
Abstract
Iron supplementation is a common method for alleviating symptoms of iron deficiency, but excessive iron intake may lead to systemic copper deficiencies and hypercholesterolemia. In our study, we explored the intricate relationship between dietary iron and copper levels and their impact on cholesterol metabolism. Using a rat model, we conducted dietary interventions with varying iron and copper concentrations and analyzed hepatic transcriptomes. High iron intake coupled with low copper intake induced hypercholesterolemia and altered the expression of genes associated with cholesterol and lipid metabolism, thereby, exacerbating cardiovascular disease risks. Conversely, copper supplementation mitigated these hepatic gene expression alterations, suggesting that dietary copper plays a role in cholesterol regulation. Transcriptomic analysis revealed significant upregulation of genes involved in cholesterol synthesis and antioxidative pathways in response to high iron intake, while genes involved in cholesterol elimination were downregulated. Furthermore, high iron consumption was associated with cellular apoptosis and the activation of cholesterol synthesis. Our findings underscore the importance of balanced iron and copper intake in cholesterol homeostasis and highlight the potential of copper supplementation for mitigating iron-induced hypercholesterolemia.
Iron deficiency is a physiological condition characterized by inadequate iron storage and utilization in the blood and/or tissues. 1 Several factors can lead to iron deficiency, including dietary insufficiency, blood loss, and gastrointestinal (GI) surgery. 1 Dietary insufficiency primarily occurs from insufficient consumption of iron-rich foods. 2 Blood loss-induced iron deficiency can stem from menstruation in women, 3 internal bleeding, or surgery. 4,5 GI surgery also contributes to iron deficiency, as iron is mainly absorbed in the upper GI tract. 6 Prolonged iron deficiency triggers multiple pathological disturbances. Fatigue and weakness are commonly observed in individuals with iron deficiency, as an insufficient oxygen supply may lead to tiredness, weakness, and an inability to perform daily activities. 7 Additionally, shortness of breath can occur easily in those with iron deficiency owing to reduced oxygen delivery to peripheral tissues. In severe cases, iron deficiency may lead to anemia, which significantly lowers hemoglobin levels in the blood. 8
A simple yet effective method to prevent or overcome iron deficiency is additional dietary iron supplementation. 9 However, a series of findings suggest that increased iron consumption may induce systemic copper deficiencies. 1,10 –12 Excessive iron intake can reduce serum and tissue copper levels, leading to growth retardation, cardiac hypertrophy, and decreased ceruloplasmin activity. 1,10 –13 However, when a diet high in iron is supplemented with extra copper, most abnormalities induced by high iron are significantly mitigated. 12 –14 Moreover, recent studies have shown that high iron consumption can also significantly alter lipid profiles. 15 When high dietary iron is accompanied by lower dietary copper, there is a significant increase in overall serum cholesterol levels. 16 Intriguingly, increasing dietary copper intake in conjunction with high iron intake can significantly and gradually reduce overall blood cholesterol levels. 16
Populations genetically predisposed to iron overload have an increased risk of developing cardiovascular diseases (CVDs). 17 In addition, excessive iron consumption may increase the risk of developing CVDs, as high iron intake is associated with cardiac hypertrophy and hypercholesterolemia. 17 At the cellular level, a study has shown that exposing primary human umbilical vein endothelial cells to excess iron (specifically, nontransferrin-bound iron) initiates de novo cholesterol synthesis, likely through the regulation of sterol regulatory element-binding protein 2. 18 Additionally, our previous study revealed that high iron combined with high copper levels reduced overall blood cholesterol. 16 Thus, the levels of dietary and cellular iron and copper play a crucial role in cholesterol metabolism, including cholesterol synthesis, uptake, and efflux. However, detailed genetic insights into how dietary iron and copper influence cholesterol metabolism remain unclear. To further investigate this mechanism, we analyzed the hepatic transcriptome following dietary interventions with six different concentrations of iron and copper.
