
Abstract
Diabetic nephropathy (DN) is a prevalent complication and serious microvascular of diabetes mellitus. After previous studies, we found that phenylethanol glycosides (CPhGs) derived from Cistanche tubulosa (Schenk) Wight exerts antidiabetic and renoprotective effects. However, the effects of CPhGs on DN remain incompletely understood. The study aimed to examine the effects of CPhGs on DN in rats and explore the underlying mechanism involved. A DN rat model was established by streptozotocin (STZ) combined with a high-fat diet. Reagent kits were used to assess the extent to which CPhGs ameliorate hyperglycemia, insulin resistance (IR), renal dysfunction, kidney oxidative stress, and peripheral inflammation. Histology and immunohistochemical staining were used to detect the changes in renal tissue structure and the expression levels of α-smooth muscle actin (α-SMA) and collagen I. Furthermore, we analyzed the cecal contents of DN rats to investigate the effect of CPhGs on gut microbiota by using 16S rRNA sequencing and broad-spectrum metabolite profiling. The results showed that CPhGs demonstrated a range of advantageous outcomes in DN, encompassing the enhancement of kidney function and alleviation of hyperglycemia, IR, renal injury, oxidative stress, and peripheral inflammatory reactions. In addition, CPhGs regulated the abundance of the [Eubacterium]_coprostanoligenes_group, Oscillospiraceae_UCG-005, etc. to modulate the gut microbiota. CPhGs significantly upregulated the content of vitamin B6 and tyrosyl-tryptophan and downregulated histamine, L-methionine, etc. In summary, the therapeutic efficacy of CPhGs on DN rats may be achieved by modulating the gut microbiota and cecal metabolites to restore the metabolic disorders of vitamin B6, histidine, etc.
INTRODUCTION
There is an increase in diabetes mellitus (DM) cases globally; by 2030, the prevalence is projected to be 10.9% of the adult population. 1 A significant complication of DM is diabetic nephropathy (DN), which ranks as the primary cause of chronic kidney disease. 2,3 Currently, the management of DN focuses on controlling blood pressure and blood glucose levels while avoiding nephrotoxic drugs. 4,5 Dapagliflozin is a new hypoglycemic agent with independent cardiovascular and renal benefits in addition to its hypoglycemic effects, and is currently the preferred treatment for DN. 6,7 Nevertheless, owing to the complex pathogenic mechanisms of DN and the absence of specific targets for intervention, current treatment approaches have limitations, necessitating the exploration of new strategies. 8
Notably, Chinese medicine shows promise as a primary or alternative therapy for DN treatment because of its multitarget effects. Research studies increasingly focus on identifying bioactive ingredients and elucidating the molecular mechanisms underlying the renoprotective properties of Chinese medicine. 9 Traditional Chinese medicine commonly uses herbal extracts such as Salvia miltiorrhiza, 10 Tripterygium wilfordii, 11 Flos Abelmoschi, 12 and Radix Astragali 13 for DN treatment. Moreover, triptolide has demonstrated the ability to reduce diabetic kidney fibrosis by promoting the restoration of autophagy. 14 Astragalus membranaceus injection has been shown to have renal protective effects and to improve systemic status, as demonstrated in a meta-analysis of its clinical efficacy in DN patients. 15 Baicalin, which prevents Notch 1-Snail axis-induced podocyte epithelial–mesenchymal transition, has shown promise in reducing the proteinuria and focal segmental glomerulosclerosis induced by adriamycin. 16
The gut microbiota is a diverse community of numerous bacteria, archaea, fungi, protozoa, and viruses. Within this ecosystem, there are more than 1000 bacteria that make up the intestinal flora. 17 An imbalance of the intestinal flora, known as dysbiosis, is associated with both structural changes and functional abnormalities. Structural changes often result in significant reductions in beneficial bacteria and the proliferation of opportunistic pathogens. This leads to functional changes in the intestinal flora, impacting substance and energy metabolism as well as the host’s immune response. These disruptions in the gut microbiota contribute to an elevated susceptibility to disease, thereby perpetuating a detrimental cycle. 18 The dysregulation of intestinal flora is an essential factor in metabolic disorders, including diabetes, obesity, and atherosclerotic coronary heart disease. 19,20 The “gut-kidney axis” theory provides a new insight into intestinal flora etiology. 21,22 Numerous studies have reported a correlation between an imbalance in the intestinal flora and occurrence and progression of renal injury, such as kidney stones 23 and lupus nephritis. 24 The gut microbiota acts as a messenger between the kidney and external environment, playing a significant role in DN. 25 –27 Changes in the gut or kidney can impact the intestinal mucosa, microbiota, and immune inflammation. 28 In late-stage DN, kidney function is severely impaired, causing an increase in nitrogen waste released through the intestines. This can lead to an overgrowth of harmful bacteria in the gut, worsening the imbalance of intestinal flora. In addition, this imbalance and a weakened gut barrier can trigger inflammation throughout the body, ultimately worsening kidney injury. 29,30
Cistanche tubulosa (Schenk) Wight belongs to the Orobanchaceae family and is commonly referred to as “desert ginseng.” It is widely used in traditional TCM for its therapeutic effects on kidney insufficiency, senile constipation, etc. 31,32 The chemical composition of Cistanche tubulosa consists primarily of phenylethanol glycosides (CPhGs), volatile oils, iridoids, lignans, aldithols, etc. 33 Previous studies have demonstrated that CPhGs exhibit pharmacological properties, such as antidiabetic, 34 renal protection, 35 antiliver cancer, 36 and regulatory effects on intestinal microorganisms. 32 Animal studies have also shown that CPhGs can alleviate kidney injury in db/db mice. 35 However, the specific action mechanisms of CPhGs on DN are not fully understood.
Based on the aforementioned research findings, we hypothesized that CPhGs could ameliorate kidney injury in DN by modulating intestinal flora and metabolites. In this study, a DN rat model was established by streptozotocin (STZ) injection combined with a high-fat diet (HFD). The cecal contents DN rats underwent analysis through 16S rRNA gene sequencing and broad-spectrum metabolite profiling to investigate the mechanism of CPhGs in ameliorating DN, which could serve as a valuable reference for future clinical research.
MATERIALS AND METHODS
Preparation of CPhGs and experimental animals
CPhGs produced by Hetian Dichen Biotech Co., Ltd. contain over 80% total substance, including echinacoside at 44.5% and verbascoside at 16.1%. Seventy male SD rats, aged 8 weeks and weighing 240.0 ± 20.0 g, were used in the study. The study conformed to all relevant international and national animal care and use regulations. An Experimental Animal Ethics Committee at Xinjiang Medical University approved all animal care and use procedures for this study (IACUC-20220127-19).
