
Abstract
Our aim in the current study was to determine the in vitro and in vivo synergistic antiinflammatory and antiallergic effect associated with the IL-12 production of guaijaverin and epigallocatechin gallate (EGCG) complex (GEC) and ILS-F-2301 (2:8 extract of Psidium guajava and Camellia sinensis). Compared to EGCG alone, GEC showed synergistic inhibition of nitric oxide (NO), inducible NO synthase, and cyclooxygenase-2 by 3.8, 5.1, and 4.1%, respectively. The downregulation of interleukin-12 (IL-12) by 2,4-dinitrophenyl-human serum albumin conjugate/DNP-immunoglobulin E or ovalbumin (OVA) was synergistically increased by GEC by about 7.5% or 5.4% compared to EGCG alone. The level of downregulation of IL-12 in plasma increased by 100 mg/kg with ILS-F-2301 (28.7%) when compared to the OVA/Alu-treated group. Also, GEC synergistically increased by GEC by about 7.5% or 5.4% compared to EGCG alone. The level of down and cyclooxygenase C synergistically inhibited p-Akt, PI3K, mTOR, p-STAT6, and GATA3 by 4.9%, 4.1%, 19.2%, 23.8%, and 35.3%, respectively, while increasing the expressions of p-STAT1 and T-bet (showing 53.3% and 9.4% activation) when compared to EGCG alone. In an allergenic rhinitis mouse model, 100 mg/kg of ILS-F-2301 was shown to inhibit p-Akt, PI3K, mTOR, p-c-Jun N-terminal kinase (p-JNK), p-extracellular signal-regulated kinase (p-ERK), and p-p38 by 23.3%, 43.8%, 17.2%, 32.2%, 29.1%, and 41.8% when compared to the OVA/Alu-sensitized group. Taken together, our findings suggest that ILS-F-2301 may have potential as a functional food for alleviating antiallergic rhinitis.
INTRODUCTION
Allergic rhinitis (AR) is a major allergic disease that is known to be an allergen-induced. AR induces various allergic symptoms including runny nose, nasal itching, nasal mucus, and repetitive and convulsive sneezing. 1 Many studies have reported that 40% of the population has suffered from allergen-mediated AR. AR is caused by house dust mites or animal skin, which are known to cause allergic symptoms. 1,2
AR and asthma are caused by exposure to an allergen, and these are diagnosed as allergic disorders. Allergic reactions are associated with decreased production of antiinflammatory cytokines (interferon gamma [IFN-γ] and interleukin-12 [IL-12]) along with increased production of proinflammatory cytokines (IL-4 and -13). 3 –5 Each cytokine helps T-cell differentiation by activating a specific type of transcription factor. For T helper 1 (Th1) cells, IL-12 and IFN-γ activate the differentiation of naïve CD4+ T cells by increasing the expression of the transcription factors signal transducer as well as the activations of both transcription 1 (STAT1) and T-box transcription factor TBX21 (T-bet). 3,4 IL-12 is classified based on Th1 cytokine and IL-12 production-regulated CD4+ T cell differentiation; this makes the secretion of IL-12 in mast cells an important marker for evaluating Th1 cytokine regulation. In Th2 cell differentiation, the expression of the transcription factors signal transducer and the activator of transcription 6 (STAT6) and GATA-binding protein 3 (GATA3) are increased and activated by IL-4. 5 When the balance of Th1/Th2 cells is disrupted, an allergic reaction is induced.
Guaijaverin is known for its antiplaque 6 and antidiabetic activities. 7 Moreover, a previous study of ours confirmed that guaijaverin exerts antiallergenic activity through the STAT1/T-bet and STAT6/GATA3 signaling pathway in ovalbumin (OVA)-treated EL-4T lymphocytes. 8 However, we did not study the effect of the mixture extract of Psidium guajava leaf and Cammellia sinensis leaf (ILS-F-2301) and its active compounds, guaijaverin and epigallocatechin gallate (EGCG) on the production of IL-12, a Th-1-related cytokine. Therefore, we evaluated the synergistic effects on IL-12 production of complexes of guaijaverin and EGCG (GEC), and their application material, ILS-F − 2301, using 2,4-dinitrophenyl-human serum albumin conjugate (DNP-HSA)/DNP-immunoglobulin E (DNP-IgE)-sensitized RBL-2H3 mast cells, OVA-treated EL-4T lymphocytes, and the OVA/aluminum hydroxide (Alu)-sensitized mouse model. We also confirmed the synergistic antiinflammatory effect of GEC and ILS-F-2301 in lipopolysaccharide (LPS)-stimulated Raw264.7 macrophages.
