
Abstract
Fermented red ginseng (FRG) enhances the bioactivity and bioavailability of ginsenosides, which possess various immunomodulatory, antiaging, anti-obesity, and antidiabetic properties. However, the effects of FRG extract on muscle atrophy and the underlying molecular mechanisms remain unclear. This study aimed to elucidate the effects of FRG extract on muscle atrophy using both in vitro and in vivo models. In vitro experiments used dexamethasone (DEX)-induced C2C12 myotubes to assess cell viability, myotube diameter, and fusion index. In vivo experiments were conducted on hind limb immobilization (HI)-induced mice to evaluate grip strength, muscle mass, and fiber cross-sectional area (CSA) of the gastrocnemius (GAS), quadriceps (QUA), and soleus (SOL) muscles. Molecular mechanisms were investigated through the analysis of key signaling pathways associated with muscle protein synthesis, energy metabolism, and protein degradation. FRG extract treatment enhanced viability of DEX-induced C2C12 myotubes and restored myotube diameter and fusion index. In HI-induced mice, FRG extract improved grip strength, increased muscle mass and CSA of GAS, QUA, and SOL muscles. Mechanistic studies revealed that FRG extract activated the insulin-like growth factor 1/protein kinase B (Akt)/mammalian target of rapamycin signaling pathway, promoted muscle energy metabolism via the sirtuin 1/peroxisome proliferator-activated receptor gamma-coactivator-1α pathway, and inhibited muscle protein degradation by suppressing the forkhead box O3a, muscle ring-finger 1, and F-box protein (Fbx32) signaling pathways. FRG extract shows promise for ameliorating muscle atrophy by modulating key molecular pathways associated with muscle protein synthesis, energy metabolism, and protein degradation, offering insights for future drug development.
INTRODUCTION
Regulation of protein synthesis and degradation maintains dynamic equilibrium and stability in skeletal muscle tissue, contributing to skeletal muscle plasticity. 1 When protein degradation rate is significantly faster than synthesis the rate, there is a net loss of muscle tissue, thus decreasing muscle strength and muscle fiber cross-sectional area (CSA), thereby triggering muscle atrophy. 2,3 It is known that the expression of the insulin-like growth factor 1 (IGF-1)/protein kinase B (Akt)/mammalian target of rapamycin (mTOR) pathway affects protein synthesis, 4 –6 and the ubiquitin-proteasome system has a significant impact on protein degradation. 7,8 Furthermore, mitochondrial oxidative metabolism and energy transduction are essential for maintaining muscle function and health. 9,10
Red ginseng mainly contains various amino acids, saponins (such as ginsenosides), and non-saponins. 11 Although red ginseng contains abundant ginsenosides, their absorption in the intestine is poor. Intestinal bacteria must metabolically transform them through processes such as deglycosylation and fatty acid esterification to convert them into biologically active forms. 12,13 The currently known methods for producing fermented red ginseng (FRG) involve either inoculating microorganisms or treating with enzymes isolated from microorganisms. Fermenting red ginseng using these methods has been reported to generate specific ginsenosides and enhance various bioactive compounds. Microbial fermentation has been shown to convert major ginsenosides (Rb1, Rb2, Rc, Rd, Re, and Rg1) into minor ginsenosides (Rg3, Rh2, compound K, Rg2, and Rh1) and further degrade them into small molecules, thereby significantly enhancing endogenous ginsenoside absorption and improving their overall efficacy. 14,15 FRG contains a diverse array of ginsenosides. 16,17 These components exhibit a wide range of biological activities. 18 –22
Although various biological activities of FRG have been extensively studied, its efficacy and mechanisms in ameliorating muscle atrophy are not well-studied. Therefore, this study aimed to thoroughly investigate the potential effects of FRG extract on muscle atrophy and elucidate its underlying molecular mechanisms using both dexamethasone (DEX)-induced C2C12 cells and a hind limb immobilization (HI)-induced muscle atrophy mouse model. Through our results, we hope to reveal the potential effect of FRG extract in alleviating muscle atrophy and provide scientific support for the future treatment of muscle-related diseases.
MATERIALS AND METHODS
Chemicals
Dulbecco’s modified Eagle’s medium (DMEM), phosphate-buffered saline (PBS), horse serum (HS), penicillin-streptomycin (P/S), and fetal bovine serum (FBS) were purchased from Gibco (Grand Island, NY, USA). 2,3-bis(2-methoxy-4-nitro-5-sulfophenyl)−2H-tetrazolium-5-carboxanilide (XTT) and N-methyldibenzopyrazine methyl sulfate (PMS) were purchased from Welgene (Gyeongsan, Korea). Ginsenosides Rg2 and Rh1 were obtained from MedChem Express (Monmouth Junction, NJ, USA). DEX and the ginsenoside Rg3 were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Preparation of FRG extract
FRG extract was provided by F&B Bio Co., Ltd. (Cheonan, Korea). Ginseng was steamed at 92 ± 2°C for 1 h, kept warm for 30 min, and then dried for 3 h in a hot air dryer (HD-300, Korean Dry Tech, Jincheon, Korea) at 60 ± 2°C with 20–30% moisture content. This process was repeated three times, and the ginseng was extracted in 50% ethanol at 1:20 (v/v) and 95 ± 2°C for 24 h. The extract was filtered and concentrated under reduced pressure to obtain the concentrated solution (22 ± 2 Brix), which was treated at 40 ± 5°C for 3 h using Sumizyme AC and Cellulsin AL8 (3%) derived from Aspergillus niger and fermented at 40 ± 5°C for 24 h. Subsequently, it was sterilized at 95 ± 2°C, resulting in the FRG extract. The types and contents of ginsenosides in FRG extract were determined using ultra-performance liquid chromatography (Supplementary Table S1 and Supplementary Table S2).