Animal modeling and grouping
Each rat was given their respective diets after a week of adaptable feeding. Ten rats were assigned to the Control group and fed a conventional diet, whereas the remaining rats were given an HFD (cat. no. B1135DM containing 35.5% fat, 2.6% protein, and 43.9% carbohydrates, Beijing Boai Port Biotechnology Co., Ltd.). After 6 weeks, STZ (30 mg·kg−1) was intraperitoneally injected at a concentration of 0.1 mmol·L−1 in sodium citrate buffer. If fasting blood glucose (FBG) was ≥ 16.7 mmol·L−1 after 72 h, the DM model was considered successfully established. The rats were subsequently fed an HFD for an additional 2 weeks. A successful animal model of DN was defined as a positive urine glucose level and a urine albumin excretion rate (UAER) ≥ 20 mg. Unfortunately, three rats died during the modeling process and seven rats failed to successfully establish the model. Fifty rats that exhibited successful development of DN were randomly allocated into five groups: the Model group, Dapagliflozin group, as well as low-dose (CPhGs-L), middle-dose (CPhGs-M), and high-dose (CPhGs-H) CPhGs groups (n = 10). Groups of rats in Control and Model groups received 1 mL of distilled water per 200 g body weight via gavage. Dapagliflozin was administered at 1 mg·kg−1 to the dapagliflozin group (cat. no. NB2248; AstraZeneca Pharmaceuticals Co., Ltd.), whereas the CPhGs groups were given CPhGs at 125, 250, or 500 mg/kg, respectively, via gavage once daily for 6 weeks. The concentration of CPhGs was determined based on preliminary experiments. Body weight was measured weekly, and FBG was assessed every 2 weeks. Subsequently, the rats were euthanized, and kidney, blood, serum, cecal contents, and 24-hour urine samples were collected.
Measurement of biochemical indicators
Several tests were conducted to determine the levels of creatinine (Cr), blood urea nitrogen (BUN), urine protein, uric acid (UA), and glycosylated hemoglobin (HbA1c) in each group of rats following the manufacturer’s recommendations (cat. nos. C011-2-1, C013-2-1, C035-2-1, C012-2-1, A056-1-1; Nanjing Jiancheng Biological Engineering Institute). Activation of superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), and malondialdehyde (MDA) levels (cat. nos. RXWB0477, RXWB0100, RXWB0005; Quanzhou Ruixin Biotechnology Co., Ltd.) in homogenized kidney tissue were assessed according to the instructions provided with each kit.
ELISA
Enzyme Linked immunosorbent Assay (ELISA) was performed to determine the levels of insulin, cystatin C (Cys C), interleukin-1β (IL-1β), fasting insulin (FINS), interleukin-6 (IL-6), tumor necrosis factor alpha (TNF-α), and β2-microglobulin (β2-MG) in the serum (cat. nos. JL10692, JL20884, JL20896, JL13202, JL11506, JL15772; Shanghai Jianglai Biotechnology Co., Ltd.) and homeostatic model assessment of insulin resistance (HOMA-IR) scores were calculated for each group of rats after 6 weeks of CPhG intervention.
Histology and immunohistochemical staining
The rats’ kidneys were surgically removed, preserved in 10% formalin, and embedded in paraffin following a 6-week CPhGs intervention. After that, the kidney tissue samples were divided into 4-μm pieces and xylene was used to deparaffinize them. The sections were stained with PAS (cat. nos. DC0032, DG0005; Beijing Regen Biotechnology Co., Ltd.), Masson’s trichrome, or conventional hematoxylin and eosin (HE). The kidney levels of α-SMA and collagen I protein expression were evaluated using immunohistochemistry. Goat serum for closure (cat. no. ZLI-9022; Nakasugi Golden Bridge, Inc.) was used to block the sections after the sample’s endogenous peroxidase activity was quenched for 60 min at 25°C. Primary antibodies against α-SMA (1:3,000; cat. no. 14395-1-AP; Proteintech Group, Inc.) and collagen I (1:100; cat. no. AF7001; Affinity, Inc.) were incubated on sections overnight at 4°C. A light optical microscope was used to take the pictures. Pathological tissue slices were analyzed using ImageJ 1.8.0 and integrated optical density (IOD). IOD/sum area × 100% was the formula used to compute the positive area (%).
S rRNA sequencing
After 6 weeks of CPhG intervention, the cecal contents were collected. The cetyltrimethylammonium bromide method was applied for obtaining the whole DNA from the cecal contents of rats with DN. The 16S rRNA gene was amplified using primers 341F and 805R in a polymerase chain reaction (PCR) mixture with template DNA. The PCR products were purified using AMPure XT beads (cat. no. A63882; Beckman Coulter, Inc.) and quantified using Qubit (ThermoFisher Scientific, Inc.) Detection was performed using a 2% agarose gel. Purified PCR products were evaluated using an Agilent 2100 Bioanalyzer (ThermoFisher Scientific, Inc.) and an Illumina library quantification kit (cat. no. NQ101; Novizan Biotechnology Co., Ltd.). The qualifying library concentration was greater than 2 nmol/L. Each qualifying sequencing library (with no repeated index sequence) was diluted and combined at the appropriate ratio for sequencing. After denaturation to a single strand with NaOH, 500 cycles of two-end sequencing of 2250 bp were performed.
Cecal content metabolite measurement
Initially, 50 mg of cecal contents from each rat was extracted using a mixture of methanol, acetonitrile, and water. The extract was then frozen at −40°C for 1 h and centrifuged at 21,367 g for 15 min at 4°C. The resulting supernatant was used as the test sample. All the samples were prepared in the same manner and evaluated repeatedly throughout the analytical procedure to ensure the accuracy and stability of each measurement.
Statistical analysis
The data were analyzed using SPSS version 25.0 and presented as mean values ± standard deviations (SD). One-way analysis of variance (ANOVA) was used to compare groups. principal component analysis (PCA) and orthogonal partial least squares discriminant analysis (OPLS-DA) were utilized to discern variations in metabolic profiles across different groups using SIMCA software. The correlation between the components was measured using Spearman’s correlation coefficient, with statistically significant findings accepted at P-values < 0.05 and < 0.01.
RESULTS
CPhGs on biochemical indicators, pathological changes, and in DN rats
In comparison to the Control group, the DN rats had significantly decreased body weights after the last dose (P < 0.01), which is typical of DM. The body weights of DN rats in the Dapagliflozin, CPhGs-H, and CPhGs-M dosage groups were notably higher (P < 0.05, Fig. 1A) than those in the Model group. After 2 weeks of injecting STZ, the FBG levels in all groups rose above 20.0 mmol/L. Following a 6-week treatment, the levels of FBG showed a notable decrease in the Dapagliflozin, CPhGs-H, CPhGs-M, and CPhGs-L dosage categories compared with the Model group (P < 0.05, Fig. 1B). The Model group exhibited significantly elevated levels of HbA1c, FINS, and HOMA-IR compared with the Control group (P < 0.01). Dapagliflozin, CPhGs-H, or CPhGs-M intervention led to significantly lower levels of HbA1c, FINS, and HOMA-IR in DN rats than in Model rats (P < 0.01, Fig. 1C–E).