P. guajava (guava), which belongs to the Myrtaceae family, is a traditional plant that has been shown to have antispasmodic, 9 antiinflammatory, 10 antidiarrheic, 11 antihypertensive, 12 antiobesity, 13 and antidiabetic effects. 14 –17 Moreover, our previous study confirmed the antiallergenic effect of P. guajava leaf extract in DNP-HSA/DNP-IgE-sensitized RBL-2H3 mast cells. We also identified four major/active compounds (quercetin, ellagic acid, guaijaverin, and avicularin) in P. guajava leaf extract, and guaijaverin is the main active compound in antiallergenic rhinitis.
C. sinensis (green and black tea) belongs to the Theaceae family, and it has anticariogenic, 18 antiinflammatory, 19 and antioxidant 20 activities through EGCG. Moreover, data from our previous study suggested that C. sinensis extract had an antiallergic effect, as did its active compounds, caffeine, EGCG, and epicatechin gallate (ECG). EGCG is also the main active compound determining the antiallergenic effect of C. sinensis extract.
Our study is the first to investigate the synergistic antiinflammatory and antiallergenic effects of GEC. We have also confirmed the synergistic antiinflammatory mechanism and antiallergenic mechanism of GEC and its mixture extract (ILS-F-2301) in DNP-HSA/DNP-IgE-sensitized RBL-2H3 mast cells and an OVA-sensitized mouse model through modulation of the Th1-related proteins STAT1/T-bet and cytokine IL-12, as well as the Th2-related proteins STAT6/GATA3.
MATERIALS AND METHODS
Materials
Dulbecco’s modified Eagle medium (DMEM), fetal bovine serum (FBS), minimum essential medium (MEM), and penicillin-streptomycin were obtained from Welgene (Kyongsan-si, Republic of Korea). Alu, DNP-HSA, DNP-IgE, methanol, ethanol, LPS, Griess reagent, and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazilium (MTT) reagent were all purchased from Sigma-Aldrich (St. Louis, MO, USA). The antibodies to p-protein kinase B (p-Akt, #4060), p-extracellular signal-regulated kinase (p-ERK, #4370), ERK (#4695), p-c-Jun N-terminal kinase (p-JNK, #4668), JNK (#9252), p-p38 (#4511), p38 (#9212), cyclooxygenase-2 (COX-2, #12282), p-phosphoinositide 3-kinase (p-PI3K, #4228), p-mammalian target of rapamycin (p-mTOR, #5536), p-STAT1 (#8826), β-actin (#8457), antirabbit secondary antibody (#7074), and antimouse secondary antibody (#7076) were purchased from Cell Signaling Technology (Danvers, MA, USA). Inducible nitric oxide synthase (iNOS, sc-7271), GATA3 (sc-514427), and T-bet (sc-21749) were purchased from Santa Cruz Biotechnology (CA, USA). Finally, p-STAT6 (ab217998) was purchased from Abcam Biotechnology (MA, USA).
Preparation of the ILS-F-2301
P. guajava L. (Kota Bogor, Indonesia) and C. sinensis (Ha-dong, Republic of Korea) were extracted with 50% ethanol under reflux at 70 ± 2°C for 3 h, then subjected to filtration using a 50-μm filter, concentration at 50°C for 3 h, and drying at 85°C for 12–24 h. Each P. guajava or C. sinensis extract powder homogenate was able to pass through 40 mesh. The ILS-F-2301 was prepared by mixing P. guajava L. and C. sinensis extract powders in a ratio of 2:8, and then storing at −20°C until use. In a previous study, we successfully identified major and/or active compounds in ILS-F-2301. On the basis of HPLC-PDA data, we could calculate the contents of guaijaverin (0.039 μg/mL) and EGCG (5.275 μg/mL) of 50 μg/mL in ILS-F-2301. For confirming the antiinflammatory and antiallergenic effects of ILS-F-2301, we used the calculated concentrations of guaijaverin and EGCG in the experiment.
Cell culture
Raw264.7 macrophages or RBL-2H3 mast cells or EL-4T lymphocytes were obtained from the Korean Cell Line Bank (Seoul, Korea). Raw264.7 cells or EL-4T lymphocytes were cultured in DMEM supplemented with 10% FBS and 1% penicillin-streptomycin in an incubator with 5% CO2 at 37°C. Moreover, RBL-2H3 cells were cultured in MEM supplemented with 10% FBS and 1% penicillin-streptomycin in an incubator with 5% CO2 at 37°C.