Cell culture and differentiation
C2C12 myoblasts were maintained in DMEM supplemented with 1% P/S and 10% FBS. The cells were incubated at 37°C in a 5% CO2 atmosphere and the medium was refreshed every two days. Upon reaching 80–90% confluence, the medium was replaced with DMEM containing 1% P/S and 2% HS to induce differentiation. We have previously demonstrated that a muscle atrophy cell model can be successfully established by 200 μM DEX treatment. 3 Therefore, after cell differentiation for 5 days, the cells were co-treated with a medium containing 200 μM DEX and different FRG extract concentrations (50–200 μg/mL) for 24 h. Following the treatment, the cells were collected for subsequent experiments.
Cell viability assay
C2C12 myoblasts were cultured in a 96-well plate and media to induce differentiation were added to the wells. Subsequently, they were treated with 200 μM DEX and/or different FRG extract concentrations (50–200 μg/mL) for 24 h. Then, a 1:50 (v/v) PMS and XTT mixture were added to each well, followed by incubation at 37°C for 4 h. Finally, the absorbance changes at 450 and 690 nm were measured using a microplate reader (Molecular Devices, Sunnyvale, CA, USA) to evaluate cell viability.
Jenner–Giemsa staining
Methanol was slowly added to each well for cell fixation for 5 min and dried at room temperature for 10 min. The dried cells were stained with Jenner staining solution for 5 min, the staining solution was removed, and the cells were washed twice with deionized water (DW). The cells were then stained with Giemsa staining solution for 10 min and washed twice with DW. Finally, the plates were observed under an inverted microscope (CKX41SF; Olympus Co. Ltd., Tokyo, Japan) and imaged.
Jenner staining solution preparation: The Jenner staining reagent was mixed with methanol at 1:0.4 (mg/mL), and the resulting solution was combined with DW at 1:1 (v/v). This mixture was combined with 1 mM sodium phosphate buffer (pH 5.6) at 1:3 (v/v) and filtered to obtain the final solution.
Giemsa staining solution preparation: The Giemsa staining reagent was mixed with 1 mM sodium phosphate buffer (pH 5.6) at 1:20 (v/v) and filtered to obtain the final solution.
C2C12 myotube diameter and fusion index measurement
The stained cells were randomly imaged at 200 × magnification under an inverted microscope. Five random images were captured from each well, and the ImageJ software was used to calculate and measure the diameter of the myotube cells containing three or more nuclei. The fusion index was calculated using the following formula:
Animal and experimental design
All experimental animal protocols were reviewed and approved by the Institutional Animal Care and Use Committee of the Kangwon National University (Approval Number: KW-221025-1). Five-week-old male C57BL/6J mice were obtained from DBL Co. Ltd. (Eumseong, Korea) and maintained in a controlled environment at 55 ± 5% relative humidity and 23 ± 2°C with a 12 h light/dark cycle.
Before the experiment, all mice were acclimated for one week with free access to water and food. Subsequently, the mice were randomly divided into two main groups: normal (n = 5) and HI (n = 30). After one week of HI, a mouse model of muscle atrophy was successfully established. The HI-induced mice were further divided into six experimental groups: HI control group (n = 5); 200 mg/kg/day Schisandra chinensis extract (SE) treatment group (SE 200, n = 5); 15 mg/kg/day mixed ginsenosides Rh1, Rg2, and Rg3 (MIX) treatment group (MIX 15, n = 5); 50 mg/kg/day FRG extract treatment group (FRG 50, n = 5); 100 mg/kg/day FRG extract treatment group (FRG 100, n = 5); and 200 mg/kg/day FRG extract treatment group (FRG 200, n = 5).
Since SE enhances muscle strength, 6 it was used as a positive control in this study. Ginsenosides Rh1, Rg2, and Rg3 mitigate muscle atrophy. 3 In this experiment, the MIX group was included to investigate the synergistic effects of the assorted ginsenosides present in the fermented red mountain-cultivated ginseng, enhancing our understanding of their potential in combating muscle atrophy.
In this experiment, mice hind limbs were continuously immobilized, and oral administration was conducted for two weeks. Throughout this period, grip strength and body weight were assessed weekly. After two weeks, the mice were anesthetized using isoflurane, and muscle tissues such as the quadriceps (QUA), gastrocnemius (GAS), and soleus (SOL) were harvested from the immobilized hind limbs, weighed, photographed, and stored at −80°C for subsequent analysis.
Grip strength test
The grip strength of mice was assessed using a grip strength meter (JD-A-22, JEUNG DO BIO & PLANT Co. Ltd., Seoul, Korea). The mice were placed on a mesh platform connected to a force meter to ensure that all four limbs grasped the mesh. Maximal grip strength was determined by horizontally pulling the mouse tail backward. Each mouse underwent five trials, and the data were recorded. Grip strength results were normalized to the weight of the mice.