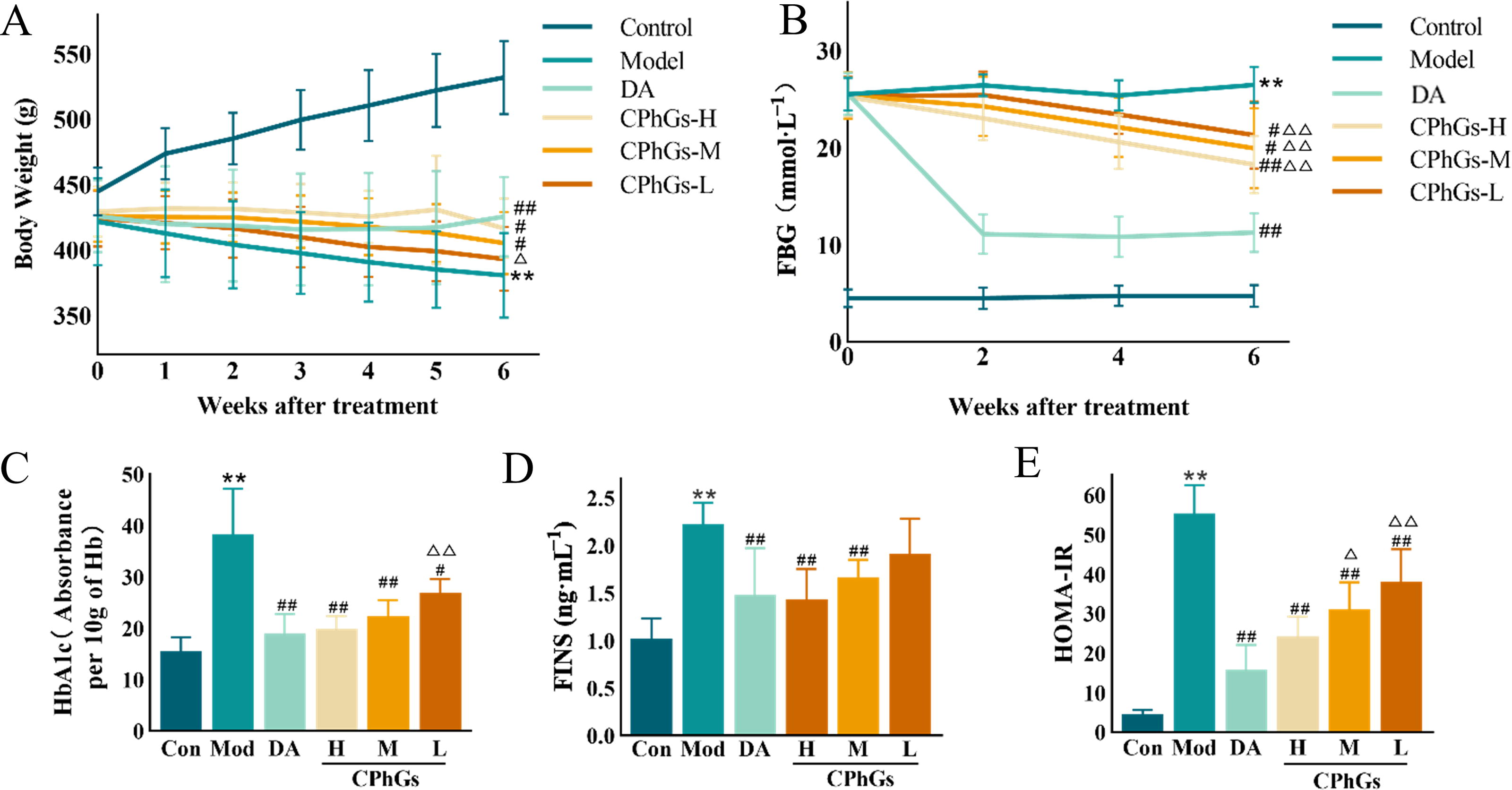
CPhG treatment ameliorated the body weight change and glucose metabolism of DN rats.
Renal function tests showed higher levels of UAER, Cr, BUN, Cys C, and β2-MG (P < 0.01) in the Model group compared with the Control group. Intervention with Dapagliflozin, CPhGs-H, or CPhGs-M resulted in significantly lower UAER, Cr, BUN, Cys C, and β2-MG (P < 0.05) levels in DN rats than in Model rats. The CPhG-L dose resulted in significantly lower UAER, Cys C, and β2-MG (P < 0.01, Fig. 2A–F) levels in DN rats than in Model rats.