Cell viability
Raw264.7 cells (1 × 105 cells/well) or RBL-2H3 mast cells (1 × 105 cells/well) or EL-4T lymphocyte (1 × 105 cells/well) were seeded into 96-well plates at 37°C for 24 h, after which ILS-F-2301 (50, 100, and 250 μg/mL), guaijaverin (0.039, 0.25, 0.5, and 1 μg/mL), EGCG (0.5, 1, and 5.275 μg/mL), and GEC (mixture of 0.039 μg/mL of guaijaverin and 5.275 μg/mL of EGCG corresponding to 50 μg/mL of ILS-F-2301) were added to the wells at 37°C for 24 h. After 24 h of incubation, WST reagent was added, after which the plates were incubated at 37°C for 4 h. After this additional 4 h of incubation, cell viability was measured using an ELISA reader at 550 nm wavelength. Viability was expressed in terms of the percentage of live cells in each well.
Nitric oxide production
Briefly, Raw264.7 cells (5 × 104 cells/well) were seeded into 96-well plates and incubated at 37°C for 24 h. After 24 h of incubation, each well was treated with ILS-F-2301 (50, 100, and 250 μg/mL), guaijaverin (0.039, 0.25, 0.5, and 1 μg/mL), EGCG (0.5, 1, and 5.275 μg/mL), or GEC (mixture of 0.039 μg/mL of guaijaverin and 5.275 μg/mL of EGCG corresponding to 50 μg/mL of ILS-F-2301), followed by treatment with LPS (100 ng/mL) at 37°C for 24 h. After 24 h of incubation, the supernatant was collected, and the amount of nitric oxide (NO) production was measured using Griess reagent. The NO content was measured with an ELISA reader at 540 nm.
IL-12 production
RBL-2H3 cells (1 × 105 cells/well) were harvested by trypsinization, transferred to 24-well plates, and incubated overnight with DNP-IgE (50 ng/mL). After this 24 h of incubation, the cells were washed twice with MEM medium containing 1% FBS, and were then added to MEM at 37°C for 20 min. After this additional 20 min of incubation, ILS-F-2301 (50, 100, and 250 μg/mL), guaijaverin (0.039, 0.25, 0.5, and 1 μg/mL), EGCG (0.5, 1, and 5.275 μg/mL), or GEC (mixture of 0.039 μg/mL of guaijaverin and 5.275 μg/mL of EGCG corresponding to 50 μg/mL of ILS-F-2301) was added to each well at 37°C for 1 h. After this 1 h of incubation, DNP-HSA (100 ng/mL) was added for 4 h at 37°C.
EL-4T lymphocytes (2 × 105 cells/well) were seeded into 24-well cell culture plates, and then incubated for 24 h at 37°C. After this 24 h of incubation, ILS-F-2301 (50, 100, and 250 μg/mL), guaijaverin (0.039, 0.25, 0.5, and 1 μg/mL), EGCG (0.5, 1, and 5.275 μg/mL), or GEC (mixture of 0.039 μg/mL of guaijaverin and 5.275 μg/mL of EGCG corresponding to 50 μg/mL of ILS-F-2301) plus OVA (200 μg/mL) was added to each well for 24 h.
Supernatants from RBL-2H3 mast cells or EL-4T lymphocytes were analyzed with a specific focus on their IL-12 concentrations using ELISA (Abcam biotechnology, MA, USA) kits while following the manufacturer’s instructions.
Animals
Six-week-old male BALB/c mice were purchased from Hana Bio (Gyeonggi-do, Republic of Korea). All protocols for animal use in this study were reviewed and approved by the SEMI Institutional Animal Care and Use Committee (SEMI-23-011). The mice were housed under controlled conditions of temperature of 23 ± 3°C with relative humidity of 40–60% and 12 h light/dark cycles. Food and water were provided ad libitum.
Sensitization and challenges
The in vivo anti-AR model used in the current work was based on a modification of a previously described model. 21,22 The BALB/c mice were divided into five groups (seven mice/group): (1) normal group (N)—no sensitization; (2) OVA/Alu group (OVA/Alu)—sensitization with OVA (50 μg) and Alu (2 mg); (3) 100 mg/kg of ILS-F-2301 group—sensitization with OVA and Alu plus 100 mg/kg of ILS-F-2301 by oral administration (per os; p.o.); (4) 150 mg/kg of ILS-F-2301 group—sensitization with OVA and Alu plus 150 mg/kg of ILS-F-2301 by oral administration (p.o.); and (5) 200 mg/kg of ILS-F-2301 group—sensitization with OVA and Alu plus 200 mg/kg of ILS-F-2301 by oral administration (p.o.).
After a 1-week acclimation period, the mice were OVA-sensitized by intraperitoneal injection of 50 μg OVA in 200 μL phosphate-buffered saline (PBS) containing 2 mg alum on days 14 and 28 (Fig. 1). Sensitization was followed by daily intranasal challenges for 7 days with OVA/Alu. The mice were sacrificed one day after the last challenge on day 35. The mice were treated with ILS-F-2301 at doses of 100, 150, and 200 mg/kg by p.o. from days 7 to 35. The mice in the control and OVA/Alu groups were administered saline by p.o. At the end of 4 weeks, all mice were fasted overnight before being euthanized with CO2.

