
Abstract
Menopausal depression, often associated with hormonal fluctuations such as decreased estrogen levels, imposes significant mental health burdens. Despite the antidepressant biological properties of standardized rice bran supplement (RBS), its impact on menopausal depression and underlying mechanisms remains largely unexplored. In this study, we investigated the antidepressant effects of RBS in a mouse model of estrogen deficiency–induced depression. Ovariectomized (OVX) mice received oral doses of RBS (250 and 1000 mg/kg) and 17β estradiol over a 20-week period. RBS administration resulted in decreased immobility time in the tail suspension and forced swim tests, along with increased locomotor activity in the open field test. Furthermore, RBS enhanced nitric oxide production and neuronal nitric oxide synthase (nNOS) expression in the hippocampi of OVX mice. Additionally, RBS administration phosphorylated extracellular signal-regulated kinase (ERK), cAMP response element-binding protein (CREB), and tropomyosin receptor kinase B and increased the protein expression of brain-derived neurotrophic factor (BDNF) in the hippocampus. These findings suggest that RBS alleviated depressive behaviors in OVX mice by augmenting hippocampal nNOS expression and activating the ERK-CREB-BDNF signaling pathway. Therefore, based on these results, we propose that RBS is a promising agent to treat menopausal depression, a challenging condition.
INTRODUCTION
The onset of menopause, characterized by a decline in estrogen levels, marks a significant hormonal transition in women’s lives. This hormonal decrease often precipitates a range of physiological and psychological alterations, particularly depression. 1 Menopausal depression, triggered by estrogen deficiency, presents a considerable challenge to women’s health, impacting their quality of life. 2 Traditionally, menopausal hormone therapy (MHT) has served as an effective intervention for managing menopausal depression, employing a range of pharmaceutical agents such as estrogens, progestogens, and synthetic steroids. 3,4 However, the association between MHT and significant adverse effects, such as an elevated risk of breast cancer, cardiovascular disease, and stroke, underscores the need to explore alternative treatments for menopausal depression. 5
In recent years, the potential therapeutic properties of rice bran as a safer alternative to pharmaceutical interventions have been extensively researched. Rice bran, a by-product of rice milling, is rich in protein, fat, vitamins, and phytochemicals compared with bran from other cereals. 6 Numerous studies have demonstrated its diverse effects, including antidiabetic, anti-inflammatory, antioxidant, and lipid-lowering activities, including cognitive enhancement and sleep promotion. 7 –10 Our previous studies have shown that rice bran supplement (RBS) induces reduced sleep latency and enhanced non-rapid eye movement sleep in mice through inhibition of histamine H1 receptors. Shendy et al. 11 reported that rice bran extract attenuated colitis-induced depressive-like behavior in a forced swimming experiment. Furthermore, γ-oryzanol, a major constituent of rice bran, exerts antidepressant properties in models of estrogen deficiency–induced depression by modulating the hippocampal nitric oxide–neuronal nitric oxide synthase (NO-nNOS) pathway mediated by the estrogen receptor β. 12 These findings suggest the promising role of RBS in ameliorating depression symptoms. Consequently, the objective of this study was to explore the effects of RBS on mice with estrogen deficiency–induced depression and elucidate its underlying mechanisms, specifically focusing on the hippocampal NO-nNOS and the extracellular signal-regulated kinase (ERK)–cAMP response element-binding protein (CREB)–brain-derived neurotrophic factor (BDNF) pathway.
MATERIALS AND METHODS
Preparation of RBS
The standardized RBS (Lot No. 221116-010), sourced from S&D (Cheongju, South Korea), was prepared according to established procedures. 9,10 In brief, rice (Oryza sativa L.) bran underwent extraction in a 20% ethanol (10 times, v/w) for 8 h at 40°C. After concentration and spray-drying, the extract was to be powdered RBS. HPLC analysis was conducted using a LiChrospher 100 DIOL column (250 × 4.0 mm, 5 μm; Merck Milipore, Billerica, MA, USA). Solvent systems used for separation were an isocratic elution (A: 0.1% acetic acid in cyclohexane and B: 0.1% acetic acid in ethyl acetate) in a ratio of 85:15 (v/v). The injection volume was 10 μL, and ultraviolet detection was set at 315 nm; chromatography was executed at 30°C with a flow rate of 0.5 mL/min. The HPLC quantitative analysis was replicated three times. After this, RBS was standardized to contain 4.5 mg/g extract of γ-oryzanol.
