
Abstract
Muscle atrophy, characterized by diminished muscle mass and impaired function, poses a substantial global health concern. Boesenbergia pandurata (Roxb.) Schltr., commonly known as fingerroot, possesses a variety of advantageous activities, including anti-inflammatory, antioxidant, antibacterial, and anticancer effects. However, there are currently no preclinical studies available that explore the potential of B. pandurata extract (BPE) to mitigate muscle atrophy. In this study, we aimed to explore the protective effects of BPE, standardized to panduratin A content, against muscle atrophy and its underlying molecular mechanisms in a dexamethasone-induced muscle atrophy mouse model. Compared with the dexamethasone group, BPE significantly restored muscle mass, muscle volume, muscle fiber cross-sectional area, grip strength, and exercise endurance. Additionally, BPE suppressed inflammatory responses by downregulating the expressions of nuclear factor kappa B and inflammatory cytokines while also enhancing antioxidant effects by increasing the expressions of antioxidant enzymes. Moreover, BPE promoted protein synthesis and muscle differentiation by stimulating the phosphatidylinositol 3-kinase/Akt/mammalian target of rapamycin signaling pathway. Furthermore, it suppressed myostatin expression and inhibited the expressions of E3 ubiquitin ligases by preventing the nuclear translocation of forkhead box O3a, thereby alleviating proteolysis. Overall, BPE effectively regulates unbalanced protein metabolism, suggesting its potential as a functional food ingredient for preventing muscle wasting diseases.
INTRODUCTION
Muscle atrophy can result from various factors, including cancer, aging, and drug side effects. It leads to diminished muscle function, increasing mortality and morbidity risks while lowering patients’ quality of life. 1 Muscle atrophy arises from a disruption in the balance between muscle protein degradation and synthesis. Therefore, efforts to regulate the factors involved in protein anabolic and catabolic pathways are essential to restore balanced protein metabolism. Proteolysis typically occurs via the ubiquitin-proteasome system (UPS). Heightened UPS activity promotes the expressions of E3 ubiquitin ligases, such as muscle RING-finger protein-1 (MuRF1) and atrogin-1, leading to muscle proteolysis. 2 Myostatin (Mstn) and forkhead box O3a (FoxO3a) play crucial roles in this process by regulating these E3 ubiquitin ligases. 3,4 Inflammation and oxidative stress also accelerate protein degradation. Excessive reactive oxygen species (ROS) induce oxidative stress and activate the nuclear factor kappa B (NF-κB) signaling pathway, triggering inflammation. ROS and activated NF-κB further stimulate MuRF1 and atrogin-1, resulting in accelerated protein degradation. 5
The phosphatidylinositol 3-kinase (PI3K)/Akt signaling pathway is critical in regulating protein synthesis by stimulating activation of mammalian target of rapamycin (mTOR) and the downstream factors, 70-kDa ribosomal protein S6 kinase (p70S6K), and eukaryotic translation initiation factor 4E-binding protein 1 (4EBP1). Additionally, activation of this pathway promotes myogenesis by enhancing muscle cell differentiation and growth while simultaneously suppressing protein degradation mechanisms. 6 This dual functionality underscores the pathway’s critical importance in preserving muscle mass and function. Understanding these mechanisms and identifying materials that can effectively regulate protein synthesis and degradation pathways may offer a powerful strategy for preventing muscle atrophy.
Glucocorticoids, a class of steroid hormones, are commonly used as anti-inflammatory agents. However, high doses or prolonged use can cause side effects, such as osteoporosis, hypertension, and muscle atrophy. 7 Dexamethasone (DEX), a synthetic glucocorticoid, induces muscle atrophy primarily by enhancing protein catabolic pathways and inhibiting anabolic pathways, making DEX treatment a well-established model in numerous studies. 8,9 Therefore, DEX was employed to induce muscle atrophy in this study.