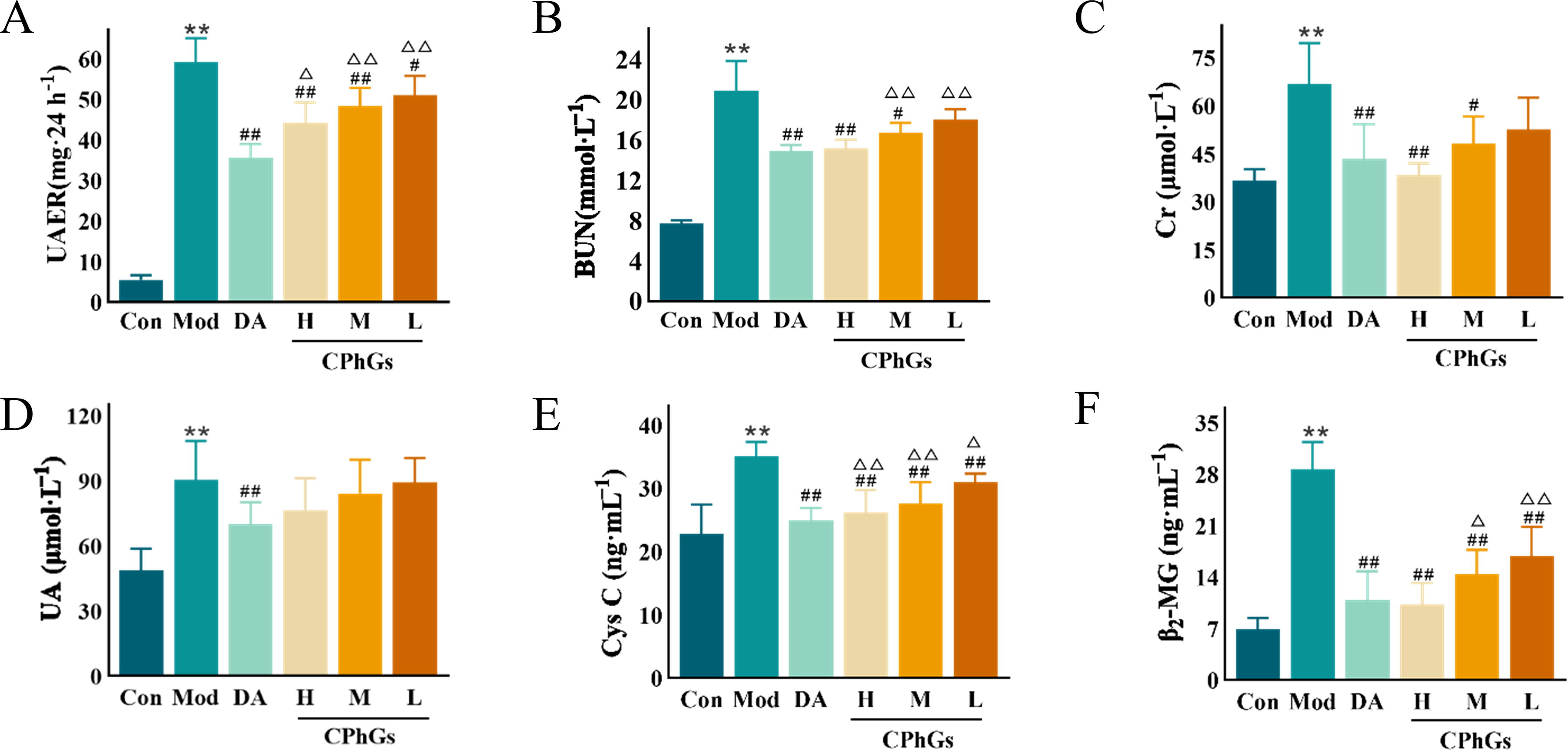
CPhG treatment ameliorated the renal function of DN rats.
The Control group rats showed intact glomeruli and tubules without any signs of inflammatory cell invasion or mesangial stromal hyperplasia, as indicated by the HE staining results. Conversely, the Model group showed mesangial hyperplasia, glomerular enlargement, slight thickening of the basement membrane, and localized degeneration and atrophy of renal tubules. Lesions were reduced in the Dapagliflozin group, as well as in the CPhGs-H and CPhGs-M dose groups, compared with the Model group (Fig. 3A). Masson staining revealed that the groups administered with Dapagliflozin, CPhGs-H, and CPhGs-M doses exhibited decreased collagen deposition in both the glomerular and tubulointerstitial regions as compared with the Model group (P < 0.05, Fig. 3A and B). PAS staining showed a notable increase in glomerular glycogen accumulation in the Model group compared with the Control group (P < 0.01), whereas the Dapagliflozin, CPhGs-H, and CPhGs-M dosage groups exhibited reduced glycogen accumulation compared with the Model group (P < 0.05, Fig. 3A and C).
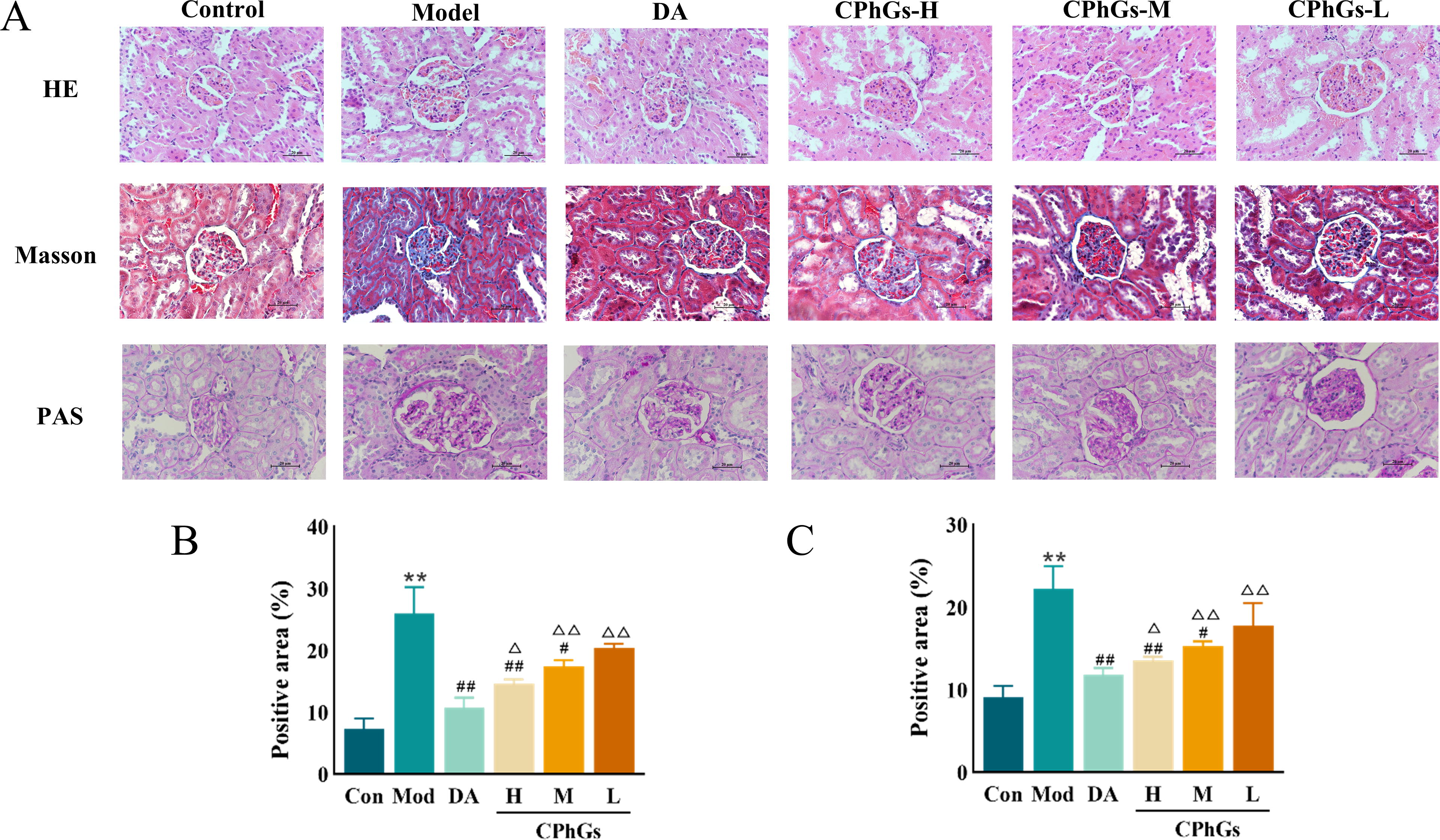
CPhG treatment ameliorated the pathological changes in kidney tissue of DN rats.