The experimental protocols involving animals were reviewed and approved by the Institutional Animal Care and Use Committee (protocol number DKU-21–043) at Dankook University. Five-week-old male Sprague–Dawley rats, sourced from Doo Yeol Biotech, Inc., Seoul, Korea, underwent a week of acclimatization before being housed in stainless-steel cages with wire mesh bottoms and overhangs and were subjected to a five-week dietary intervention. This methodology was consistent with that of previous peer-reviewed research that highlighted the need for increased iron and copper intake during rapid growth phases. 12 The rats had ad libitum access to their specific diets, which were modified from the AIN-93G formula and prepared with distilled, mineral-free water. The diets varied in iron content (adequate iron [AdFe] or high iron [HFe]) and were supplemented with different levels of copper: low (LCu; 0.34 ppm), adequate (AdCu; 4.72 ppm), or high (HCu; 164.16 ppm). The iron concentration in the AdFe diet was increased from the standard of 39 ppm, as recommended by the AIN-93G guidelines, to approximately 120 ppm. Therefore, the AdFe in our experimental protocol may be near the upper limit used in clinical settings. The HFe in our experiment was set to over ∼78 times higher in concentration than AdFe and may not be applicable to clinical settings, but it is a value widely accepted in high dietary iron studies. 10 –16
For hepatic transcriptome analysis, the liver was removed and immediately frozen when the animals were euthanized. 19 –21 Total hepatic RNA was isolated following methods described in a prior study. 22 To prepare multiplexed libraries, 1 μg of DNase-treated total RNA was processed using the TruSeq Stranded Total RNA Kit from Illumina (San Diego, CA, USA). Next, the samples were treated with the Ribo-Zero Removal Kit (Illumina) to eliminate ribosomal RNA. Subsequently, the RNA was subjected to purification, fragmentation, reverse transcription, adapter ligation, and PCR amplification to enrich the DNA fragments. The prepared RNA-Seq libraries were then sequenced using a paired-end 150-base pair read length on an Illumina NovaSeq 6000 system. The conversion of bcl files to FASTQ format and the demultiplexing of samples were accomplished using Illumina’s BaseSpace.
After discarding low-quality sequencing reads (Trimmomatic v. 0.39), the quality-enhanced sequencing data were aligned to the reference genome (mRATBM7) using HISAT2 version 2.2.1. SAMtools was then employed to isolate unique reads, which were subsequently quantified using feature counts from the subread package (v. 2.0.3). Following the exclusion of genes exhibiting low expression, normalization of the data was performed employing the DESeq2 package in R software (v. 4.2.2). R and Python (v. 3.10) softwares were used to generate a heatmap that visually represented the significant differentially expressed genes.
The lipid panel data clearly indicated that a high-iron, low-copper (HFe/LCu) diet induced hypercholesterolemia. 16 Among the six different dietary interventions, only HFe/LCu significantly altered the hepatic transcriptome, specifically increasing the expression of genes related to cholesterol and lipid metabolism. Notably, HFe/LCu significantly decreased the expression of the hepatic ATP binding cassette subfamily G member 5 (ABCG5) gene (Fig. 1). The ABCG5 gene regulates intestinal absorption and promotes the biliary excretion of cholesterol; therefore, its downregulation contributes to the development of hypercholesterolemia. 23 Additionally, the HFe/LCu group exhibited deceased expressions of catabolic genes involved in lipid metabolism, such as progestin and adipoQ receptor (PAQR), 2,4-dienoyl-CoA reductase (DECR1), and carnitine O-octanoyltransferase (CROT), indicating diminished lipid catabolism (Fig. 1, Table 1). Furthermore, mice that were fed the HFe/LCu diet exhibited cardiac hypertrophy, a key pathological feature of CVD. This condition can be attributed to high iron-induced copper deficiency. 24 Hepatic transcriptome analysis also revealed significant decreases in carnitine O-acetyltransferase (CRAT) and steroid sulfatase (STS) expression, indicating that these genes are involved in the development of cardiac function abnormalities associated with the HFe/LCu diet (Fig. 1, Table 1).