Animals and experimental procedure
Female C57BL/6N mice (aged 5 weeks old) were obtained from Koatech Animal Inc. (Pyeongtak, South Korea). The mice were housed in a controlled environment with a 12-h light/dark cycle and had free access to food and water. After a 7-day acclimatization period, the mice underwent anesthesia with 2% isoflurane. After anesthesia, bilateral dorsal flank incisions were made to perform ovariectomy (OVX). In sham-operated (SHAM) mice, the ovaries were exposed but left intact. After a 2-week recovery period, a total of 50 mice (aged 8 weeks old) were allocated into five groups (n = 10 per group): (1) SHAM+VEH group, administered the vehicle (VEH); (2) OVX+VEH group, administered the VEH; (3) OVX+17β-estradiol (E2) group, administered 10 μg/kg body weight of E2; (4) OVX+RBS250, administered RBS 250 mg/kg body weight; and (5) OVX+RBS1000, administered RBS 1000 mg/kg body weight. Throughout the 20-week duration, all mice received their designated treatments via daily oral administration. The experimental timeline is shown in Figure 1. After this period, behavioral assessments were performed, following which the mice were euthanized, and their brain tissues were collected and stored at −80°C for further analysis. This study was conducted according to the Animal Research: Reporting of In Vivo Experiments guidelines 2.0. and approved by the Institutional Animal Care and Use Committee of the Korea Food Research Institute (KFRI-M-20020).
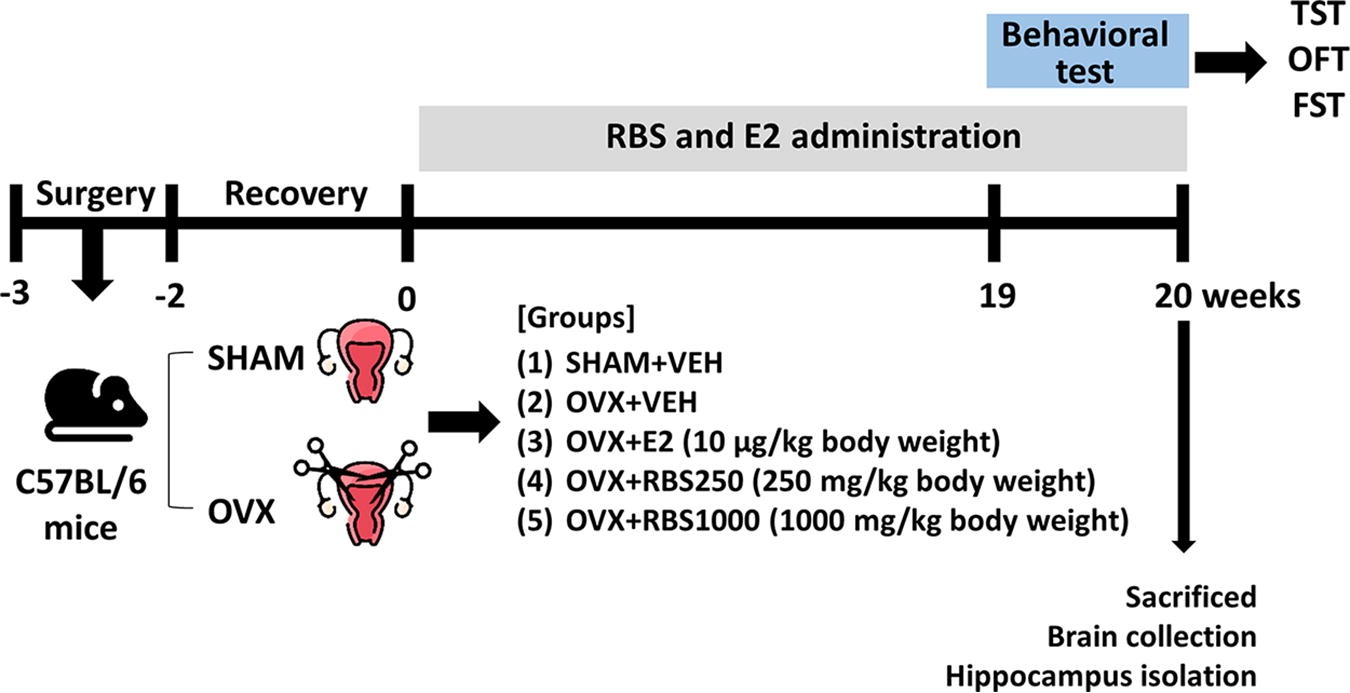
Experimental protocol: The female C57BL/6N mice underwent OVX. After a 2-week habituation period, the OVX mice were administered either the VEH, RBS (250 or 1000 mg/kg per day, p.o.) and E2 (10 μg/kg per day) over 20 weeks. Subsequently, behavioral tests were conducted. After completion of their behavioral tests, the mice were euthanized, and their brains were isolated. The hippocampi were dissected from the brains and subjected to analysis. E2, 17β-estradiol; FST, forced swimming test; OFT, open field test; OVX, ovariectomized; RBS, rice bran supplement; TST, tail suspension test; VEH, vehicle.