CPhGs suppress oxidative stress in the kidneys of DN rats
Compared with Model rats, renal tissue homogenates from animals treated with dapagliflozin, CPhGs-H, or CPhGs-M exhibited greater SOD and GSH-Px activities and lower MDA levels (P < 0.01, Fig. 4A–C).
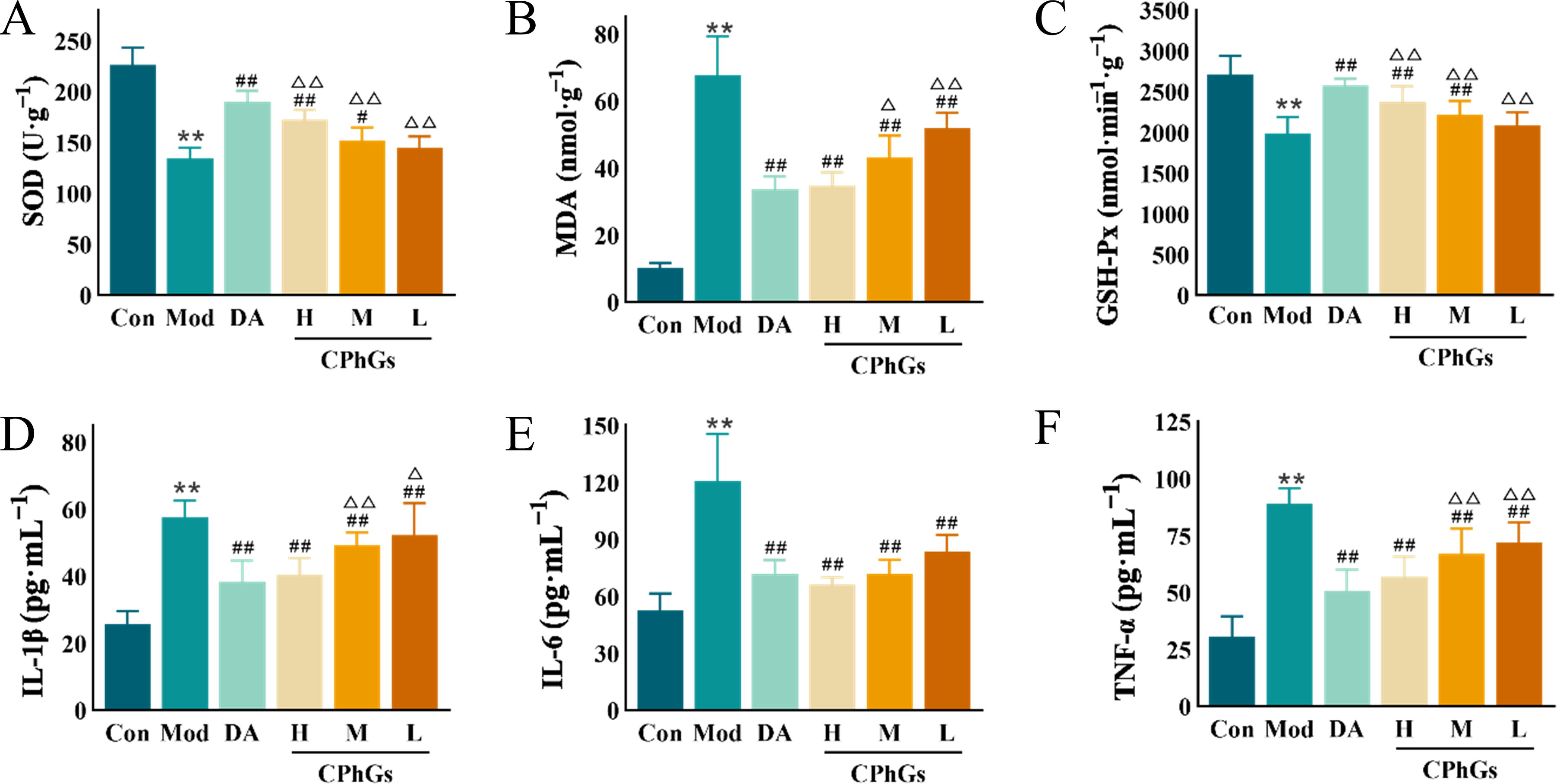
CPhGs treatment ameliorated the levels of oxidative stress and inflammatory factors of DN rats.
CPhGs decrease inflammatory factors in the serum of DN rats
The serum levels of the proinflammatory factors IL-1β, IL-6, and TNF-α were significantly greater (P < 0.01) in the Model group than in the Control group. Treatment with dapagliflozin, CPhGs-H, CPhGs-M, or CPhGs-L resulted in significantly lower levels of IL-6, IL-1β, and TNF-α in the sera of DN rats (P < 0.01, Fig. 4D–F).
CPhGs lower protein expression of α-SMA and collagen I in the kidneys of DN rats
The results demonstrated that the α-SMA and collagen I expression levels were markedly decreased in the CPhG-treated groups compared with the Model group (P < 0.01, Fig. 5A–C). The above results demonstrated the therapeutic benefits of CPhGs in DN rats, with the greatest effectiveness observed at a large dosage. Thus, the CPhGs-H dose group was chosen for further intestinal flora and metabolomics research.

CPhG treatment decreased the expression of α-SMA and collagen I proteins in kidney tissue of DN rats.
CPhGs modulate the gut microbiota in DN rats
According to the results of the microbiome analysis, an increase in the number of species in the environment did not significantly increase the sample dilution curve (Fig. 6A). This finding suggested that the sample sequence was adequate and that the extracted samples contained the majority of the microbial species. The Shannon and Simpson indices were significantly lower in the Model group compared with the Control group (P < 0.05, Fig. 6B and C). PCoA and clustering analysis indicated that samples from the Model group were different from the Control group, but the CPhG-H dosage group was similar to the Control group. (Fig. 6D and E). Firmicutes and Bacteroidetes were the main phyla in each group (Fig. 6F), with Firmicutes significantly lower in the Model group compared with the Control group but significantly higher in the CPhGs-H dosage group compared with the Model group (P < 0.01). Conversely, Bacteroidetes showed the opposite trend (Fig. 6G). At the genus level, the Model group had a greater relative abundance of Paludicola and a lower relative abundance of [Eubacterium]_coprostanoligenes_group, Oscillospiraceae_UCG-005, Clostridia_UCG-014, Christensenellaceae_R-7_group, and Monoglobus (P < 0.05) than the Control group. In addition, the Model group had lower abundances of [Eubacterium]_coprostanoligenes_group, Oscillospiraceae_UCG-005, Clostridia_UCG-014, Christensenellaceae_R-7_group, and Monoglobus and a greater abundance of Paludicola (P < 0.05) than did the CPhGs-H dosage group (Fig. 6H). These findings indicate that the overall structure and composition of the intestinal microbiota were significantly altered in the Model group and that a high dosage of CPhGs successfully reversed these alterations. The predicted functional alterations in the intestinal microbiota of DN rats were analyzed using PICRUSt2. The common differential metabolic pathways were the metabolisms of cyanoamino acids, D-glutamine and D-glutamate, propanoate, starch, and sucrose. Specifically, compared with the Model group, the metabolism of cyanoamino acids, starch and sucrose was downregulated, and the metabolism of D-glutamine and D-glutamate and propionate was upregulated in the CPhGs-H dose group (Fig. 6I and J).