Histological analysis
The QUA and GAS muscles were fixed in 4% paraformaldehyde, embedded in paraffin, and sectioned into 5 μm thick sections. Subsequently, the sections were stained with hematoxylin and eosin (H&E) stain. Afterward, the muscle fiber CSA were imaged using an optical microscope (Carl Zeiss, Germany) and quantified using ImageJ software.
Western blot analysis
Cells and muscle tissue were lysed using a protein lysis buffer (0.25% sodium deoxycholate, 0.1% sodium dodecyl sulfate, 1% Nonidet P-40, 150 mM sodium chloride, and 50 mM Tris-HCl) and centrifuged at 12,000 × g and 4°C for 20 min. After blocking with Tris-buffered saline containing Tween-20 containing 5% skim milk for 1 h, the membranes were incubated for 12 h at 4°C with primary antibodies (forkhead box O3a [FoxO3a], p-mTOR, p-Akt, mTOR, Akt, sirtuin 1 [SIRT1]) and β-actin (1:1000; Cell Signaling Technology, Danvers, MA, USA); muscle ring-finger 1 (MuRF1) and peroxisome proliferator-activated receptor gamma-coactivator-1α (PGC-1α) (1:1000; Santa Cruz Biotechnology, Dallas, TX, USA); F-box protein (Fbx32); and IGF-1 (1:1000; Abcam, Cambridge, UK)]. Subsequently, the membranes were incubated with mouse or rabbit secondary antibodies (1:2000) at room temperature for 1 h. Protein bands were detected using the ECL detection reagent on a ChemiDoc imaging system (Bio-Rad Laboratories, Inc., Hercules, CA, USA), and protein expression levels were quantified using ImageJ software.
Statistical analysis
Data are presented as means ± standard deviations. Statistical analysis was conducted using SPSS software (version 24.0; SPSS Inc., Chicago, IL, USA). Duncan’s multiple range test was used to assess the significance among experimental groups. P < 0.05 was considered statistically significant.
RESULTS
Effect of FRG extract on myotube atrophy in DEX-induced C2C12 myotubes