Forced swimming test
The forced swimming test (FST) was conducted following procedures outlined in our previous study. 13 Each mouse was individually placed in a cylinder (diameter 13 cm; height, 24 cm) filled with water (depth, 10 cm; temperature, 22 ± 1°C) and allowed to swim for 6 min. Their motionless times were recorded and analyzed using SMART 3.0 software (Panlab SL, Barcelona, Spain).
Tail suspension test
The tail suspension test (TST) was conducted in accordance with methodologies described in our previous studies. 13 Adhesive tape was attached to the tails of mice; thereafter, mice were hung on a TST apparatus, and their motionless times were recorded for a duration of 6 min using an automated device (BioSeb, Chaville, France).
Open field test
The open field test (OFT) was performed according to our previous studies. 13 Each mouse was individually placed in an acrylic box (40 × 40 × 40 cm) and allowed to freely explore the environment for 6 min. Their movement, including the total distance covered, was monitored and quantified using SMART 3.0 software (Panlab SL).
Measurement of NO levels
To indirectly analyze NO production, we employed a Griess assay reagent (Promega, Madison, WI, USA). Brain tissues underwent homogenization in Radioimmunoprecipitation Assay buffer followed by centrifugation. The supernatant obtained was collected, and equal volumes of the mixed sulfanilamide solution and supernatant were combined in microtubes. Subsequent centrifugation at 3000 g for 5 min at 4°C was performed. Afterward, the resulting supernatant was transferred to a 96-well plate for absorbance measurement at 540 nm using a microplate reader (Molecular Devices, Sunnyvale, CA, USA). Finally, nitrite concentrations were normalized to the protein content of the brain tissues.
Immunohistochemistry
The brains were initially fixed in 10% formalin overnight, followed by dehydration in a 30% sucrose solution. Subsequently, brains were incubated with optimal cutting temperature solution and frozen at –80°C. Using a cryostat (Leica Microsystems, Mannheim, Germany), the frozen brains were sectioned into 20 μm serial hippocampal sections. Afterward, the sections were rinsed thrice for 5 min each in phosphate-buffered saline with triton X-100 (PBST), followed by blocking with 10% goat serum in PBST for 1 h. Subsequently, the sections were exposed to a primary anti-nNOS antibody (1:1000) overnight at 4°C. After rinsing, the sections were treated with a secondary Alexa Flour-conjugated 488 antibody (1:200) for 2 h. After final washing, the sections were mounted on slides using a mounting solution containing DAPI, and visualization of nNOS expression was performed using a confocal microscope (Zeiss, Jena, Germany).
Western blot analysis
Hippocampal tissues were homogenized, and the protein concentrations were determined using a BCA protein assay kit (Thermo Fisher Scientific, Waltham, MA, USA). Subsequently, 20 μg of proteins were separated by SDS-PAGE and transferred onto PVDF membranes. After blocking with 5% skim milk in Tris-buffered saline with 0.1% Tween-20 (TBST) for 1 h, the membranes were incubated with primary antibodies (BDNF, CREB, p-CREB, ERK, p-ERK, tropomyosin receptor kinase B [TrkB], p-TrkB, and β-actin; Cell signaling Technology, Inc., Danvers, MA, USA) overnight at 4°C. After washing with TBST, the membranes were incubated with an HRT-conjugated secondary antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) for 1 h. Protein expression levels were analyzed using an ECL solution (Thermo Fisher Scientific) and normalized to β-actin. The relative protein intensities were quantified using ImageJ (National Institutes of Health, Bethesda, MD, USA).