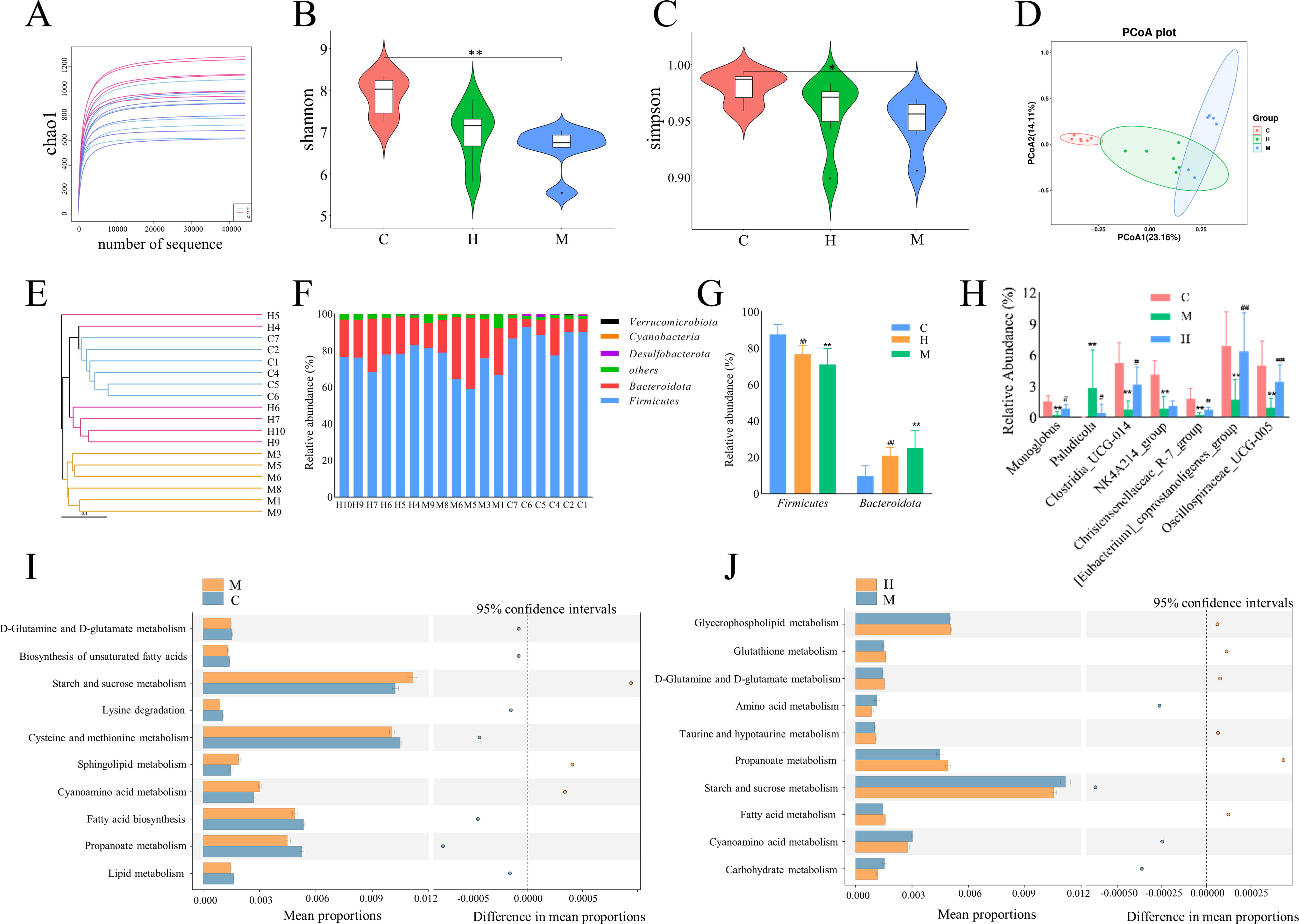
CPhG treatment affected the diversity and differences of gut microbiota in DN rats.
Effects of CPhGs on cecal metabolites in DN rats
Total ion current maps of the metabolites were obtained through liquid chromatography-tandem mass spectrometry analysis of the cecal contents from each group of animal samples. The datasets were subjected to PCA utilizing a linear combination of metabolite variables with specific weights. The PCA plots clearly revealed differences among the Control group, Model group, and CPhG-H dose group (Fig. 7A). The OPLS-DA model demonstrated the reliability and accuracy of the model for making predictions (Fig. 7B–E). he screening criteria for differentially abundant metabolites were a variable importance projection > 1.0 and a significance threshold of P < 0.05. In the Model group, 307 metabolites showed differential abundances compared with the Control group, with 219 downregulated and 88 upregulated metabolites (Fig. 7F). In the CPhGs-H dose group, 154 metabolites differed from the Model group, with 25 downregulated and 129 upregulated metabolites (Fig. 7G). CPhGs significantly upregulated the content of vitamin B6, tyrosyl-tryptophan, and downregulated histamine, L-methionine, etc. (Table 1). Furthermore, the enrichment results of metabolic pathways revealed that the CPhGs mainly influenced metabolic pathways such as vitamin B6, histidine, tyrosine, cysteine, and methionine metabolism, as well as steroid hormone synthesis in DN rats (Fig. 7H).