The MIX, SE, and FRG extract concentrations that were tested were not cytotoxic (Fig. 1A). Compared with that of the control (CON) group myotubes, the viability of 200 μM DEX-treated myotubes were significantly reduced. However, the viability of DEX-treated cells significantly improved upon treatment with SE, MIX, or FRG extract (Fig. 1B). Representative images showed C2C12 myotubes stained with Jenner–Giemsa (Fig. 1C). Compared with that of CON group myotubes, the diameter and fusion index of DEX-treated myotubes were significantly reduced. However, SE, MIX, or FRG extract treatment effectively counteracted the DEX-induced reductions (Fig. 1D,E).
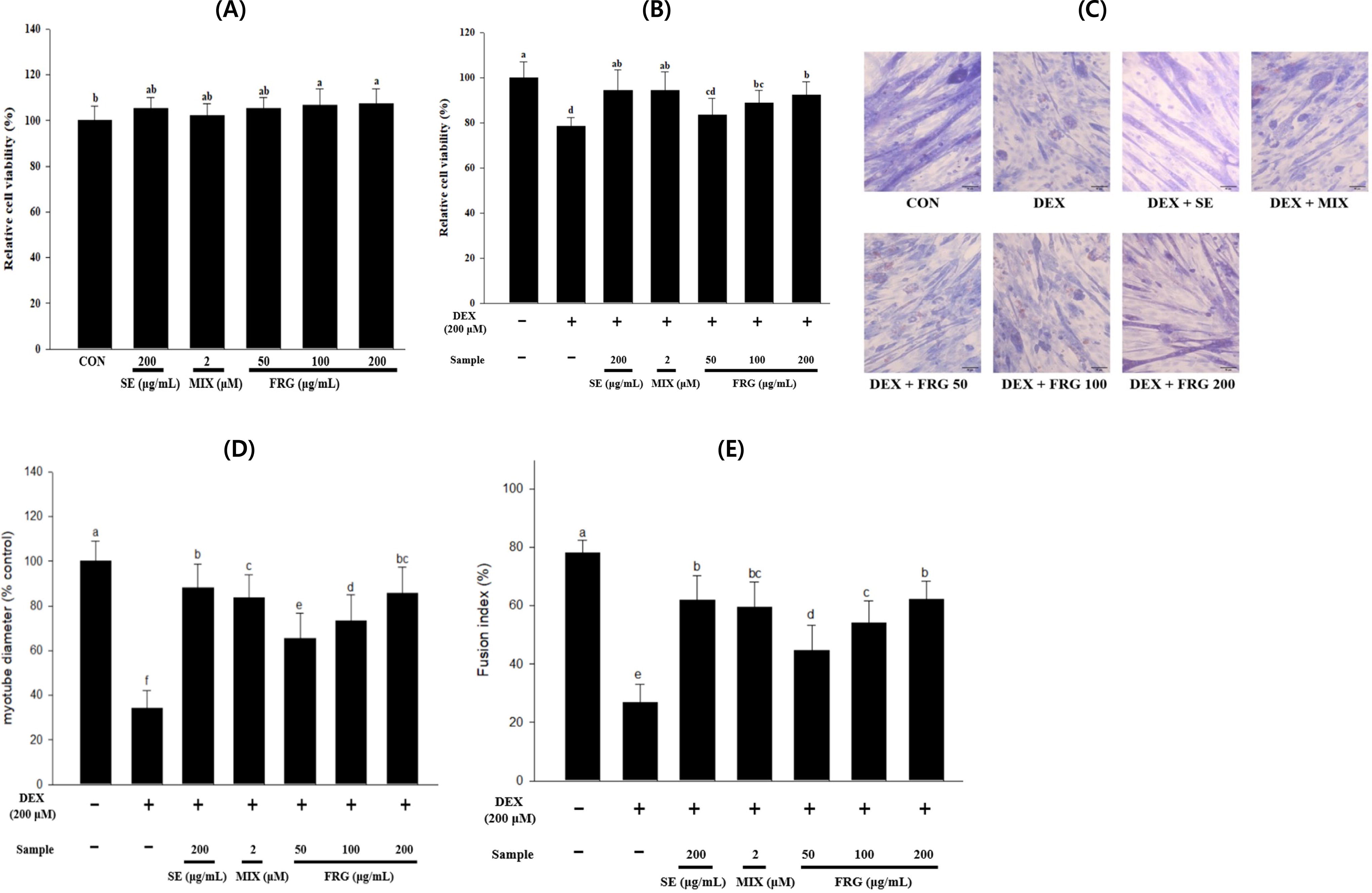
Effects of SE, MIX, and FRG extract on DEX-induced atrophy in C2C12 myotubes. We evaluate the viability of C2C12 myotubes after
Effect of FRG extract on protein expression in DEX-induced atrophy of C2C12 myotubes
The IGF-1 expression levels in the DEX group were significantly lower than that in the CON group, and Akt and mTOR phosphorylation was also significantly inhibited (Fig. 2A–C). However, FRG extract treatment restored their levels dose-dependently. The IGF-1 expression levels and Akt and mTOR phosphorylation levels were comparable with or even higher than those in the SE and CON groups (Fig. 2A–C).
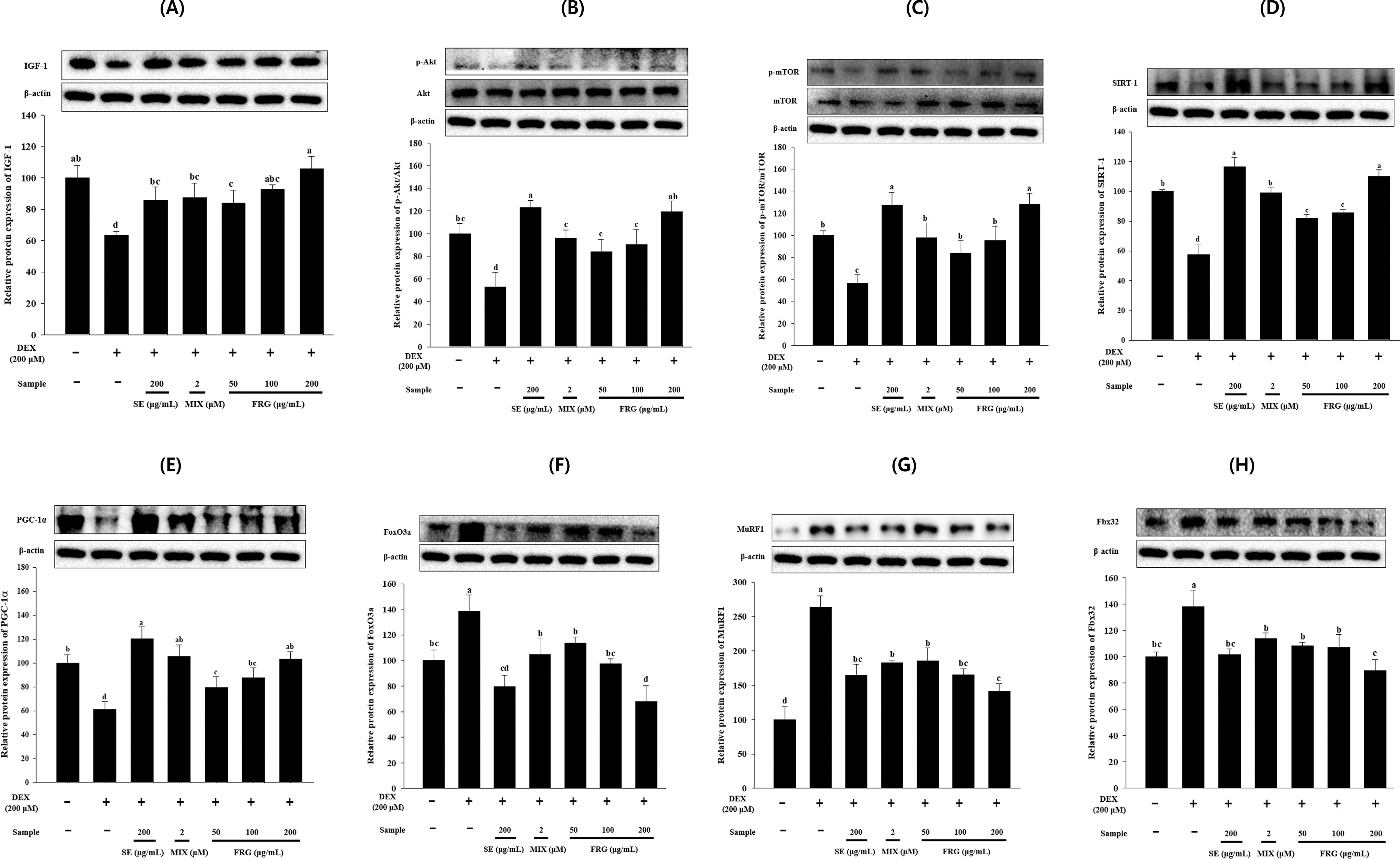
Effect of FRG extract on the expression of muscle protein synthesis-related proteins [
The SIRT1 protein expression levels in DEX-treated C2C12 myotubes were significantly decreased compared with those in the CON myotubes. However, FRG extract treatment significantly increased SIRT1 and PGC-1α expression levels in DEX-treated cells concentration dependently (Fig. 2D,E). Notably, both SIRT1 and PGC-1α protein expression levels in the FRG 200 group were comparable to, or even higher than those in the CON and SE groups (Fig. 2D,E).