Real-time quantitative polymerase chain reaction
Hippocampal RNA was extracted utilizing the Nucleospin RNA plus kit (Macherey-Nagel, Düren, Germany). Subsequently, cDNA synthesis was performed using the ReverTra Ace qPCR RT Master Mix Kit (TOYOBO, Osaka, Japan), followed by real-time quantitative polymerase chain reaction (PCR) conducted on the QuantStudio 6 real-time PCR system (Applied Biosystems, Waltham, MA, USA) with SYBR Premix Taq (TOYOBO), with a final reaction volume of 20 μL. The primer sequences used were as follows: nNOS-forward: 5′-GTGGCCATCGTGTCCTACCATAC-3′ and reverse: 5′-GTTTCGAGGCAGGTGGAAGCTA-3′: β-actin-forward: 5′-CGAGCACAGCTTCTTTGCAGC-3′ and reverse: 5′-CCTTCTGACCCATTCCCACC-3′. After normalization with β-actin gene expression, relative changes were determined by the 2−ΔΔCT comparative method.
Statistical analysis
All data were presented as means ± standard errors of the means. One-way analysis of variance was performed using Prism 10 software (GraphPad Software, Inc., San Diego, CA, USA) to analyze the statistical differences between groups using Tukey’s post hoc test for multiple comparisons. A significance level of P < .05 was considered statistically significant.
RESULTS
RBS improves depressive-like behaviors in OVX mice
In the OVX+VEH group, the immobility time in both the TST and FST significantly increased compared with the SHAM+VEH group (TST, P < .001; FST, P < .01) (Fig. 2A, B). However, RBS administration significantly decreased the immobility time in the TST compared with in the OVX+VEH group (P < .01). In the FST, both the OVX+RBS250 (P < .01) and the OVX+RBS1000 