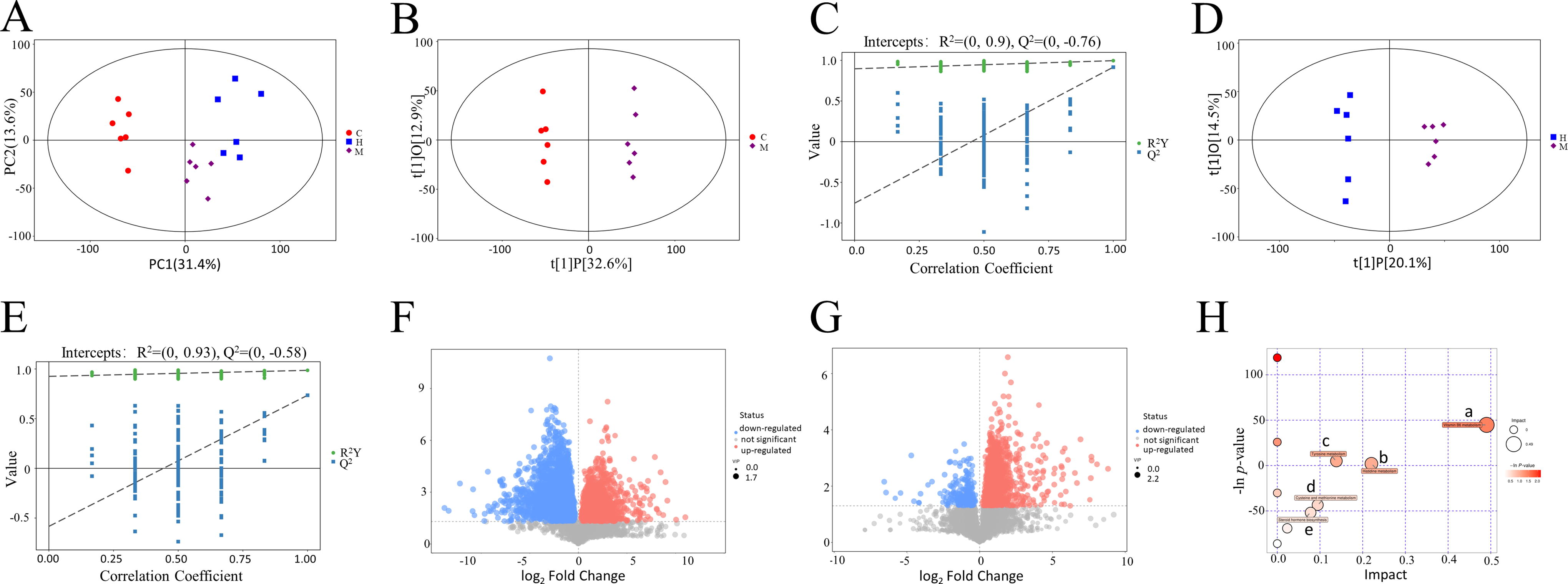
CPhG treatment modulated the cecal metabolites of DN rats.
The Differential Metabolites in Cecal Contents After CPhGs Treatment
Model and CPhG high-dose groups (n = 6/group). Compared with the Model group.
P < 0.05, ** P < 0.01.
↑, content increased; ↓, content decreased; FC, fold change.
Relationships between biochemical indices, differentially abundant metabolites, and differential gut microbiota in DN rats
To investigate the effect of CPhGs on the intestinal flora composition and its connection with DN-related characteristics, we conducted an analysis using Spearman’s correlation. The results showed that the [Eubacterium]_coprostanoligenes_group, Oscillospiraceae UCG-005, Clostridia UCG-014, Christensenellaceae_R-7_group, Monoglobus, and NK4A214 groups exhibited a negative relationship with proinflammatory cytokines and oxidative stress, whereas Paludicola showed a positive relationship (Fig. 8A). In addition, [Eubacterium]_coprostanoligenes_group, Oscillospiraceae_UCG-005, Clostridia_UCG-014, Christensenellaceae_R-7_group, Monoglobus, the NK4A214 group, and Paludicola showed strong correlations with most of the differentially abundant metabolites (Fig. 8B).

Spearman’s correlation analysis of physiological indicators, oxidative stress and inflammatory factors, differential metabolites of cecal contents, and gut microbiota.
DISCUSSION
IR significantly contributes to DN, and our findings reveal that the DN rats had notably higher levels of FBG and HbA1c compared with the Control group. Compared with the Control group, the Cr, BUN, UAER, Cys C, and β2-MG were increased in the Model group, among which Cys C and β2-MG are increasingly recognized as valuable indices of kidney function. 37,38 The results of biochemical indicators and pathology demonstrated that CPhG intervention effectively enhanced kidney function, and decreased renal tissue damage and the deposition of glycogen and collagen in DN rats. According to the recommendations of the China Diabetes Kidney Disease Prevention and Treatment Guide, patients with DN should prioritize drugs that have been proven to be beneficial for the kidneys when selecting hypoglycemic drugs, so we selected dapagliflozin as the positive control drug. We discovered no statistically significant difference in improving renal function or reducing pathological damage compared to the high dose of CPhGs. However, the ability of the CPhG-treated rats to regulate blood glucose levels was slightly lower than that of the rats in the positive drug group. These slight variations indicate that CPhGs have the potential to be a long-term treatment option for DN, preventing the toxic side effects that long-term use of chemicals.
In addition, our findings suggest that intervention with CPhGs assists in reducing the oxidative stress response of renal tissue in DN rats. Oxidative stress is considered the primary cause of DN onset. 39 DN development enhances the production of reactive oxygen species (ROS) through pathways such as hexosamine, polyol, and advanced glycation end products (AGEs), thereby intensifying oxidative stress responses. 40,41 Hyperglycemia of DN triggers chronic inflammation in the body, ultimately resulting in the production of substantial amounts of proinflammatory substances in renal tissue. 42 In accordance with our findings, DN rats’ serum levels of markers of inflammation decreased as a result of CPhGs therapy, indicating its anti-inflammatory effect.
