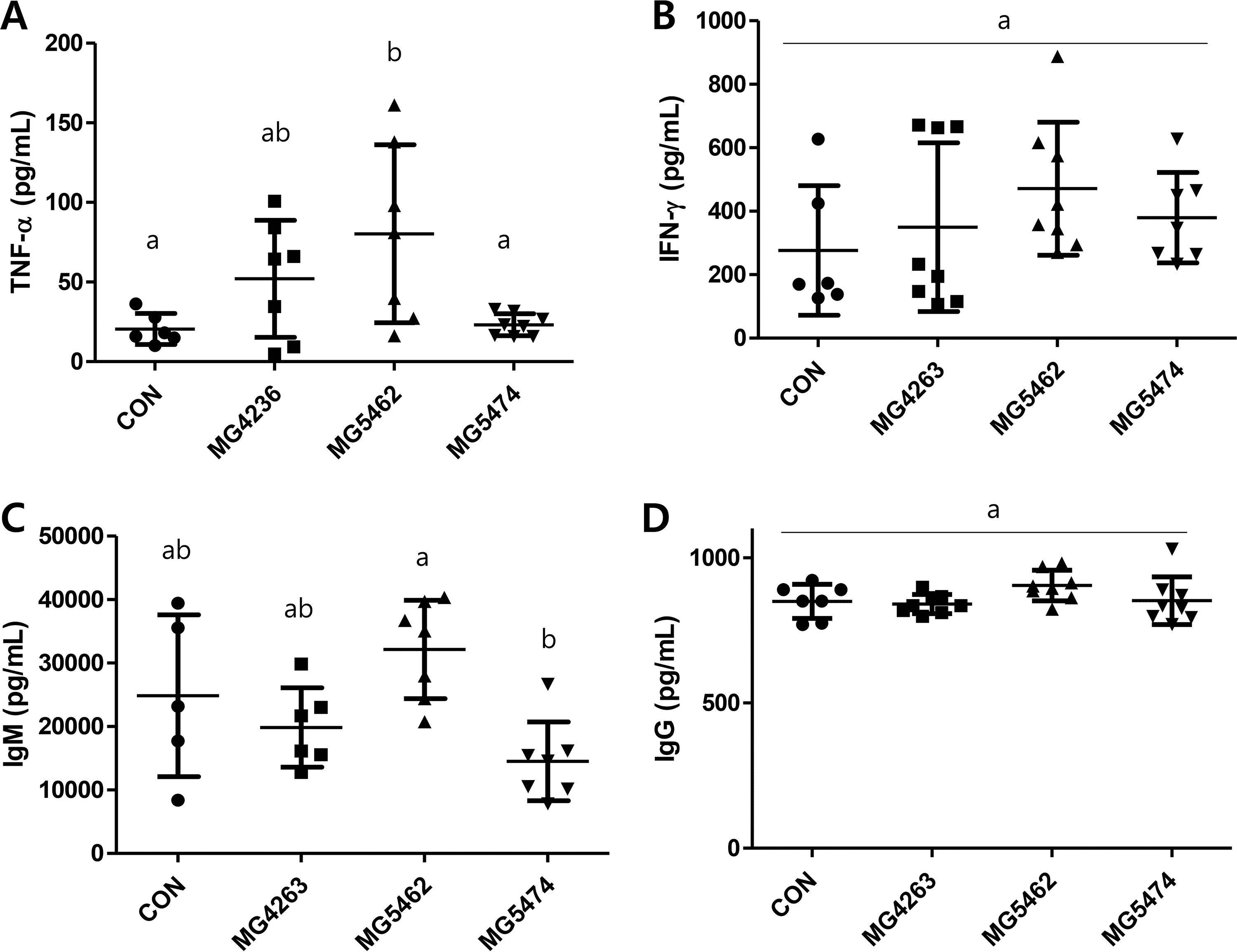
Effects of LAB strains on splenocyte cytokines production (TNF-α, IFN-γ) and antibodies (IgM, total IgG) in C57BL/6J mice. Splenocytes were harvested directly after dissection, then incubated for 72 h. ELISA kits were used to quantify the production of
DISCUSSION
Several recent studies have demonstrated immunomodulatory effects through the interaction of probiotics with immune cells, but most of these have focused on the metabolic or nutritional properties of probiotics. 12 Previously, we investigated the immune-recovering effects of Limosilactobacillus fermentum strains in vitro and in vivo. We found that seven strains of Bifidobacterium or Limocilactobacillus stimulate the production of pro-inflammatory cytokines in RAW 264.7 cells, and that oral administration of selected Limosilactobacillus fermentum strains restores the proliferative capacity of spleen cells and the frequency of lymphocyte populations in cyclophosphamide-induced immunosuppressed BALB/c mice. 10
The use of immunodeficient animal models to test immunostimulatory activity allows experiments to be performed under conditions that deviate from the normal operation of the immune system, which can provide important information for the development of therapies for patients with true immunodeficiency. However, in healthy individuals with normal immune systems, immunocompetent animal models can provide more reliable and diverse information. 13 In this study, we aimed to examine the immune-boosting effects of three lactic acid bacterial strains including Limosilactobacillus fermentum MG4263, Limosilactobacillus reuteri MG5462, and Lactococcus lactis MG5474, using immunocompetent C57BL/6J mice.