groups (P < .001), as well as the OVX+E2 group, exhibited decreased immobility time compared with in the OVX+VEH group (P < .01) (Fig. 2B). Additionally, the OVX+VEH group indicated a reduced total movement distance in the OFT compared with the SHAM+VEH group (P < .01), whereas administration of RBS and E2 significantly increased total movement distance (Fig. 2C, D). Therefore, our results demonstrated that RBS administration improved depressive-like behaviors in OVX mice, similar to E2.
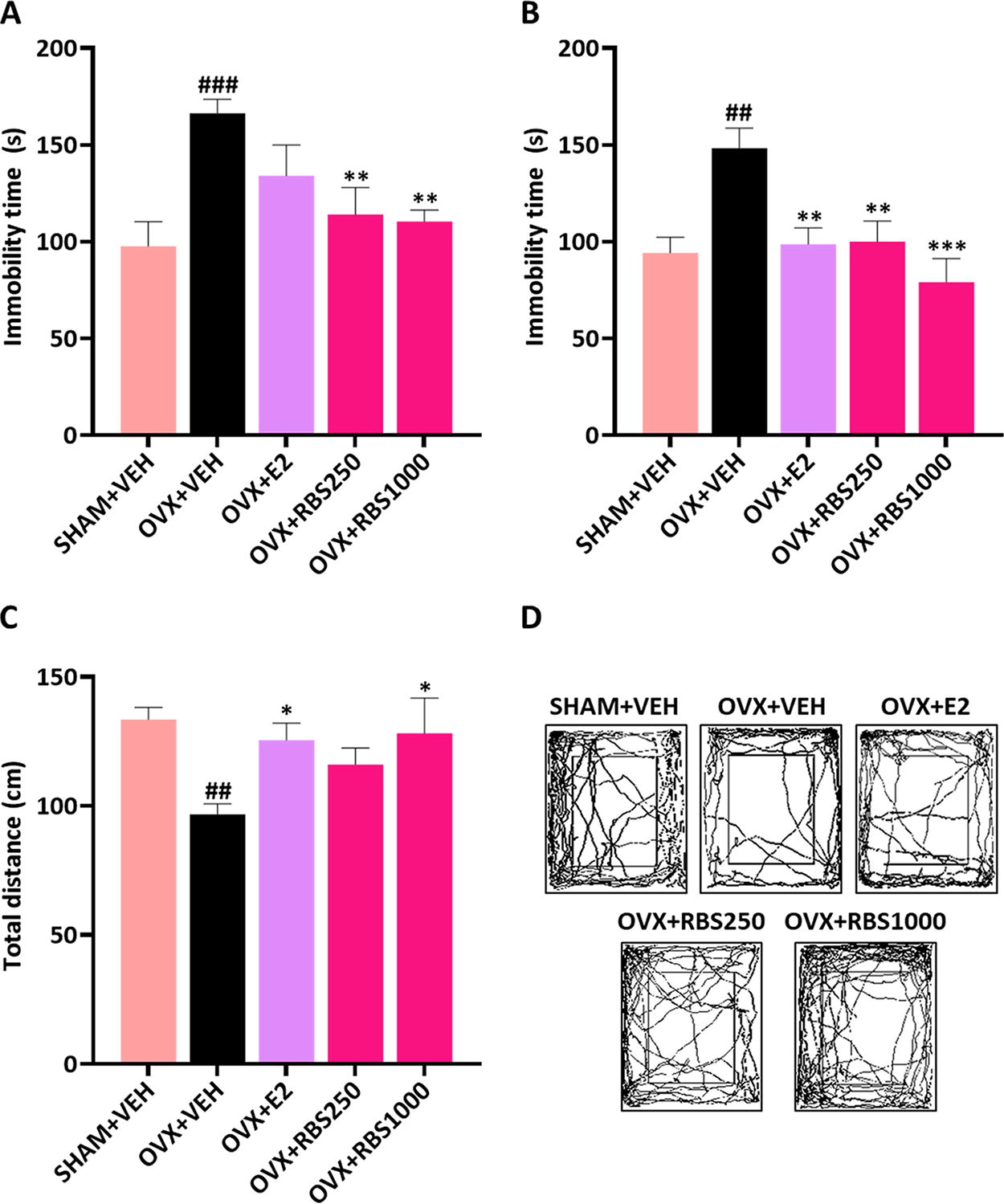
RBS attenuated depressive-like behaviors in estrogen deficiency–induced mice After administering RBS and E2 for 20 weeks, depression-related behavioral tests were conducted.
RBS increases NO production by modulating nNOS expression in the hippocampi of OVX mice
To investigate the potential link between the ameliorative impact of RBS on estrogen deficiency–induced depression and NO production, we assessed the levels of NO in the brains of OVX mice. Our results revealed a notable decrease in NO levels in the brains of the OVX+VEH group compared with the SHAM+VEH group (P < .001). However, RBS administration elicited a significant increase in NO levels (Fig. 3A).
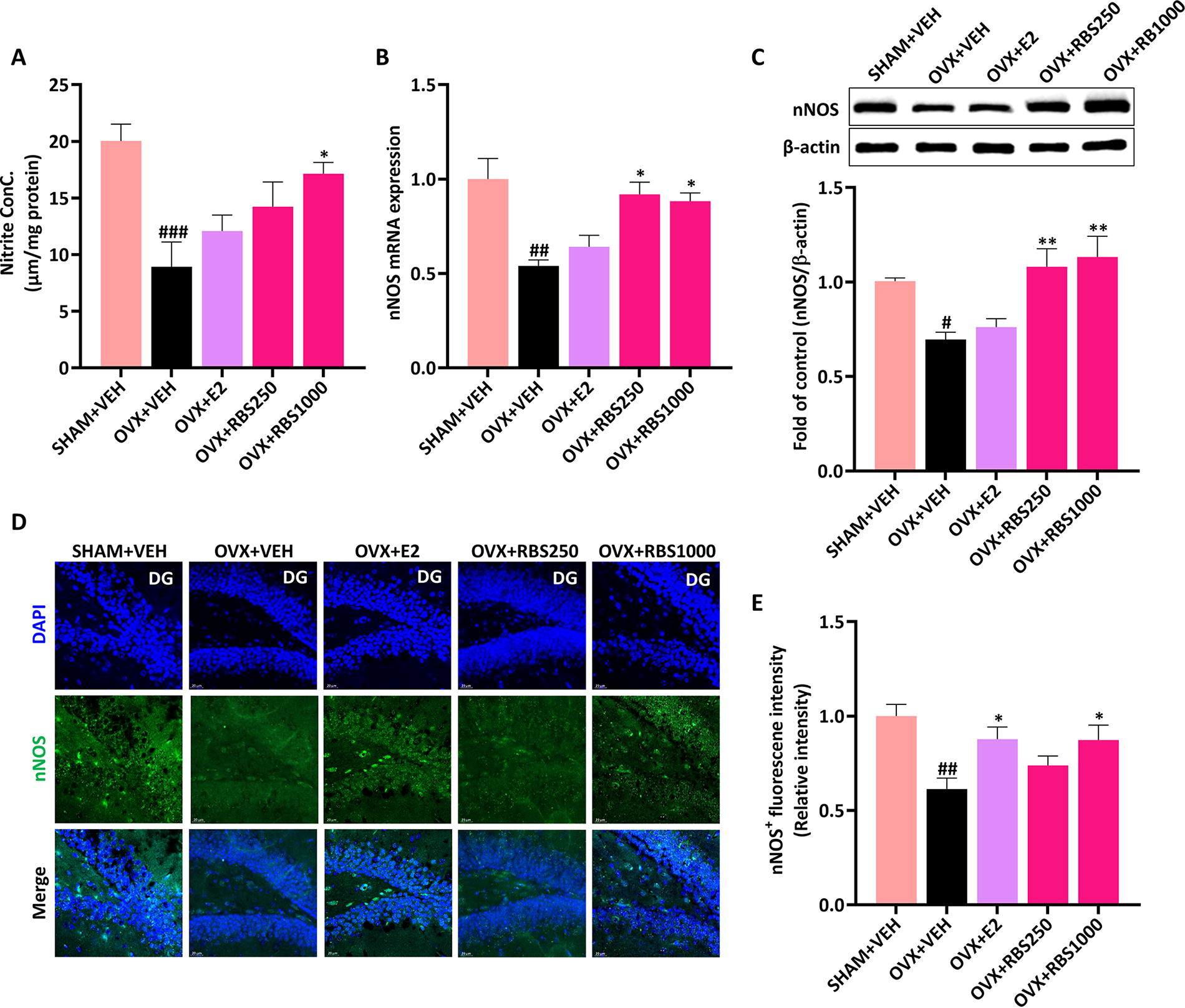
RBS increased NO production in the brains of OVX mice.
Next, we examined the mRNA and protein expression levels of nNOS in the hippocampi of OVX mice. As depicted in Figure 3B and C, nNOS mRNA and protein expressions displayed a significant reduction in the hippocampi of the OVX+VEH group. Conversely, RBS administration at doses of 250 and 1000 mg/kg body weight led