Boesenbergia pandurata (Roxb.) Schltr., commonly referred to as fingerroot, is an herb in the Zingiberaceae family. 10 The rhizome of B. pandurata is employed as a spice and has traditionally been used to treat various diseases, such as asthma, diarrhea, indigestion, itching, ulcer, leukorrhea, and dysentery. 11 Additionally, B. pandurata has been recognized for its anti-inflammatory, antioxidant, antibacterial, anticancer, antiobesity, and antiaging properties, as demonstrated in numerous pharmacological studies. 12 Notably, B. pandurata contains panduratin A and a chalcone derivative, as a key bioactive compound. Panduratin A has been documented to inhibit muscle loss by stimulating the PI3K/Akt/mTOR signaling pathway in tumor necrosis factor α (TNF-α)-treated L6 skeletal muscle cells. 13 Furthermore, B. pandurata extract (BPE), standardized to panduratin A content, has been shown to improve exercise performance by promoting mitochondrial biogenesis in both obese and normal mice. 14 However, there are currently no preclinical studies exploring the potential of BPE to mitigate muscle wasting. The purpose of this current study was to demonstrate whether BPE inhibits the development of muscle atrophy by elucidating the underlying molecular mechanisms in DEX-induced muscle atrophy mice.
MATERIALS AND METHODS
Plant materials
Dried rhizomes of B. pandurata were extracted using 95% ethanol at 40°C for 8 h and then filtered. The solvent in filtrates was completely removed using a rotary evaporator to obtain BPE (12%, w/w). The standardization of BPE was carried out with panduratin A. Quantitative analysis of panduratin A in the standardized BPE was performed using a high-performance liquid chromatography system (YL Instrument Co. Ltd., Anyang, Korea), and the content of panduratin A in the BPE was 8% (w/w) (Supplementary Fig. S1). The dried extract was stored at −20°C for further experiments.
Animal experiment
Six-week-old male C57BL/6 mice were acquired from Samtako (Osan, Korea) and housed under a constant environment (a temperature of 23 ± 2°C, a humidity of 55 ± 5%, and a 12 h light/dark cycle) at the Yonsei Laboratory Animal Research Center (Seoul, Korea). All mice had free access to tap water and a standard diet. After 10 days of acclimation, the mice were randomly distributed into four distinct groups (n = 10) as follows: (I) CON (control group); (II) DEX (10 mg/kg/day DEX injection and saline-treated group); (III) DEX+BPE100 (10 mg/kg/day DEX injection and 100 mg/kg/day BPE-treated group); and (IV) DEX+BPE200 (10 mg/kg/day DEX injection and 200 mg/kg/day BPE-treated group). BPE dissolved in saline was administered daily via oral gavage for 17 days until the end of the experiment. The CON and DEX groups were orally treated with saline instead of BPE. One week after the commencement of oral administration, DEX dissolved in saline was administered daily for 10 days through intraperitoneal (i.p.) injection until the end of the experiment. The mice underwent tests to assess physical performance, subjected to muscle volume measurement, and then sacrificed under anesthesia with 1.25% 2,2,2-tribromoethanol (Sigma-Aldrich, St. Louis, MO, USA). After sacrificing the mice, the gastrocnemius (GA), soleus (SOL), tibialis anterior (TA), and extensor digitorum longus (EDL) muscle tissues were isolated and weighed. All muscle tissues were stored at −80°C for further analyses. This study was conducted in accordance with the guidelines established by the Institutional Animal Care and Use Committee of Yonsei University (Permit number: IACUC-A-202309-1729-03).
Micro-computed tomography imaging
Micro-computed tomography (micro-CT) was conducted using an animal positron emission tomography/CT/single-photon emission tomography system (Siemens Medical Solutions, Knoxville, TN, USA). Under anesthesia with 1% isoflurane inhalation, mice were positioned prone on the scanner, and their hindlimb muscles were scanned through a 360° rotation. CT images were analyzed utilizing Inveon Research Workplace software (Siemens Medical Solutions).