We previously reported no general toxicity by oral administration of Limosilactobacillus fermentum which agrees with data shown in Figure 1A. Also, a steady weight gain in all treatment groups indicates that Limosilactobacillus reuteri and Lactococcus lactis are safe to take orally. 10 In our previous study, reduced spleen weights by cyclophosphamide-induced immunosuppression were reversed by Limosilactobacillus fermentum treatment. 10 Interestingly, we observed little difference in a spleen weight between CON group and treatment groups in this study, which suggests the oral administration of LAB strains may provide a homeostatic immunomodulatory activity in immunocompetent mouse model (Fig. 1B and C). In addition, the treatment of LAB strains has no effect on hematological markers, including white blood cell, red blood cell, hemoglobin concentration, hematocrit, etc., in immunocompetent C57BL/6J mice (Table 1). However, a slight increase in platelet count was observed in MG5462 and MG5474 groups compared to others, which partly agrees with the significantly high values in all Limosilactobacillus fermentum treatment groups from our previous study. 10
T cells primarily play crucial roles in adaptive immune responses and are categorized into CD4+ and CD8+ T cells. 14 CD4+ T cells can differentiate into various subsets in response to cytokine signals within the microenvironment. They produce cytokines, regulate immune responses by enhancing antibody production, modulating innate immunity, and promoting immunological memory. 15 On the other hand, CD8+ T cells, also known as cytotoxic T lymphocytes, directly eliminate target cells, including virus-infected cells or tumor cells. 16 Our results show that Limosilactobacillus reuteri MG5462 increased the proportion of both CD4+ T cell and CD8+ T cell subsets, which directly contributes to the proportion of CD3+ T cells (Fig. 2). In fact, oral administration of Limosilactobacillus fermentum had little effect on the frequency of CD4+ T cells and CD8+ T cells in our previous immunosuppressed mouse model, which suggests that LAB strains could provide immune-enhancing effects in immunocompetent conditions, but not under immunodeficiency.10 On the other hand, we found that Lactococcus lactis MG5474 significantly increased the percentage of NK1.1+ cells, (Fig. 2D), which suggests that Lactococcus lactis could play an important role in innate immune responses against viral infection and cancer.
Further, we investigated whether the treatment of LAB strains could increase the proliferative capacity of primary splenocytes. The harvested primary splenocytes were cultured with or without Concanavalin A (Con A), which is an antigen-independent mitogen and can be used as an alternative T cell stimulus. 17 Unfortunately, we were not able to observe significant proliferation of splenocytes in any treatment groups compared to the CON group (Fig. 3A and B), which does not agree with our previous study done in an immunosuppressed mouse model. Together with data shown in Figure 1, there might be a homeostatic immunomodulatory mechanism existing under immunocompetent condition.
TNF-α and IFN-γ are cytokines that play roles in both innate and adaptive immune responses. 18 Our findings in FIG4 A indicate that supplementing with Limosilactobacillus reuteri led to a significant enhancement in TNF-α production in immunocompetent mice. IFN-γ also seems to be upregulated by the treatment of Limosilactobacillus reuteri, but it was not statistically significant (Fig. 4B). We previously reported that Limosilactobacillus reuteri induced the production of proinflammatory cytokines such as TNF-α, IL-1β, and IL-6 in RAW264.7 cells in vitro, which partly agrees with our data shown in Figure 4A. 10
Human antibodies offer extensive protection against pathogens and infectious diseases by binding to the pathogen, thereby activating both the complement system and immune cells. 19 Primarily involved in humoral antibody responses, Immunoglobulin G (IgG) plays a pivotal role in mediating effector functions, regardless of whether the protection is against viral or cellular pathogens. We found that the production of antibodies, either IgM or IgG, was not significantly increased by the treatments with LAB strains (Fig. 4C and D).
Our study confirmed that oral administration of Limosilactobacillus reuteri MG5462 enhances immune responses by increasing T cell proliferation and upregulating the levels of immune-activating cytokines in immunocompetent mice.
AUTHORS’ CONTRIBUTIONS
Conceptualization: H.J., H.H.L., H.K., and H.C. Investigation: H J. and H.H.L., Article: H.K. and H. C.
Footnotes
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This research was funded by Industry University Institute Collaboration R&D (Development of functional food for immune enhancing using the novel probiotics based on microbiome) funded by TIPA (S3301283). This work was also supported by the Priority Research Center Program through the NRF funded by the Korean Ministry of Education, Science and Technology (2016R1A6A1A03007648).