Animal design of allergic rhinitis.
Plasma analysis
Plasma was collected from the blood of the sacrificed mice. The production of IL-12 was measured using ELISA kits (Abcam, CA, USA) according to the manufacturer’s protocols.
Western blot analysis
Briefly, Raw264.7 cells (1 × 105 cells/well) or RBL-2H3 cells (1 × 105 cells/well) were seeded into 6-well cell culture plates and then incubated for 24 h at 37°C. After 24 h of incubation, ILS-F-2301 (50, 100, and 250 μg/mL), guaijaverin (0.039 μg/mL), EGCG (5.275 μg/mL), or GEC (mixture of 0.039 μg/mL of guaijaverin and 5.275 μg/mL of EGCG corresponding to 50 μg/mL of ILS-F-2301) plus LPS (100 ng/mL) were added to Raw264.7 cells. In addition, samples were treated with RBL-2H3 cells in the same way to confirm the antiallergenic mechanism of the sample. Raw264.7 cells or RBL-2H3 cells were harvested, after which they were lysed with RIPA buffer containing phosphatase/protease cocktails. The collected liver or lung tissues were also lysed using a tissue extraction buffer. AR and asthma are classified the allergic disorders. In addition, many research teams use the OVA/Alu sensitization model to evaluate pulmonary inflammatory through the activation of MAPK pathway and liver disease through PI3K/mTOR pathway. 23 –25 The protein concentration of the samples was estimated by the BCA protein assay (Gendepot, Barker, TX, USA) before the addition of the loading buffer. Samples were then heated at 100°C for 5 min and separated at 20 μg of protein/lane by 10% sodium dodecyl sulfate (SDS) polyacrylamide gel electrophoresis (SDS-PAGE) before electro-transfer to polyvinylidene (PVDF) membranes. After blocking with 5% skim milk for 1 h, the PVDF membranes were incubated overnight at 4°C with primary antibodies (Raw264.7: iNOS, COX-2; RBL-2H3: p-Akt, p-mTOR, p-PI3K, p-STAT1, T-bet, p-STAT6, GATA3; liver tissue: p-Akt, p-PI3K, p-mTOR; lung tissue: p-ERK, ERK, p-JNK, JNK, p-p38, p38). Following incubation with secondary antibodies, signals were detected using enhanced chemiluminescence (ECL; ThermoFisher Scientific, Rockford, IL, USA).
Statistical analysis
All statistical analyses were performed using SPSS v21.0 software (SPSS, Chicago, IL, USA). Unless stated otherwise, data are shown in the form of the means ± standard errors of means (SEM) of three independent experiments conducted in vitro and seven independent experiments conducted in vivo, unless stated otherwise. Differences between treatment groups were evaluated using one-way analysis of variance (ANOVA) followed by Tukey–Kramer test; P< .05 was considered to indicate a statistically significant difference.
RESULTS
Synergistic antiinflammatory effect and its mechanism of GEC in LPS-treated Raw264.7 cells
To investigate the synergistic antiinflammatory effect of GEC, we tested the inhibition of NO production in GEC in LPS-stimulated Raw264.7 cells using Griess reagent. NO is a major biomarker that is well known for its reaction to inflammation, which is useful for evaluating antiinflammatory effects. In Figure 2a, ILS-F-2301, guaijaverin, EGCG, and GEC exhibited no cytotoxicity at any concentrations in LPS-treated Raw264.7 cells. LPS stimulated NO production about 5.2-fold (P < .001) when compared to untreated cells (Fig. 2b). GEC also exhibited synergistic inhibition of NO production by 3.86% (P < .05) when compared with the corresponding value of 5.275 μg/mL of EGCG-treated Raw264.7 cells. Furthermore, ILS-F-2301 inhibited NO production by about 22.5%, 48.2%, and 74.3% (P < .001) when compared to LPS-stimulated Raw264.7 cells.