Grip strength test
After the administration period, grip strength test was performed using a grip strength meter (Panlab, Barcelona, Spain). The strength meter was reset to 0 g after stabilization in each trial. Each mouse was permitted to grip the pull bar, after which its tail was delicately drawn backward at a constant speed until grip failure. The maximum force was measured at the moment the mouse released its paws from the pull bar. Five successive trials were conducted for each mouse, and all tests were performed by the same person.
Treadmill test
After the administration period, treadmill test was performed using a treadmill machine (Panlab). The test commenced at a velocity of 15 cm/s for 10 min, after which the speed was incrementally raised by 1 cm/s per min until reaching 35 cm/s. The test was terminated when each mouse reached exhaustion, defined as remaining motionless for 10 s despite receiving a 0.2 mA electric shock. The distance and running time were recorded.
Histological analysis
Isolated GA muscle samples were fixed in 10% formalin solution and then embedded in paraffin. These paraffin blocks were stained with hematoxylin and eosin. The stained GA muscle fibers were stained for cross-sectional area (CSA) analysis. The stained fibers were examined under a microscope (CK40; Olympus, Tokyo, Japan), and images were captured using a microscope-mounted camera (eXcope T500; DIXI Science, Daejeon, Korea). The average CSA of the muscle fibers was assessed using ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Western blot analysis
Proteins from GA muscle tissues were extracted using NP-40 lysis buffer (Elpis Biotech, Daejeon, Korea) containing a protease inhibitor cocktail (Sigma-Aldrich). Protein concentrations were quantified using the Bradford protein assay, with bovine serum albumin as the standard reference. Equal amounts of proteins were separated using 7–11% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and then transferred to a nitrocellulose membrane (GE Healthcare, Piscataway, NJ, USA). To avoid nonspecific binding, the membranes were blocked with 5% skim milk (Difco, Detroit, MI, USA) for 1 h at room temperature (25°C) and then incubated overnight at 4°C with primary antibodies against phospho (p)-NF-κB, NF-κB, p-FoxO3a, FoxO3a, p-PI3K, PI3K, p-Akt, Akt, p-mTOR, mTOR, p-p70S6K, p70S6K, p-4EBP1, 4EBP1, and α-tubulin (Cell Signaling Technology, Beverly, MA, USA). Following this, the membranes were incubated for 1 h at room temperature with secondary antibodies. Protein bands were visualized using a G:BOX imaging system (Syngene, Cambridge, UK) with an enhanced chemiluminescence solution (Dyne Bio, Seongnam, Korea). Band intensities were quantified using ImageJ software (National Institutes of Health).
Reverse transcription-polymerase chain reaction analysis
Total RNA from GA muscles was isolated using RNAiso Plus (Takara Bio Inc., Shiga, Japan). The concentration and purity of isolated RNA were measured on a NanoDrop Lite spectrophotometer (Thermo Fisher Scientific Inc., Waltham, MA, USA). cDNA was synthesized using a reverse transcription premix (Elpis Biotech). The cDNA was amplified using a 2720 Thermal Cycler (Thermo Fisher Scientific Inc.) with SafeDry Taq PCR Premix (CellSafe, Yongin, Korea) and gene-specific primers (Bioneer, Daejeon, Korea). The sequences of primers are shown in Table 1. The final polymerase chain reaction (PCR) products were dyed with Dyne LoadingSTAR (Dyne Bio) and separated using electrophoresis on a 1.5% agarose gel. PCR bands were visualized using the G:BOX imaging system (Syngene), and band intensities were quantified using ImageJ software (National Institutes of Health).
Primer Sequences Used in RT-PCR Analysis
CAT, catalase; GPx, glutathione peroxidase; IL-6, interleukin-6; MHC, myosin heavy chain; MuRF1, muscle RING-finger protein-1; MyoG, myogenin; MyoD, myogenic determination protein; RT-PCR, reverse transcription-polymerase chain reaction; SOD, superoxide dismutase catalase; TNF-α, tumor necrosis factor α.