We investigated the impact of CPhGs on the intestinal microbiota composition in DN rats using 16S rRNA sequencing. The results, as illustrated in the PCoA figure, demonstrated that CPhGs could alter the alpha and beta diversities of the gut microbiota, 32 indicating that the intestinal microbiota of DN rats became more similar to the floral structure of the Control group rats after CPhG administration. 43 Subsequent microbiome investigation showed that the predominant species in the gut microbiota of all the groups were Firmicutes and Bacteroides at the phylum level. Notably, the CPhGs-H dose group exhibited a greater relative abundance of Firmicutes and a lower relative abundance of Bacteroides than did the Model group. These alterations in the microbiota have important implications for the inflammatory response, metabolic issues, and other clinical conditions associated with DN. 44
At the genus level, our findings showed that CPhGs increased the abundance of Oscillospiraceae_UCG-005, [Eubacterium]_coprostanoligenes_group, Clostridia_UCG-014, Monoglobus, and Christensenellaceae_R-7_group, while decreasing the abundance of Paludicola in DN model rats. Probiotic bacteria in the [Eubacterium]_coprostanoligenes_group have been shown to ameliorate oxidative stress and deregulation of the intestinal ecology 45 in type 2 DM rats. Moreover, this group is believed to alleviate hyperlipidemia by producing butyrate, which regulates lipid metabolism. 46 Oscillospiraceae UCG-005 has been found to be closely related to the serum UA level in patients with diabetes. 47 The abundance of Clostridia UCG-014 in the gut was increased by CPhG treatment, which is consistent with the effects of Asiatic acid 48 and Artemisia argyi essential oil 49 in improving metabolic processes and lipid metabolism. Recent studies have also suggested that fermented carrot juice enhances insulin sensitivity, antioxidant capacity, pancreatic and kidney morphology, and blood sugar levels in diabetic rats, in addition to increasing short-chain fatty acids (SCFAs) and the abundance of the Christensenellaceae_R–7_group in the cecum. 50 These studies indicate that promoting the growth of SCFA-producing microbiota can significantly reduce the risk of developing various metabolic disorders, such as diabetes and obesity. 51
Furthermore, we found that Paludicola was the only species with a significantly different relative abundance among the Control, Model, and CPhGs-H dose groups. Moreover, the NRS-2002 score was positively correlated with certain physiological indicators, oxidative stress indicators, and proinflammatory cytokines. However, the exact role of Paludicola in metabolic health remains unclear. 52 Future studies utilizing fecal transplantation or gut microbiota should be conducted to further investigate the potential of CPhGs in treating DN by modulating the gut microbiota. In addition, combining the gut microbiota results with the metabolomics findings revealed that vitamin B6, histidine, cysteine and methionine, and tyrosine metabolism were key metabolic pathways affected by CPhCG treatment. These findings suggested that CPhGs may exert their therapeutic effects on DN by modulating these metabolic pathways.
Vitamin B6 is involved in approximately 150 reactions that regulate glucose, lipid, amino acid, DNA, and neurotransmitter metabolism in organisms. 53 In addition, it acts as an antioxidant, preventing the production of ROS and AGEs. Research has shown that the accumulation of AGEs leads to inflammation and the impairment of blood vessels, resulting in vascular problems. 54 AGEs promote the generation of ROS, which in turn compromises DNA integrity. This creates a harmful cycle where increased ROS production encourages the synthesis of additional AGEs. 55 Furthermore, studies have demonstrated that oral pyridoxine treatment can reduce kidney damage and dysfunction caused by excess fat and fructose in mice. 56 In addition, vitamin B6 supplementation may slow the progression of DN. 57 In our study, we observed a reduction in pyridoxal levels in DN model rats, which significantly increased after treatment with CPhGs. Spearman’s analysis revealed a positive correlation between the pyridoxal concentration and abundance of the Oscillospiraceae_UCG-005, [Eubacterium]_coprostanoligenes_group, Clostridia_UCG-014, Monoglobus, Christensenellaceae_R-7_group, and NK4A214 groups. Conversely, there was a negative relationship between pyridoxal and Paludicola. Based on these findings, we hypothesize that the changes in the abundances of Oscillospiraceae_UCG-005, [Eubacterium]_coprostanoligenes_group, Clostridia_UCG-014, Christensenellaceae_R-7_group, Monoglobus, NK4A214_group, and Paludicola may be linked to the impact of CPhGs on vitamin B6 metabolism.
Histamine is an inflammatory mediator synthesized by the body from histidine through the action of decarboxylase. 58 It plays a regulatory role in various processes by generating cytokines, chemokines, and growth factors. Histamine exacerbates allergic inflammation by binding to H4 receptors and contributes to angiogenesis and fibrosis through H2 receptors. 59 In addition, accumulating evidence suggests that in addition to its impact on glomerular microcirculation hemodynamics, histamine may directly contribute to renal pathology. For example, it has been demonstrated that histamine increases tubular apoptosis induced by fasting. Moreover, the increased presence of mast cells in the tubulointerstitium and their degranulation in the progression of DN point to a direct involvement of histamine in the development of renal fibrosis and inflammation. 60 An important biomarker distinguishing DN patients from healthy controls is histamine: patients with DN exhibit significantly higher plasma histamine levels, which decrease with decreasing kidney function. 61 Similarly, our study revealed elevated histamine levels in DN rats, which decreased following intervention with CPhGs. Research has shown that alterations in the gut microbiota composition can lead to increased histamine production, resulting in intestinal barrier disruption and a subsequent inflammatory response. 62 The current study’s Spearman’s analysis indicated a positive correlation of Paludicola and a negative correlation of Oscillospiraceae_UCG-005, Clostridia_UCG-014, [Eubacterium]_coprostanoligenes_group, Christensenellaceae_R-7_group, Monoglobus, and the NK4A214_group with histamine levels. Accordingly, we hypothesize that the effects of CPhGs on histamine levels may be associated with changes in the abundances of Paludicola, Oscillospiraceae_UCG-005, Clostridia_UCG-014, [Eubacterium]_coprostanoligenes_group, Christensenellaceae_R-7_group, Monoglobus, and the NK4A214_group, leading to an improvement in DN in rats.
Metabolism of methionine and tyrosine is linked to several illnesses, including IR, fatty liver, and obesity. 63,64 Our study showed that CPhGs significantly reduced methionine content and increased tyrosine methylation levels in DN rats. This suggests that metabolisms of methionine and tyrosine are expected to be potential intervention targets for treating DN.
CONCLUSIONS
In conclusion, CPhGs demonstrated a range of advantageous outcomes in DN, encompassing the enhancement of renal function and alleviation of hyperglycemia, IR, pathological changes, oxidative stress, and inflammatory reactions. The improvement effect of CPhGs on DN rats may be achieved by improving the structure of intestinal flora and restoring the metabolic disorders of vitamin B6, histidine, etc. However, further clinical research is necessary to explore the effects of CPhGs on gut microorganisms in individuals with DN.
Footnotes
AUTHORS’ CONTRIBUTIONS
C.M.: Conceptualization, Data curation, Writing—original draft. B.J.: Conceptualization, Data curation, Writing—original draft. J.L.: Investigation, Data curation. L.W.: Data curation, Funding acquisition. Y.Z.: Validation. J.Y.: Formal analysis, Funding acquisition, Supervision, Writing—review and editing. J.H.: Funding acquisition, Project administration, Supervision, Writing—review and editing.
AUTHOR DISCLOSURE STATEMENT
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
ETHICAL GUIDELINES
All animal care and use procedures for this study were approved by the Experimental Animal Ethics Committee of Xinjiang Medical University (IACUC-20220127-19).
DATA AVAILABILITY STATEMENTS
The data that support the findings of this study are available from the corresponding author upon request.
FUNDING INFORMATION
This study was supported by