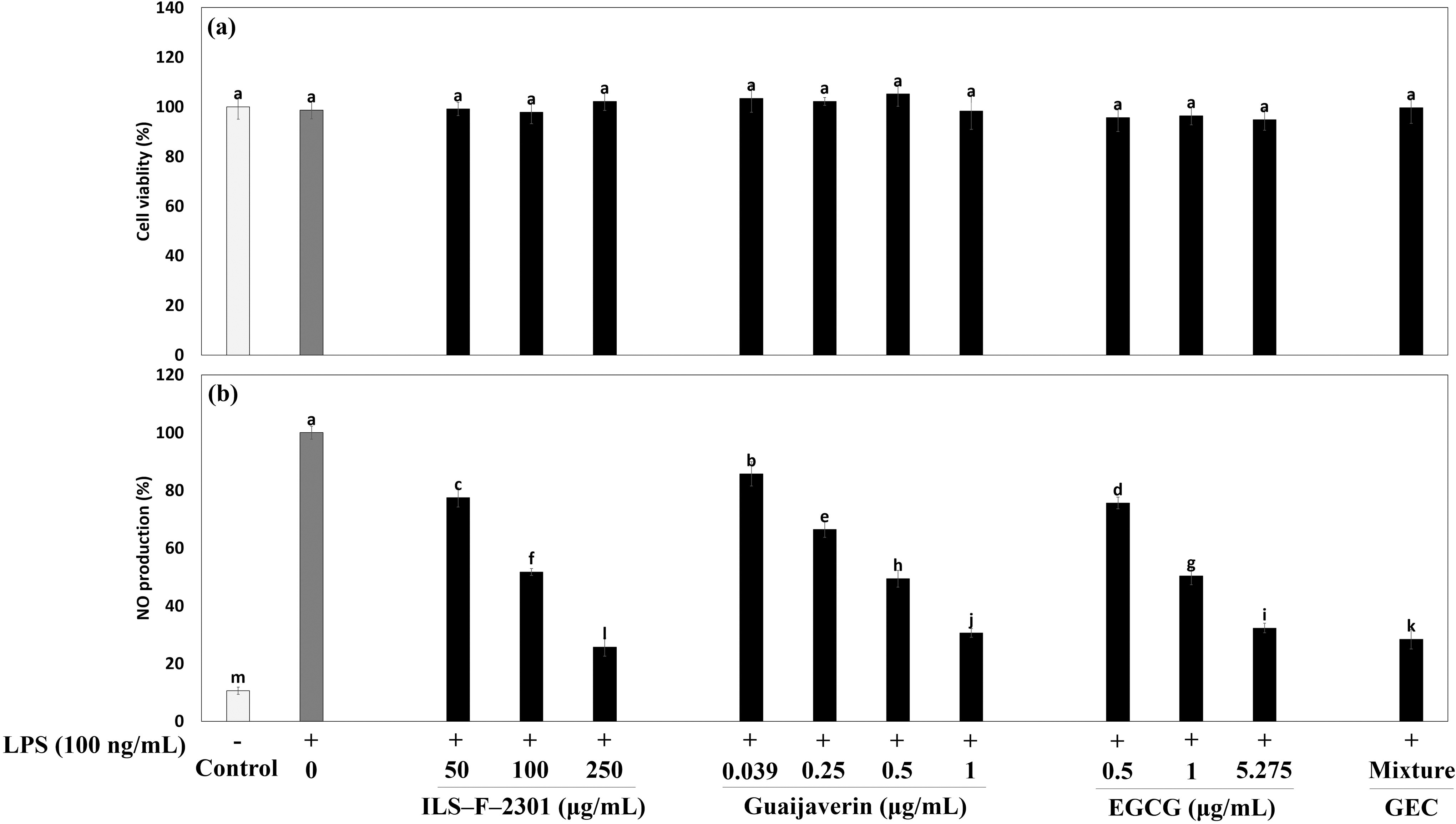
Synergistic antiinflammatory effect of GEC in LPS-treated Raw264.7 macrophages.
Next, to examine the synergistic antiinflammation mechanism of GEC, we used Western blot analysis to investigate iNOS and COX-2 protein expression. Overexpressed iNOS and COX-2 proteins by LPS were significantly suppressed by GEC by about 5.1% (P < .05) and 4.1% (P < .001), respectively, when compared to the corresponding value of 5.275 μg/mL seen with EGCG alone (Fig. 3, lanes 7 and 8). These data suggest that GEC had a synergistic antiinflammatory effect in LPS-treated cells. At 250 μg/mL, ILS-F-2301 also significantly inhibited iNOS about 1.01-fold (P < .001) and significantly inhibited COX-2 about 0.98-fold (P < .001) in terms of protein expression when compared to LPS-treated cells (Fig. 3, lanes 2–5).

Synergistic antiinflammatory mechanism of GEC in LPS-stimulated Raw264.7 macrophages. Briefly, Raw264.7 cells (1 ×105 cells/well) were incubated overnight with LPS (100 ng/mL), and then guaijaverin, EGCG, GEC, and ILS-F-2301 were added to wells for 24 h at 37°C. After 24 h of incubation, the cell pellets were collected, lysed, and Western blotted. Data are represented in the form of the mean ± standard error of mean (SEM) from three independent experiments, unless stated otherwise. Differences between treatment groups were evaluated using one-way ANOVA followed by Tukey–Kramer test, and P < .05 was considered to represent a statistically significant difference from LPS-treated cells, as determined by a one-way ANOVA. GEC: 0.037 μg/mL of guaijaverin and 5.275 μg/mL of EGCG complexes. GEC, guaijaverin and epigallocatechin gallate complex; LPS, lipopolysaccharide; EGCG, guaijaverin and epigallocatechin gallate.