Statistical analysis
The experimental data were analyzed using SPSS Statistics version 27 (IBM Corp., Armonk, NY, USA), and the results are shown as the means ± standard deviations (SD). Significant differences between groups were identified through a one-way analysis of variance followed by Duncan’s multiple range test. Differences were considered statistically significant at P < .05.
RESULTS
BPE restored muscle loss and muscle function in DEX-induced mice
The weights of all muscle types (GA, SOL, TA, and EDL) exhibited a considerable decrease in the DEX group, indicating muscle loss. In contrast, BPE administration dose-dependently restored muscle wasting in DEX-induced mice (Fig. 1A). Additionally, the BPE group showed greater muscle volume relative to the DEX group, with a particularly significant elevation observed following treatment with 200 mg/kg BPE (Fig. 1B, C).

Effects of BPE on muscle weight, muscle volume, and muscle fiber cross-sectional area in mice with DEX-induced muscle atrophy.
The inhibitory effects of BPE on muscle wasting were further measured through histological analysis of muscle tissue. The DEX group demonstrated a marked reduction in CSA of muscle fibers relative to the CON group, whereas administration of BPE improved these atrophic changes in mice experiencing muscle wasting (Fig. 1D, E).
To assess the efficacy of BPE in enhancing muscle function, a grip strength test was conducted. The DEX group showed impaired muscle strength (Fig. 2A, B). The grip strength measurements for the forelimb and fore/hindlimb in the DEX group demonstrated a notable decrease compared to that in the CON group. However, BPE drastically increased both grip strengths relative to those observed in the DEX group (Fig. 2A, B). We also investigated exercise endurance with treadmill test. Compared with that in the CON group, DEX administration led to decreases in both running distance and time. However, these reductions were mitigated by the administration of BPE (Fig. 2C, D).
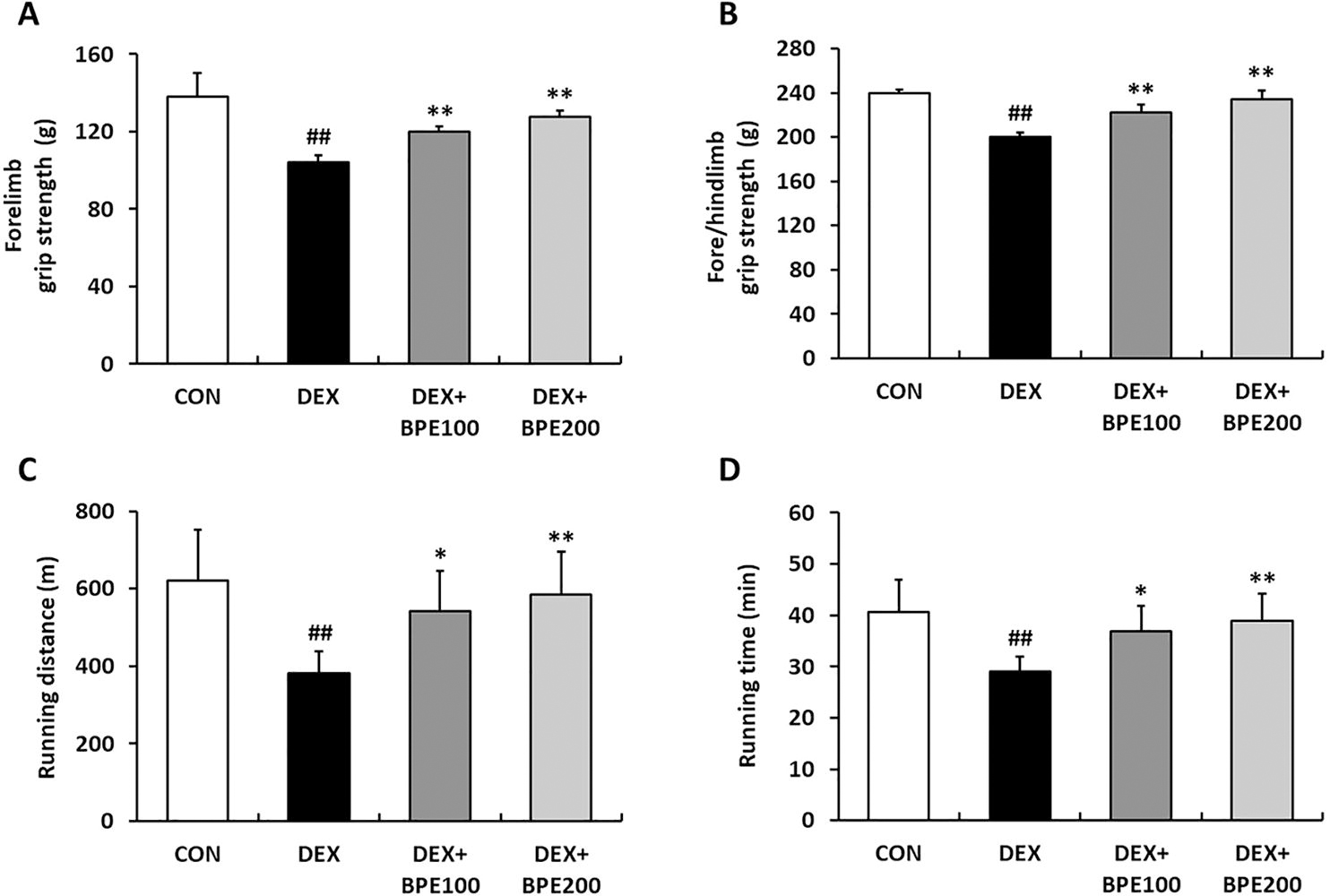
Effects of BPE on muscle function in mice with DEX-induced muscle atrophy.
BPE attenuated inflammatory responses and increased antioxidant enzymes in DEX-induced mice
The phosphorylated protein expression of NF-κB was elevated in the DEX group. However, this elevation was attenuated by BPE administration (Fig. 3A). Additionally, the mRNA expressions of TNF-α and interleukin (IL)-6, which are downstream factors of NF-κB, were markedly upregulated in the DEX group relative to those in the CON group, suggesting an increase in inflammatory responses. However, BPE dose-dependently suppressed these increases (Fig. 3B). The mRNA expressions of antioxidant enzymes, including catalase (CAT), superoxide dismutase (SOD), and glutathione peroxidase (GPx), were decreased in the DEX group, whereas BPE significantly alleviated this downregulation (Fig. 3C).