The results of the expression of muscle protein degradation-related proteins showed that their expression in the DEX-treated group were significantly increased compared with those in the CON group (Fig. 2F–H), indicating that DEX treatment promoted the expression of muscle protein degradation-related proteins, leading to myotube atrophy. However, the DEX-induced expression levels of these proteins were significantly reduced in the FRG extract-treated groups (Fig. 2F–H).
FRG extract influences muscle strength, size, and mass in HI-induced muscle atrophy mice
One week after HI induction, the grip strengths of HI group mice were significantly lower than that of normal group mice, indicating successful muscle atrophy induction. To evaluate the potential effect of FRG extract on muscle atrophy, a muscle atrophy model was established by immobilizing the hind limbs of mice for 1 week. After 2 weeks of oral administration, the grip strength of the HI group mice remained significantly lower than that of the normal group mice and continued to decline during the experiment. However, FRG extract treatment significantly improved grip strength in mice with HI-induced muscle atrophy (Fig. 3A). We selected these three muscles to identify the therapeutic effects of FRG extract on muscle atrophy. The QUA, GAS, and SOL muscle volumes in the HI group mice were significantly lower than those in the normal group mice. However, their sizes increased after FRG extract treatment (Fig. 3B). The muscle mass of the QUA, GAS, and SOL fibers in the HI group mice was significantly reduced by 32%, 40%, and 54%, respectively, then those in the normal group mice (Fig. 3C–E). However, this effect was markedly ameliorated in the FRG extract-treated groups.