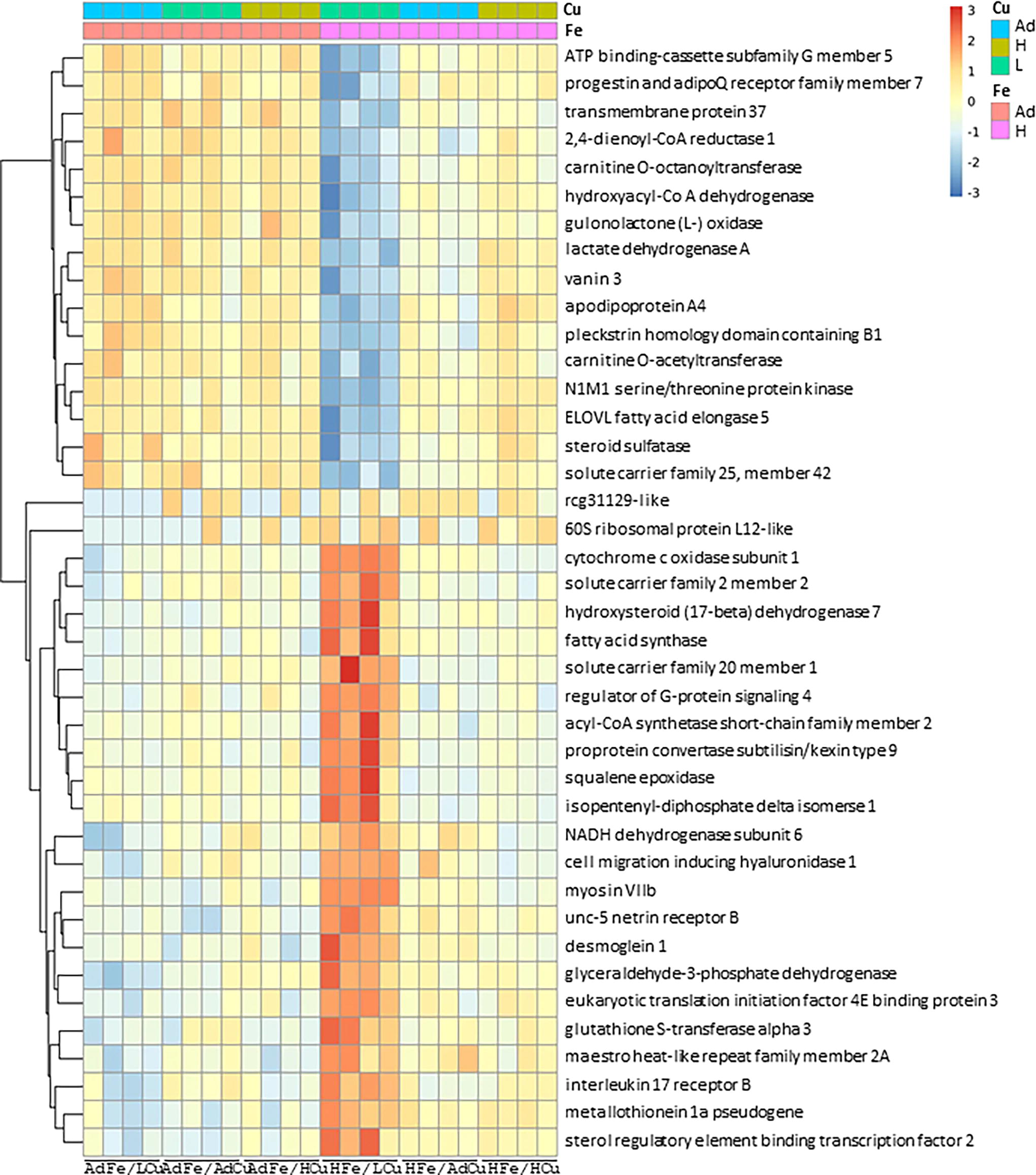
Screening and identification of DEGs using bioinformatics. Five-week-old male SD rats were fed one of the six diets, which varied only in their iron and copper content, for five weeks ad libitum. The hepatic transcriptome was analyzed and expressed as a heatmap of DEGs associated with fold differences. Ad, adequate; DEG, differentially expressed gene; H, high; L, low; SD, Sprague–Dawley.