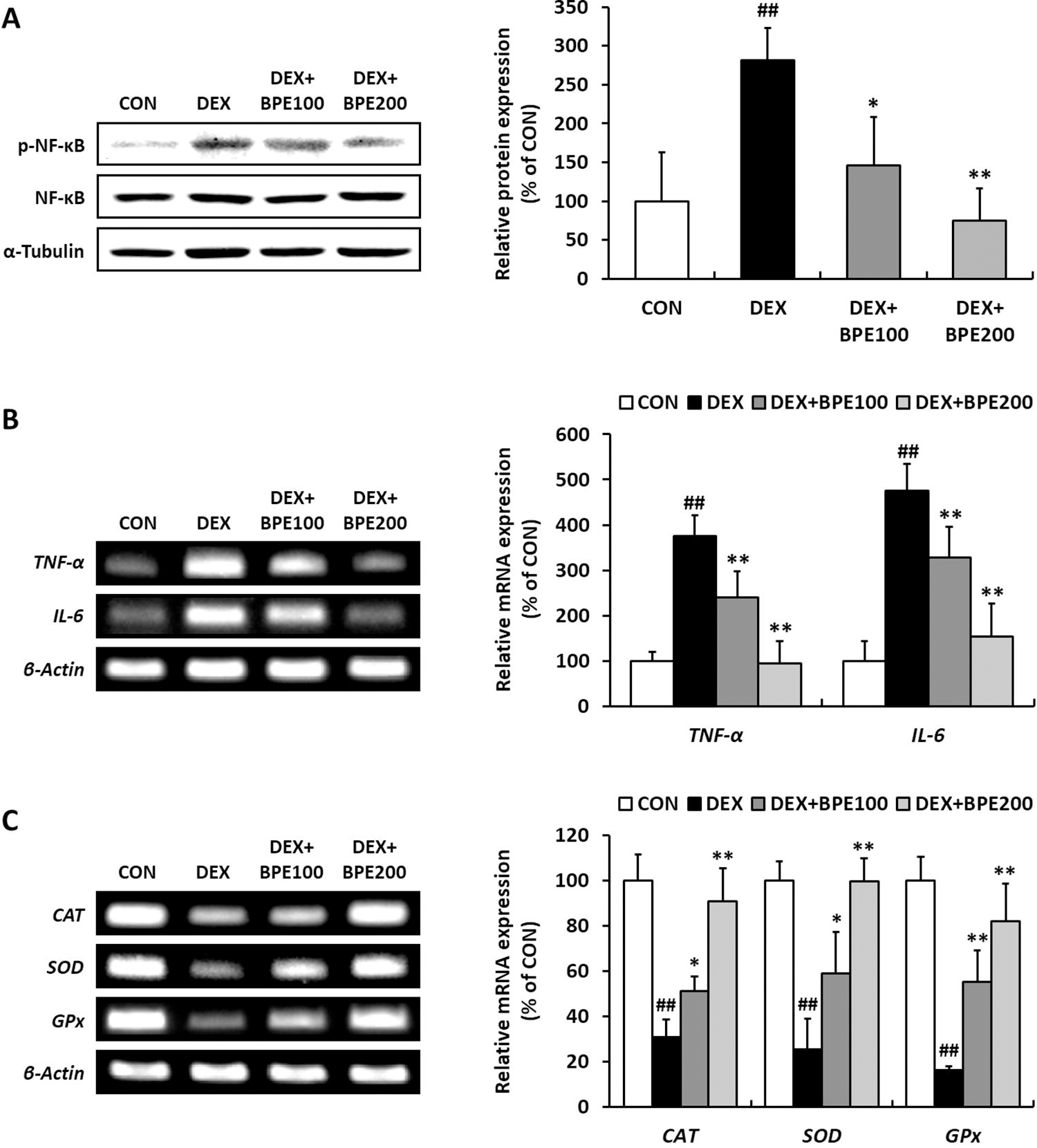
Effects of BPE on inflammatory responses and antioxidant enzymes in mice with DEX-induced muscle atrophy.
BPE suppressed protein degradation in DEX-induced mice
The DEX group demonstrated a significant upregulation in the mRNA expressions of Mstn, MuRF1, and atrogin-1 compared with the CON group. Conversely, the group treated with BPE demonstrated a marked reduction in the expressions of these markers relative to the DEX group (Fig. 4A). Additionally, the DEX group exhibited the reduced protein expression of p-FoxO3a relative to the CON group, indicating activation of the FoxO3a signaling pathway. However, BPE treatment dramatically restored the expression of p-FoxO3a (Fig. 4B).

Effects of BPE on protein degradation in mice with DEX-induced muscle atrophy.
BPE increased protein synthesis and myogenic myogenesis in DEX-induced mice
The protein expression of p-PI3K was decreased, followed by a significant reduction of p-Akt expression in the DEX group. However, BPE treatment relieved DEX-mediated downregulation of p-PI3K, p-Akt (Fig. 5A). In addition, the protein expressions of p-mTOR, p-p70S6K, and p-4EBP1, which are downstream factors of the PI3K/Akt pathway, were decreased in the DEX group, whereas treatment with BPE effectively restored the protein expressions of these factors (Fig. 5B).