FRG improved HI-induced muscle atrophy in mice, enhancing muscle strength, skeletal muscle volume, and mass. Mice were treated with SE 200, MIX 15, or FRG 50, 100, and 200 for two weeks after HI for 1 week.
FRG extract affects muscle fiber CSA in HI-induced muscle atrophy mice
Through histological analysis and fiber CSA quantification, we observed that the QUA and GAS muscle fiber CSA of HI group mice was significantly lower than those of the normal group mice. However, HI-induced CSA reduction was significantly reversed after treatment with SE 200, MIX 15, and FRG 50–200. Furthermore, the CSA of QUA and GAS myofibers enhanced FRG extract concentration dependently (Fig. 4A,B).

FRG ameliorates HI-induced reduction in muscle fiber CSA.
Effect of FRG extract on protein expression in HI-induced muscle atrophy mice
The expression of the key protein IGF-1, which is involved in muscle protein synthesis, was significantly inhibited in HI-induced QUA and GAS muscle tissues (Fig. 5A,B). Treatment of FRG 200 significantly promoted their expression, which were similar to the effects of SE 200. In addition, the p-Akt/Akt and p-mTOR/mTOR ratios in the HI group significantly decreased, indicating that HI reduced muscle protein synthesis by inhibiting Akt and mTOR phosphorylation in QUA and GAS tissues. However, FRG extract treatment significantly enhanced HI-induced Akt and mTOR phosphorylation in muscle tissues.
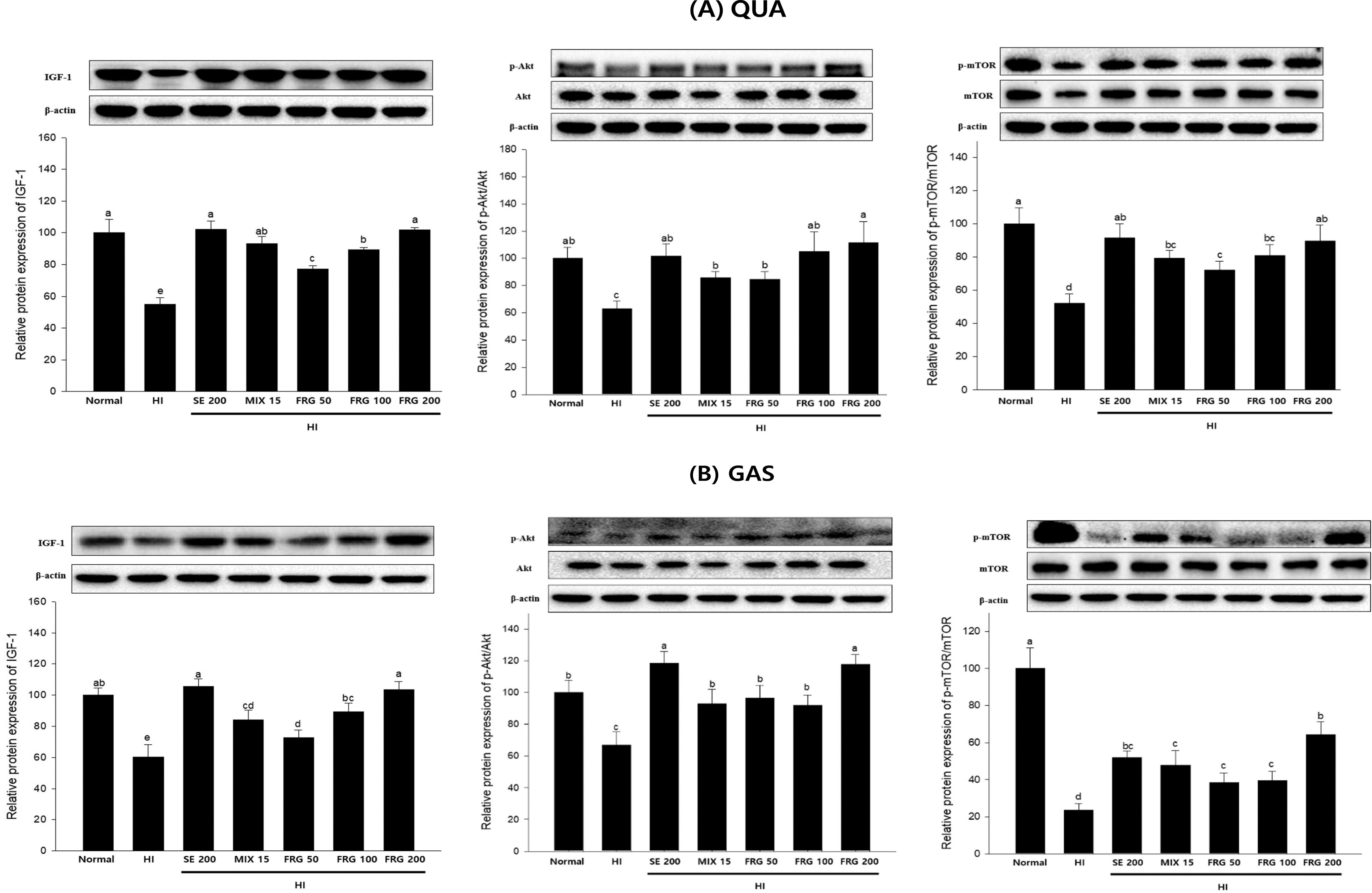
Expression levels of IGF-1/Akt/mTOR, a protein involved in muscle protein synthesis, in