to an increase in nNOS mRNA and protein expressions in the hippocampi of OVX mice. Furthermore, our immunohistochemistry analysis revealed a pronounced rise in nNOS expression within the dentate gyrus region of the hippocampus in the OVX+RBS1000 group compared with the OVX+VEH group, suggesting that RBS administration effectively reversed the decrease in estrogen deficiency-induced NO levels by enhancing the expression of hippocampal nNOS mRNA and protein (Fig. 3D, E).
RBS activates the ERK-CREB-BDNF signaling pathway in the hippocampi of OVX mice
To further elucidate the mechanisms underlying the improvement of depressive-like behavior of RBS, we investigated the ERK-CREB-BDNF signaling pathway, which is recognized as one of the key molecular pathways, in the hippocampus of depressed mice. As depicted in Figure 4A and B, the hippocampal BDNF levels and phosphorylated ERK, CREB, and TrkB exhibited significant reductions in the OVX+VEH group compared with the SHAM+VEH group (P < .001; BDNF, P < .01). However, administration of E2 significantly restored BDNF protein levels and the phosphorylation of ERK, CREB, and TrkB in the hippocampi of OVX mice (p-ERK and p-CREB; P < .001; BDNF and p-TrkB, P < .01). Similarly, treatment with RBS at both 250 mg/kg and 1,000 mg/kg doses activated the ERK-CREB-BDNF signaling pathway in the hippocampus, resembling the effects observed with E2 treatment. Overall, our findings indicate that RBS administration alleviates depressive-like behaviors through the modulation of NO-nNOS and the activation of the hippocampal ERK-CREB-BDNF signaling pathways in OVX mice.