Synergistic IL-12 production of ILS-F-2301 or GEC in mast cells and lymphocytes
Next, we investigated the synergistic IL-12 production of ILS-F-2301 in DNP-HSA/DNP-IgE-sensitized RBL-2H3 mast cells and OVA-treated EL-4T lymphocytes. IL-12 is classified based on Th1 cytokine and IL-12 production regulated CD4+ T cell differentiation; this makes the secretion of IL-12 in mast cells an important consideration for evaluating Th1 cytokine regulation. There were no instances of cytotoxicity among all concentrations of samples in RBL-2H3 mast cells or OVA-treated EL-4T lymphocytes (Fig. 4a and b). DNP-HSA/DNP-IgE-treated RBL-2H3 mast cells or OVA-treated EL-4 cells exhibited significantly inhibited the secretion of IL-12 about 0.18-(P < .001) and 0.66-fold (P < .001), respectively when compared to untreated cells (Fig. 5a and b). In addition, 250 μg/mL of ILS-F-2301 significantly increased the secretion of IL-12 about 4.75-(P < .001) and 3.24-fold (P < .001) compared to DNP-HSA/DNP-IgE-treated cells or OVA-treated cells. Moreover, GEC synergistically increased IL-12 production by about 7.5% (DNP-HSA/DNP-IgE-treated model, P < .001) and 5.4% (OVA-treated model, P < .001) compared to the value obtained by EGCG alone, i.e., 5.275 μg/mL.
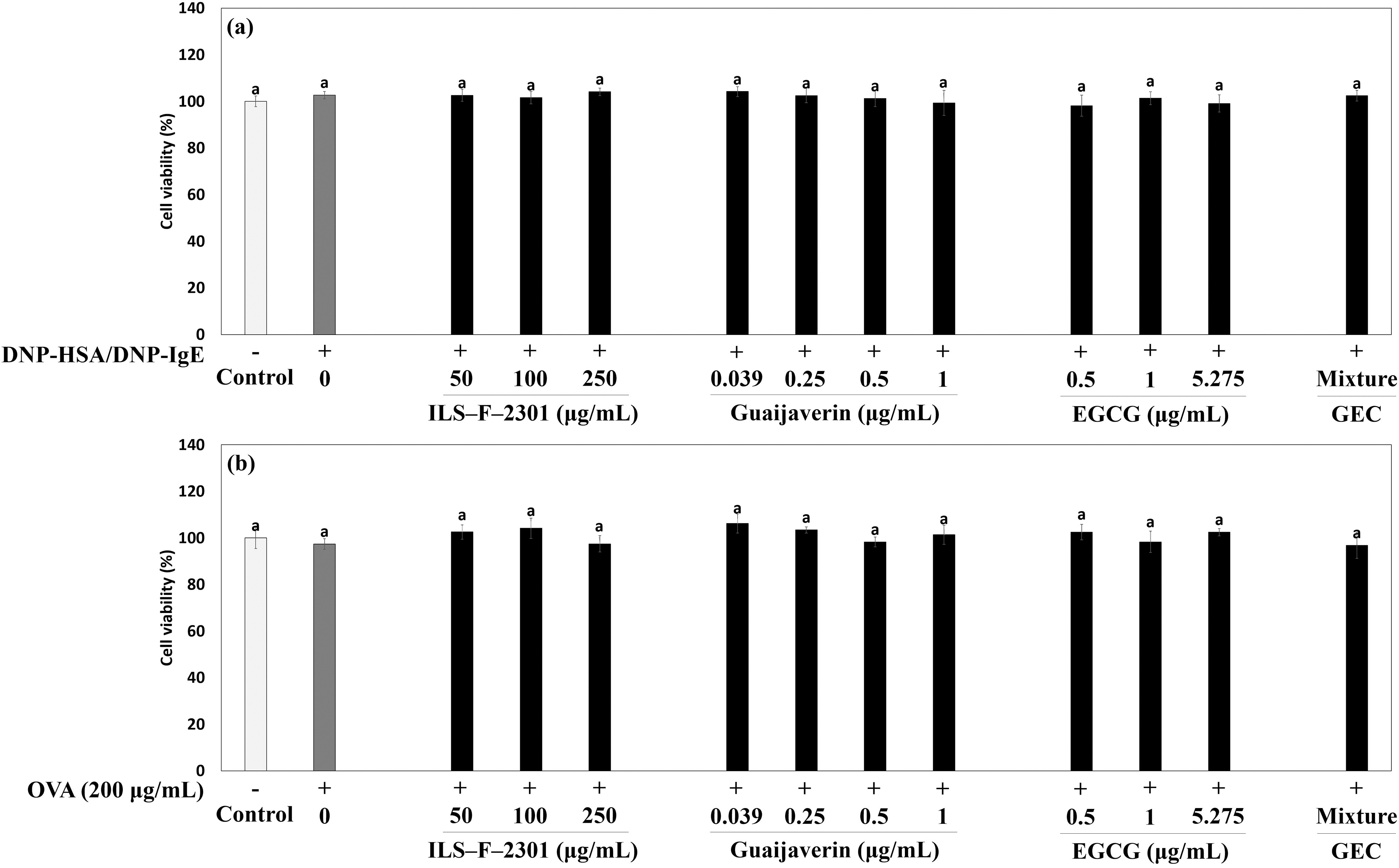
Cell viability of
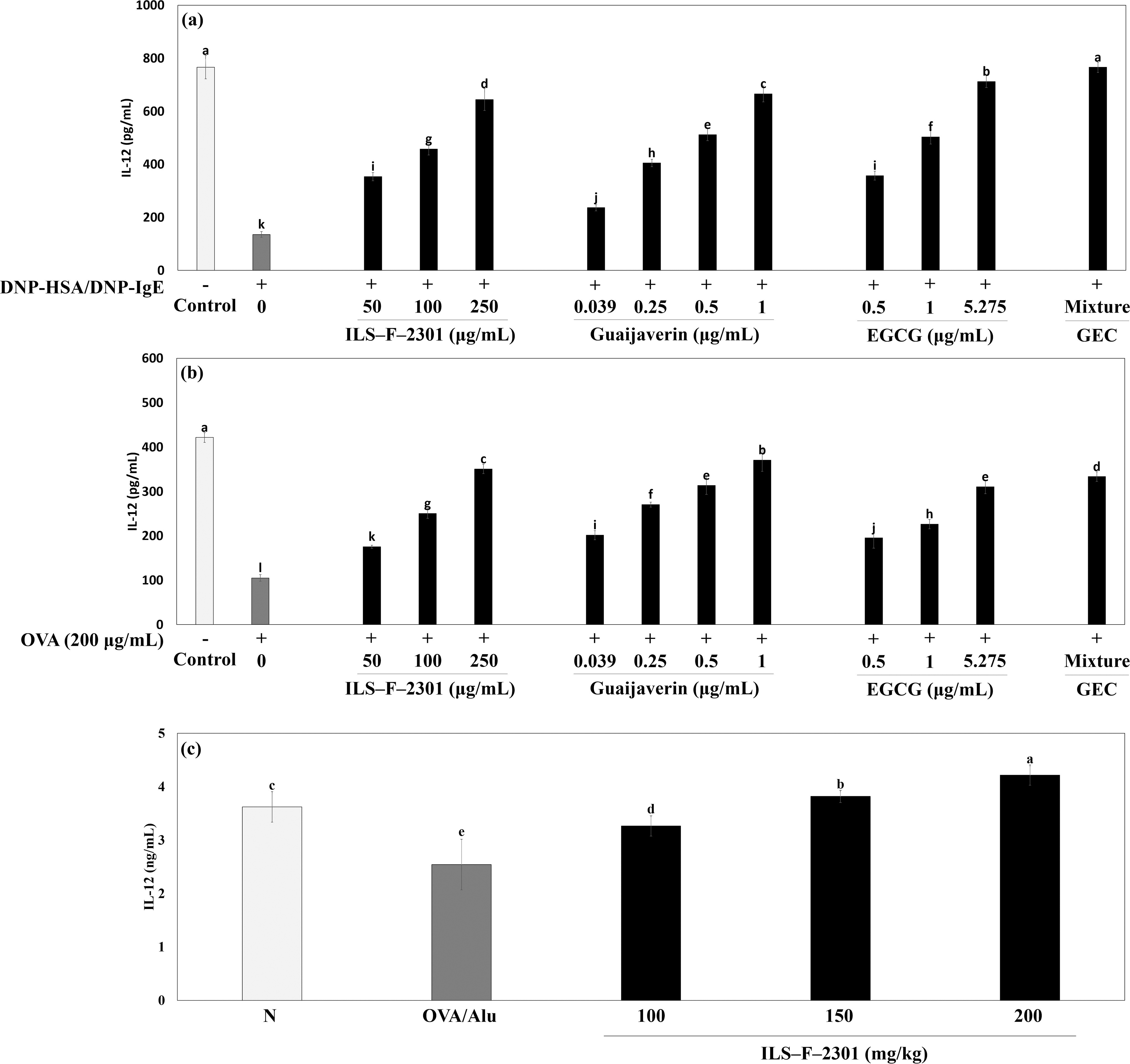
IL-12 production of GEC in
We also evaluated the production of IL-12 of ILS-F-2301 in OVA/Alu-treated mouse. IL-12 production was reduced by OVA/Alu about 0.70-fold when compared to an untreated group (Fig. 5c). ILS-F-2301 also significantly increased the production of IL-12 about 1.28-(100 mg/kg), 1.50-(150 mg/kg), and 1.66-fold (200 mg/kg) (P < .001) compared to OVA/Alu in a dose-dependent manner.