We further investigated the impact of FRG extract on the SIRT1/PGC-1α signaling pathway in HI-induced QUA and GAS muscle tissues. The results showed that SIRT1 and PGC-1α protein expression in QUA and GAS muscles of HI group mice was significantly reduced than that in those of the normal group mice. In contrast, MIX 15 and FRG 50, 100, 200 treatment significantly increased SIRT1 and PGC-1α expression in these two muscles (Fig. 6A,B).
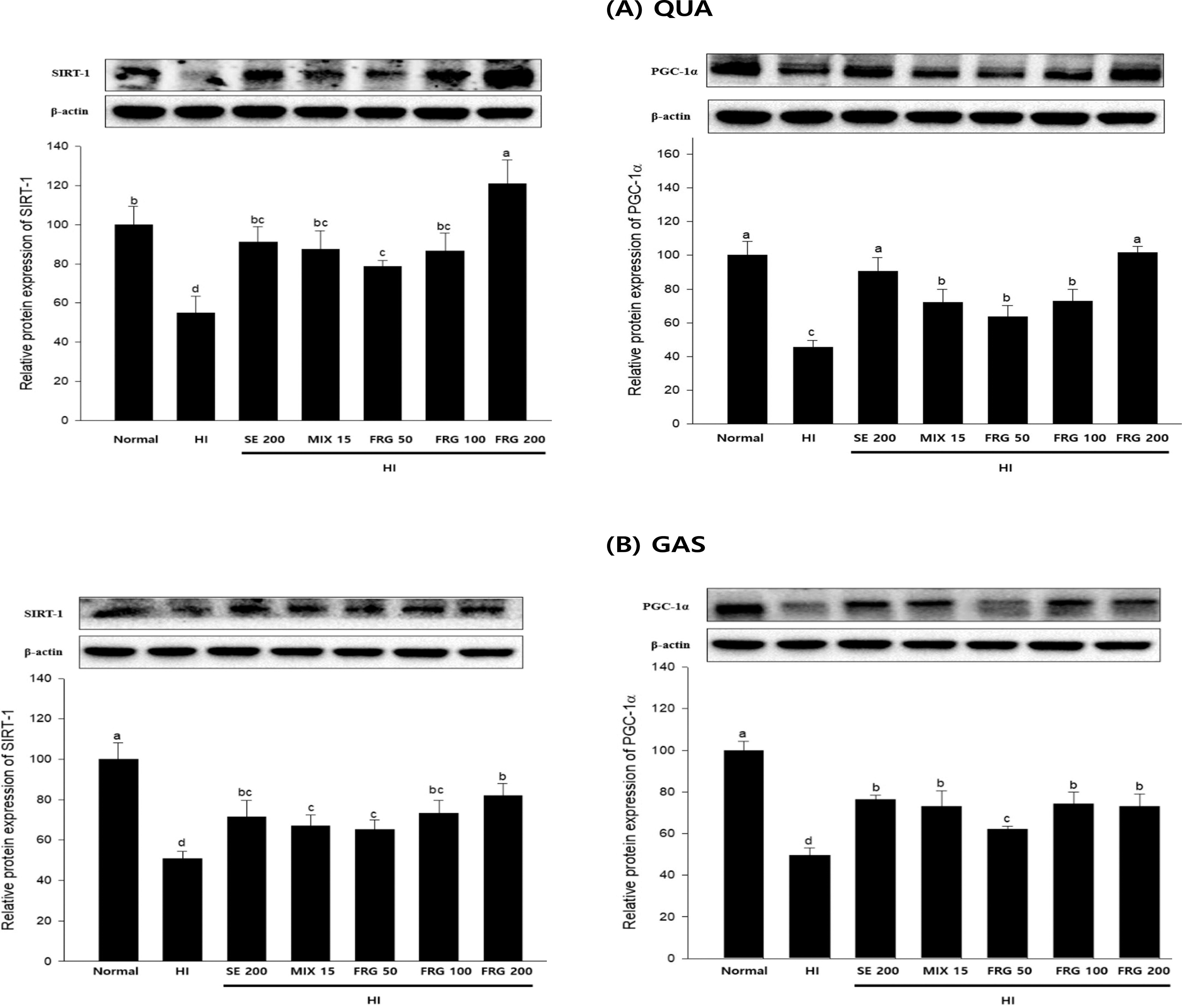
FRG restores HI-induced muscle atrophy through the SIRT1/PGC-1α pathway. SIRT1 and PGC-1α protein expression levels in
FoxO3a, MuRF1, and Fbx32 protein expressions in both muscle types were significantly increased and decreased in the HI and FRG groups, respectively, compared with those in the normal group (Fig. 7A,B). Overall, these data indicated that FRG extract effectively prevented HI-induced muscle atrophy by promoting muscle protein synthesis, activating energy metabolism-related pathways, and suppressing muscle degradation-related protein expression.
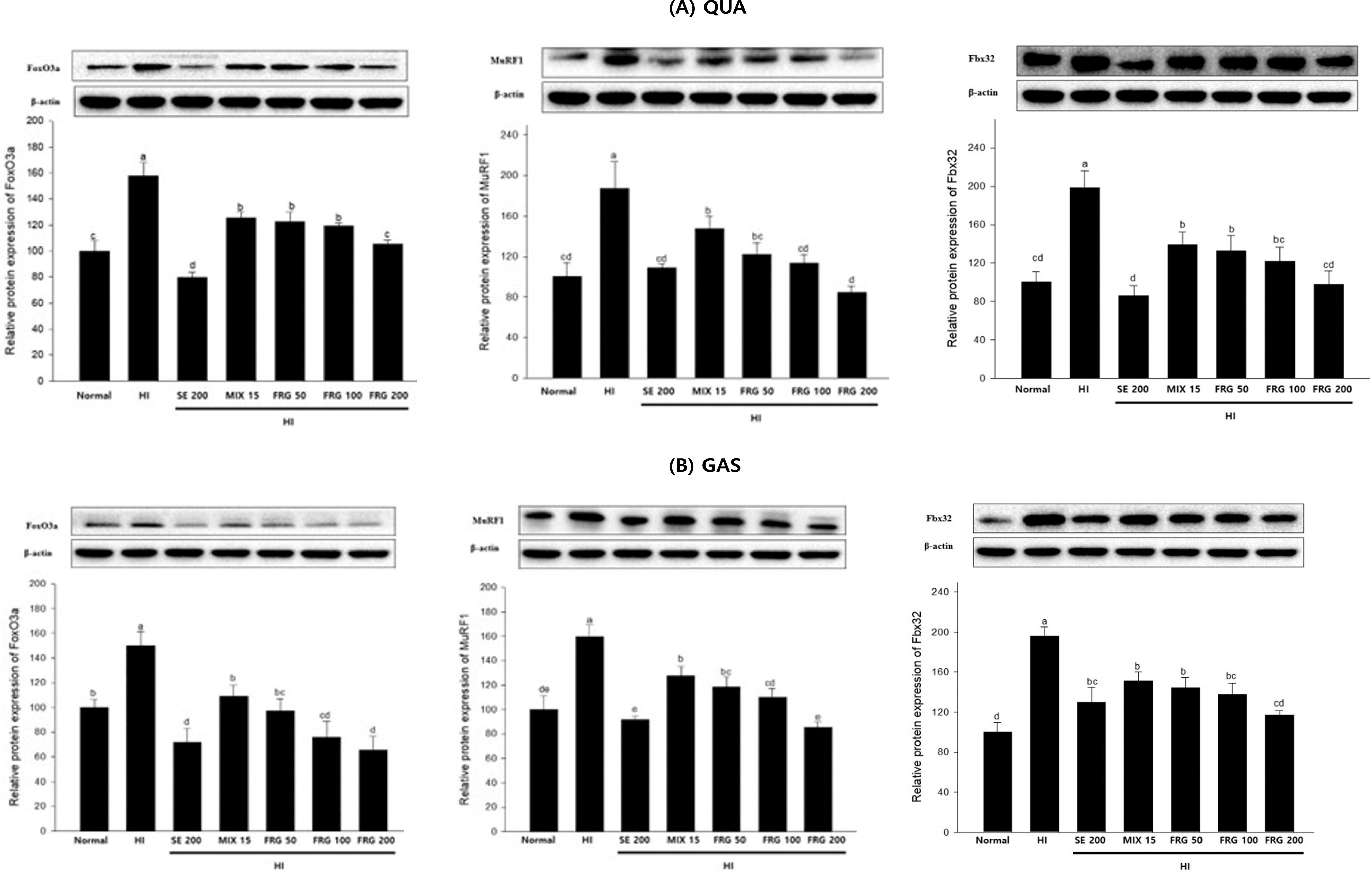
Impact of FRG on the expression of the muscle atrophy-related proteins FoxO3a, MuRF1, and Fbx32 in HI-induced muscle atrophy mice. FoxO3a, MuRF1, and Fbx32 protein expression levels in
DISCUSSION
During myoblast differentiation, fusion forms myotubes, which subsequently merge with others, increasing both their number and diameter, enhancing the fusion index. 23 We have previously indicated that treating C2C12 myotubes with 200 μM DEX significantly decreased cell viability, diameter, and fusion index. 3 In addition, HI markedly decreases muscle strength and mass in mice. 24 Therefore, in this study, we assessed the therapeutic effects of FRG extract on muscle atrophy in both DEX-induced C2C12 myotube atrophy model and HI-induced muscle atrophy mouse model and explored its mechanism of action.