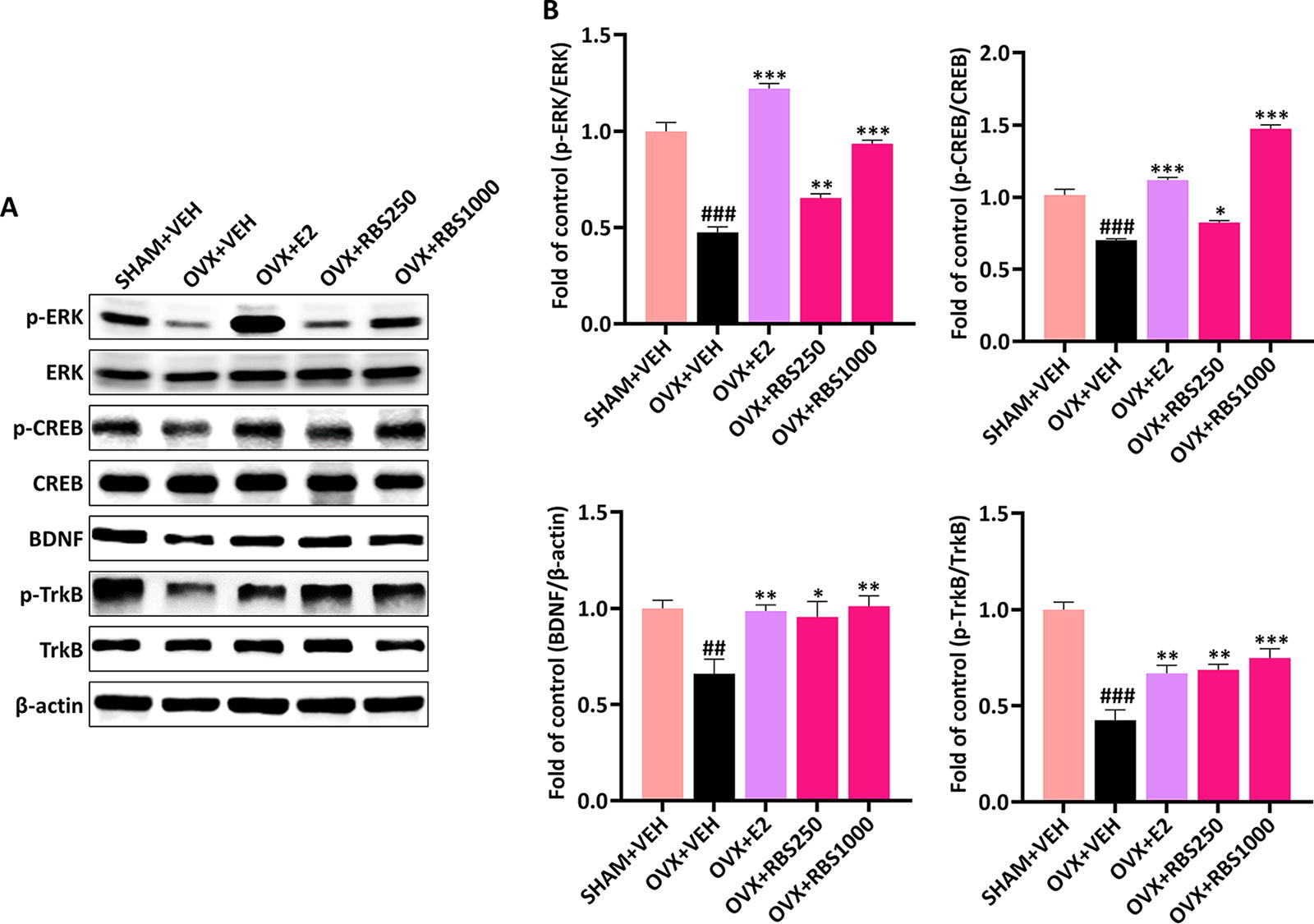
RBS activated the ERK-CREB-BDNF signaling pathway in the hippocampi of OVX mice.
DISCUSSION
Menopausal depression has been extensively studied. It is often triggered by hormonal imbalances, particularly a reduction in estrogen levels, with estrogen replacement therapy established as an effective treatment. 14,15 Therefore, we employed OVX mice as an experimental model to investigate the impact of hormonal fluctuations on mood and behavior, specifically depressive symptoms. 16 –18 Our findings are consistent with existing literature, revealing that OVX mice lacking estrogen exhibit depressive-like behaviors, while RBS administration over a 20-week period mitigated these depressive symptoms. Previous research has shown that rice bran extract has the capability to alleviate depressive-like behavior in other conditions, such as dextran sulfate sodium-induced colitis. 11 Therefore, our findings highlight the potential antidepressant efficacy of RBS in alleviating estrogen deficiency-induced depression, suggesting its promise as an alternative therapeutic option for alleviating depressive symptoms.