Effects of BPE on protein synthesis and myogenesis in mice with DEX-induced muscle atrophy.
The mRNA expressions of myogenic regulatory factors (MRFs), including myogenic determination protein (MyoD), myogenin (MyoG), and myosin heavy chain (MHC), were markedly inhibited in the DEX group. However, the administration of BPE alleviated this inhibition. In particular, 200 mg/kg BPE significantly increased the expressions of all these factors (Fig. 5C).
DISCUSSION
Persistent inflammation and oxidative stress are major factors in the progression of muscle atrophy. 5 Inflammatory cytokines, particularly TNF-α and IL-6, enhance protein degradation within the body. 15,16 These inflammatory cytokines lead to oxidative stress by elevating ROS levels, which subsequently activates the NF-κB signaling pathway and thereby upregulates E3 ubiquitin ligases. Skeletal muscles are particularly rich in mitochondria due to their high energy consumption compared to other tissues. ROS, generated as by-products of oxidative phosphorylation in the mitochondria, are essential for maintaining processes such as muscle development, mitochondrial biogenesis, and muscle recovery. However, the upregulation of inflammatory cytokines and ROS contributes to cellular damage and aging, thereby playing a role in the occurrence and progression of muscle atrophy. 17 A prior study reported that TNF-α treatment excessively increased ROS production in L6 myotubes, indicating that inflammatory and oxidative stress are not independent phenomena interrelated in atrophic muscle. 13 In contrast, antioxidant enzymes, such as CAT, SOD, and GPx, act as scavengers for ROS, protecting cells from oxidative stress and thereby preventing muscle wasting. 18 Numerous researches have evaluated the anti-inflammatory and antioxidant effects of B. pandurata and its active compounds. BPE decreased the levels of NF-κB, TNF-α, and IL-1β in hairless mice with atopic dermatitis-like skin lesions. 19 Additionally, BPE suppressed the development of periodontitis, an inflammatory disease, through the downregulation of NF-κB and IL-1β levels in both lipopolysaccharide-treated and aged rats. 20,21 Moreover, panduratin A and hydroxypanduratin A, compounds found in B. pandurata, have been noted to exhibit anti-inflammatory and antioxidant effects. 12,22 Consistent with previous findings, this study found that BPE decreased the protein level of p-NF-κB and the mRNA levels of TNF-α and IL-6 while concurrently elevating the mRNA levels of CAT, SOD, and GPx. These findings highlight BPE’s potential to inhibit muscle atrophy through anti-inflammatory and antioxidant effects.
Mstn and FoxO3a act as negative regulators of muscle hypertrophy, mediating the catabolic response within muscle tissues. Mstn antagonism has been demonstrated to improve muscle function and muscle mass in both aged and young mice. 23 Moreover, individuals exhibiting dysfunctional mutations in the Mstn gene have been observed to have improved muscle mass compared with those with normal Mstn function. 24 Thus, targeting Mstn and its downstream factors could offer a viable therapeutic strategy for managing muscle wasting. Increased Mstn levels adversely affect the PI3K/Akt pathway activation and suppress phosphorylation of FoxO3a. Dephosphorylation of FoxO3a contributes to muscle atrophy by stimulating protein degradation through the upregulation of its downstream factors, MuRF1 and atrogin-1. 4 Numerous studies have shown that in a muscle-atrophy mouse model induced by DEX, the levels of Mstn, MuRF1, and atrogin-1 were highly regulated, whereas the level of phosphorylated FoxO3a was suppressed. 25,26 Consistently, the present study demonstrated that the mRNA levels of Mstn, MuRF1, and atrogin-1 were elevated, whereas the protein level of p-FoxO3a was diminished in the DEX group. However, BPE treatment significantly mitigated these expression alterations induced by DEX administration. These results indicate that BPE may alleviate muscle atrophy by regulating Mstn and its downstream factors.