In the DEX group, cell viability, myotube diameter, and fusion index of C2C12 myotubes were significantly inhibited. Furthermore, HI-induced mice decreased muscle strength and muscle mass loss. In addition, histological studies of the HI-treated mouse muscle tissue revealed that the CSA of the QUA and GAS muscle tissues were significantly reduced, but were significantly improved by FRG extract treatment. In vitro experiments demonstrated that FRG extract significantly reversed the DEX-induced decrease in cell activity, myotube diameter, and fusion index. FRG extract treatment significantly enhanced muscle strength in vivo and effectively restored muscle mass and CSA in HI-induced mice. We also confirmed the efficacy of antimuscle atrophy using MIX, a mixture containing major compounds fermented red mountain-cultivated ginseng extract, namely ginsenoside Rh1, Rg2, and Rg3. 3
Muscle tissue plasticity hinges on adjusting protein synthesis and degradation rates. During muscle atrophy, an imbalance arises, with protein synthesis rate markedly lower than degradation rate, culminating in net muscle protein loss and subsequent atrophy. 25 Recent studies have suggested that muscle atrophy can be treated by modulating signaling pathways, such as muscle protein synthesis, muscle degradation, and mitochondrial function. 26
IGF-1, a well-researched growth factor, plays a pivotal role in stimulating cellular growth and differentiation, especially in muscle tissues. It triggers Akt activation by binding to its dedicated receptor, thereby facilitating further activation of mTOR. mTOR serves as a crucial regulatory factor, promoting protein biosynthesis through the activation of muscle-specific translation initiation factors. 27 Also, Akt inhibits protein degradation via phosphorylation and location of FoxO3a from the nucleus to the cytosol. 28,29 Therefore, the major role of IGF-1/Akt/mTOR in muscle atrophy is to inhibit protein degradation factors and increase protein synthesis. In this study, we observed that FRG extract treatment significantly enhanced the expression of muscle synthesis-related proteins in DEX-induced C2C12 myotubes and HI-induced atrophied muscle in mice.
FoxO3a is a key transcription factor involved in regulating muscle protein degradation and promotes protein ubiquitination and subsequent proteasomal degradation by activating specific ubiquitin ligases such as MuRF1 and Fbx32. 4 FRG extract treatment suppressed FoxO3a expression both in vivo and in vitro, thus downregulating the downstream target proteins MuRF1 and Fbx32, effectively ameliorated DEX- and HI-induced muscle atrophy.
To thoroughly examine how FRG extract improves muscle atrophy, we delved into the molecular mechanisms associated with mitochondrial biogenesis and their influence on muscle strength. SIRT1 has the capacity to stimulate PGC-1α, thereby fostering mitochondrial biogenesis and bolstering muscle energy levels. 4,30 In this study, FRG extract significantly upregulated SIRT1 and PGC-1α expression levels, indicating that FRG extract promotes mitochondrial biogenesis by regulating energy metabolism-related proteins, thereby preventing muscle atrophy.
CONCLUSION
This study aimed to describe the effects of FRG extract on muscle atrophy using both in vitro and in vivo models, demonstrating that FRG extract significantly improved muscle atrophy in both cellular and animal models. By modulating IGF-1/Akt/mTOR pathway, inhibiting FoxO3a/MuRF1/Fbx32, and activating SIRT1/PGC-1α, FRG extract provides a comprehensive and promising approach for treating muscle atrophy. This study not only provides crucial insights into the molecular mechanisms of muscle atrophy. This holds the prospect of bringing positive and far-reaching impacts to the field of muscle health.
Footnotes
ACKNOWLEDGMENT
The authors thank the study researchers for contributing to this study.
AUTHORS’ CONTRIBUTIONS
X.M.: Writing—original draft preparation and validation. X.H.: Visualization and software. I.-J.L.: Resources and methodology. S.-J.L.: Formal analysis. G.O.: Validation and investigation. J.-H.I.: Methodology and formal analysis. X.F.: Investigation. J.-S.L.: Software and visualization. K.S.B.: Project administration, conceptualization, and supervision. G.-S.S.: Conceptualization and supervision. D.-S.L.: Conceptualization and supervision. S.C.: Conceptualization and supervision. O.-H.L. Writing—review and editing, conceptualization, and supervision.
AUTHOR DISCLOSURE STATEMENT
The authors declare no conflict of interest.
FUNDING INFORMATION
This research was funded by the
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.