Hepatic Genes Expressions Decreased by High-Iron, Low-Copper Supplementation as Identified by Transcriptome Analysis
The HFe/LCu diet not only induced dyslipidemia but also had a significant impact on the hepatic transcriptome, particularly affecting genes involved in energy metabolism and electron transfer. Consumption of the HFe/LCu diet led to increased expression of genes associated with de novo cholesterol and lipid synthesis. Specifically, mice on the HFe/LCu diet exhibited enhanced expression of key catabolic genes involved in hepatic lipid synthesis, such as fatty acid synthase (FAS) and acyl-CoA synthetase short-chain family member 2 (ACSS2) (Fig. 1, Table 2). Remarkably, mice on the HFe/LCu diet also exhibited upregulation of genes crucial for de novo cholesterol synthesis, including proprotein convertase subtilisin/kexin type 9 (PCSK9), sterol regulatory element-binding transcription factor 2 (SREBP-2), squalene epoxidase (SQLE), and isopentenyl-diphosphate delta isomerase (IPI) (Fig. 1, Table 2). The increase in PCSK9 and SREBP-2 directly supports the synthesis of novel cholesterol molecules. 25 Moreover, the upregulation of SQLE and IPI highlights their role as rate-limiting steps in sterol and isoprenoid synthesis in the liver, respectively (Table 2). 18,26,27 In addition to promoting cholesterol synthesis, consumption of the HFe/LCu diet also increased the expression of antioxidative genes, such as cytochrome c oxidase subunit 1 (COX1) and glutathione S-transferase alpha 3 (GSTA3). These genes are crucial in radical scavenging and detoxification, suggesting that the HFe/LCu diet may induce hepatic oxidative stress.
Hepatic Genes Expressions Increased by High-Iron, Low-Copper Supplementation as Identified by Transcriptome Analysis
Several scientific studies have suggested that elevated iron levels can lead to hypercholesterolemia. One study reported that exposing primary human umbilical vein endothelial cells to excess iron, specifically nontransferrin-bound iron, initiated the synthesis of cholesterol, likely through the activation of sterol regulatory element-binding protein 2. 18 Additionally, high levels of iron exposure have been linked to cellular apoptosis, potentially driven by an increase in tumor necrosis factor alpha (TNFα). 18 This association was confirmed using the cholesterol chelator hydroxypropyl-β-cyclodextrin, which has been shown to mitigate TNFα-induced apoptosis caused by excessive iron. 18 Based on our previous study, we proposed that consuming large amounts of iron might initially stimulate cholesterol synthesis, as indicated in earlier research. 18 Through analysis of the hepatic transcriptome, we found that high iron intake increases de novo cholesterol synthesis through the upregulation of genes such as PCSK9, SREBP-2, SQLE, and IPI. High iron intake also reduces cholesterol elimination through biliary excretion by downregulating ABCG5.
Scientific interest has expanded to include the effects of copper supplementation in conjunction with high iron consumption, which has been observed to gradually reduce cholesterol levels and normalize the hepatic transcriptome in comparison with a balanced iron and copper diet (AdFe/AdCu). Specifically, high-iron diets are known to trigger hypercholesterolemia by inducing the expression of genes involved in cholesterol synthesis and reducing the expression of genes related to biliary excretion. 28 However, the addition of extra copper has been shown to mitigate the effects of high iron on the expression of hepatic genes related to blood cholesterol levels. 16 Dynamic changes in blood cholesterol levels may be associated with copper deficiency induced by high iron intake. 16 An additional significant observation was that mice on the HFe/LCu diet exhibited increased expression of the metallothionein 1A pseudogene, a response typically observed in copper deficiency. Thus, copper supplementation may help reduce overall cholesterol synthesis in the liver. However, to fully understand the precise roles and detailed molecular mechanisms of iron and copper in cholesterol metabolism, further comprehensive studies are needed under various pathological conditions.
Footnotes
AUTHORS’ CONTRIBUTIONS
Conceptualization: B.-H.K. and J.-H.H.; methodology: J.L., H.J., and J.-H.H.; software: J.L., H.J., M.D., B.-H.K., and J.-H.H.; validation: J.L., H.J., M.D., B.-H.K., and J.-H.H.; formal analysis: J.L. and J.-H.H.; resources: B.-H.K. and J.-H.H.; data curation: J.L., H.J., and J.-H.H.; writing—original draft preparation: J.L., H.J., and J.-H.H.; writing—review and editing: J.L., H.J., M.D., B.-H.K., and J.-H.H.; visualization: J.L. and H.J.; supervision: B.-H.K. and J.-H.H.; project administration: B.-H.K. and J.-H.H.; funding acquisition: J.-H.H. All authors have read and agreed to the published version of the article.
AUTHOR DISCLOSURE STATEMENT
No interests to disclose.
FUNDING INFORMATION
This research was supported by the Basic Science Research Program through the