As shown in Figure 5, our results strongly suggest that the administration of BPE can mitigate muscle wasting by significantly enhancing protein synthesis and myogenesis. The PI3K/Akt signaling pathway is recognized as a critical regulator of both muscle protein synthesis and muscle differentiation. Stimulation of this pathway results in the successive activation of mTOR and its downstream factors, p70S6K and 4EBP1, via phosphorylation, thereby inducing protein synthesis. It also promotes myogenesis by enhancing MRFs, including MyoD, MyoG, and MHC, which are crucial for muscle fiber development and differentiation. 6 Additionally, it prevents the dephosphorylation of FoxO3a, thereby suppressing its translocation to the nucleus and ultimately inhibiting proteolysis. 3 Previous studies have reported that mice lacking Akt demonstrated a significant reduction in muscle mass, grip strength, and CSA. 27,28 Consequently, focusing on the PI3K/Akt pathway could be considered a promising intervention for preserving muscle mass. Several studies have indicated that in a muscle-atrophy mouse model induced by DEX, both the PI3K/Akt pathway and MRFs were diminished. 8,29 Consistently, the present study observed a considerable decrease in the protein levels of p-PI3K, p-Akt, p-mTOR, p-p70S6K, and p-4EBP1, as well as the mRNA levels of MyoD, MyoG, and MHC following DEX administration. However, treatment with BPE resulted in an improvement in their expression levels. Additionally, a prior study showed that panduratin A elevated the expression levels of p-PI3K, p-Akt, p-mTOR, p-70S6K, p-4EBP1, MyoG, and MyoD in TNF-α-treated L6 skeletal muscle cells. 13 Thus, these findings indicate that BPE may mitigate muscle wasting by promoting the PI3K/Akt pathway and its associated downstream signaling mechanisms.
Chalcone substantially contributes to the prevention of muscle wasting. Additionally, 4-hydroxyderricin stimulated MRFs, including MHC, MyoG, and MyoD, through p38 protein activation. Furthermore, it downregulated protein degradation-related factors, such as Mstn, MuRF1, and atrogin-1 in C2C12 myotubes. 30 Furthermore, licochalcone A and B, chalcone derivatives from Glycyrrhiza uralensis, promoted cell proliferation in aged cells and diminished Mstn, MuRF1, and atrogin-1 levels in C2C12 cells and mice, suggesting that their potential as treatments for muscle atrophy. 31 Particularly, panduratin A, a chalcone derivative and the main active compound in B. pandurata, inhibited muscle wasting by stimulating the PI3K/Akt pathway. 13 Considering these results, and given that B. pandurata contains a variety of chalcone compounds, 12 the presence of these constituents in BPE may be responsible for its inhibitory effects.
CONCLUSIONS
Oral administration of BPE effectively attenuated DEX-induced muscle atrophy by restoring the imbalance of protein metabolism. Therefore, BPE may have the potential as a functional food ingredient for preventing or treating muscle wasting conditions, such as sarcopenia and muscle atrophy. However, for commercial use, further studies must be conducted to verify the effectiveness and safety of BPE through clinical trials on patients with muscle atrophy.
Footnotes
AUTHORS’ CONTRIBUTIONS
M.K.: Writing—original draft, methodology, investigation, validation, and formal analysis. T.K.: Writing—review and editing, investigation, and validation. H.J.: Investigation, validation, and formal analysis. D.K.: Supervision, project administration, and funding acquisition. D.-W.L.: Supervision, project administration, and funding acquisition. J.-K.H.: Conceptualization, supervision, project administration, and funding acquisition.
AUTHOR DISCLOSURE STATEMENT
The authors declare that they have no conflicts of interest.
FUNDING INFORMATION
This work was supported by a grant from the Ministry of SMEs and Startups (
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.