Our study suggests that RBS may exert antidepressant efficacy in mice with estrogen deficiency–induced depression through alterations in brain NO levels. However, the relationship between NO and depression remains a topic of debate in the literature. Within the central nervous system (CNS), nNOS serves as a crucial signaling molecule impacting depressive-like behavior, particularly under conditions of chronic stress. 19 Notably, the regulation of hippocampal NO levels in depression differs between sexes, with estrogen playing a significant role in modulating hippocampal NO and nNOS expression and influencing affective behaviors. 20 Specifically, deficiencies in hippocampal NO have been associated with depressive behaviors induced by ovariectomy in mice. Remarkably, E2 therapy has shown efficacy in elevating serum NO concentrations and mitigating depressive symptoms in menopausal women. 21 Consistent with these findings, our data revealed decreased NO concentrations and nNOS expression in mice, with RBS administration ameliorating these changes induced by estrogen deficiency. Moreover, it had no effect on serum E2 levels (Table S1 in Supplementary Data S1). These findings are consistent with prior research indicating a connection between specific estrogen functions in the CNS and enhanced NO production. 22 Collectively, our results imply that modulating of NO production could be a crucial mechanism underlying the therapeutic efficacy of RBS in in estrogen deficiency-induced depression.
CREB-dependent gene expression is widely acknowledged to play a multifaceted role in signal transduction cascades, responding to diverse stimuli including growth factors, hormones, and synaptic plasticiy. 23 Notably, reduced CREB expression has been observed in the postmortem brains of depressed patients. Additionally, antidepressant treatment has been demonstrated to elevate hippocampal CREB phosphorylation in rats. 24,25 ERK signaling, implicated in stress-induced depression, directly modulates CREB phosphorylation, essential for synaptic plasticity induction. 26,27 Additionally, BDNF, a neurotrophin pivotal in depression pathophysiology, is regulated by antidepressants via CREB-mediated transcription. 28 Antidepressant therapy has been shown to increase both hippocampal BDNF and TrkB levels in OVX rats exposed to chronic mild stress, despite their initial diminishment. 29 Our results mirrored these findings, demonstrating a notable decrease in phosphorylated ERK, CREB, and TrkB levels, accompanied by decreased BDNF expression in OVX mice compared with the SHAM+VEH group. However, administration of RBS led to an upregulation of these signaling molecules in the hippocampi of OVX mice, suggesting that the potential antidepressant effects of RBS may involve modulation of the hippocampal ERK-CREB-BDNF signaling pathway.
Rice bran contains high levels of γ-oryzanol, α-tocophoerol, and sitosterol. 30 These components have demonstrated antidepressant-like effects in diverse animal models of depression and anxiety. Previous research has shown that α-tocophoerol exhibits antidepressant-like effect in a depressive mouse model induced by tumor necrosis factor-alpha (TNF-α). β-sitosterol and its derivatives modulate monoamine levels to exert antidepressant effects. 31,32 Additionally, γ-oryzanol has been reported to induce an antidepressant-like effect in models of chronic unpredictable mild stress in Drosophila melanogaster and alcohol-induced anxiety in mice by regulating monoamines and BDNF and interleukin (IL)-1β signaling. 33,34 Our previous research has contributed to understanding the antidepressant efficacy of γ-oryzanol in animal models of estrogen deficiency–induced depression by modulating NO-nNOS and ERK-CREB-BDNF pathways. 12 Therefore, our findings, along with previous research on the antidepressant effects of the major components of rice bran, suggest that RBS may have antidepressant effects, likely mediated through its ability to generate NO and activate the ERK-CREB-BDNF signaling. However, future studies are needed to fully elucidate the underlying mechanisms and evaluate the clinical efficacy of RBS. Collectively, our results demonstrate that RBS ameliorates depressive-like behaviors in OVX mice by increasing hippocampal nNOS expression and upregulating the ERK-CREB-BDNF signaling pathway. These results imply that RBS shows potential as a therapeutic approach for alleviating menopausal depression.
Footnotes
AUTHORS’ CONTRIBUTIONS
M.Y.U. contributed to the study design. M.K. performed experiments and data analysis. M.Y.U. and M.K. contributed to article writing and editing. S.C. and C.L. reviewed the article. All authors approved the final article.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This work was supported by the Korea Institute of Planning and Evaluation for Technology in Food, Agriculture, Forestry and Fisheries through the High Value-Added Food Technology Development Program, funded by the Ministry of Agriculture, Food and Rural Affairs (
SUPPLEMENTARY MATERIAL
Supplementary Data S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.