Antiallergic mechanism on ILS-F-2301 or GEC in DNP-HSA/DNP-IgE-treated RBL-2H3 cells
We also investigated the antiallergic mechanism of ILS-F-2301 and GEC using Western blot analysis. GEC synergistically inhibited the protein expressions of p-Akt, p-PI3K, and p-mTOR by about 4.9% (P < .05), 4.1% (P < .05), and 19.2% (P < .001), respectively, when compared to the value obtained by EGCG alone, i.e., 5.275 μg/mL (Fig. 6a, lanes 7 and 8). The overexpressions of p-Akt, p-PI3K, and p-mTOR by DNP-HSA/DNP-IgE were suppressed by 250 μg/mL of ILS-F-2301 by about 8.22-, 1.13-, and 0.92-fold (P < .001), respectively (Fig. 6a, lanes 2 and 5).

Synergistic modulatory mechanism of GEC in DNP-HSA/DNP-IgE-sensitized RBL-2H3 mast cells. Briefly, RBL-2H3 cells (1 × 105 cells/well) were incubated overnight with DNP-IgE (50 ng/mL), after which guaijaverin, EGCG, GEC, and ILS-F-2301 were added to wells for 1 h at 37°C. After 1 h of incubation, DNP-HSA (100 ng/mL) was added to each well at 37°C for 4 h, following which the cell supernatant was harvested and reacted with p-NAG at 37°C for 1 h. After 1 h of incubation, the cell pellets were collected, lysed, and Western blotted. Data are represented as the mean ± standard error of mean (SEM) from three independent experiments, unless stated otherwise. Differences between treatment groups were evaluated using one-way ANOVA followed by Tukey–Kramer test, and P < .05 was considered to represent a statistically significant difference from DNP-HSA/DNP-IgE-treated cells, as determined by a one-way ANOVA. GEC, guaijaverin and epigallocatechin gallate complex; DNP-HSA/DNP-IgE, 2,4-dinitrophenyl-human serum albumin conjugate/DNP-immunoglobulin E.
Next, the underexpressions of p-STAT1 and T-bet protein by DNP-HSA/DNP-IgE were significantly increased by 250 μg/mL of ILS-F-2301 by about 0.69- and 0.58-fold (P < .001), respectively (Fig. 6b, lanes 2 and 5). Moreover, the overexpressions of p-STAT6 and GATA3 protein by DNP-HSA/DNP-IgE were significantly suppressed by 250 μg/mL of ILS-F-2301 by about 0.88- and 1.11-fold (P < .001), respectively (Fig. 6b, lanes 2 and 5). Furthermore, GEC synergistically increased p-STAT1 by about 53.3% (P < .001) and T-bet by about 9.4% (P < .001), while it suppressed p-STAT6 by about 23.8% (P < .001) and GATA3 by about 35.3% (P < .001) when compared to 5.275 μg/mL of EGCG alone (Fig. 6b, lanes 7 and 8). Considered together, these data confirm that guaijaverin and EGCG in ILS-F-2301 are the main active compounds influencing antiallergic activity.
Effects of ILS-F-2301 on PI3K/mTOR signaling and MAPK signaling in OVA/alu-sensitized mice
Next, to confirm the effects of the Th1 and Th2 balance mechanisms of ILS-F-2301 and GEC, we assessed the protein expressions of PI3K/mTOR signaling and MAPK signaling in an OVA/Alu-sensitized mouse model. MAPK signaling proteins (JNK, ERK, and p38) play important roles in cell differentiation as well as the degranulation of mast cells. 21 Therefore, we investigated the regulation of PI3K/mTOR and MAPK signaling proteins using Western blot analysis. OVA/Alu-treated mice exhibit overexpressed p-Akt, p-PI3K, and p-mTOR (Fig. 7a, lane 2), whereas the group that received 100 mg/kg of ILS-F-2301 showed inhibited expressions thereof by about 3.45-, 1.73-, and 5.05-fold (P < .001), respectively (Fig. 7a, lane 5). Moreover, the expressions of p-JNK, p-EKR, and p-p38 were induced by OVA/Alu, whereas these were suppressed in the 100 mg/kg ILS-F-2301 group by about 1.03-, 0.95-, and 11.8-fold (P < .001), respectively (Fig. 7b, lanes 2 and 5).
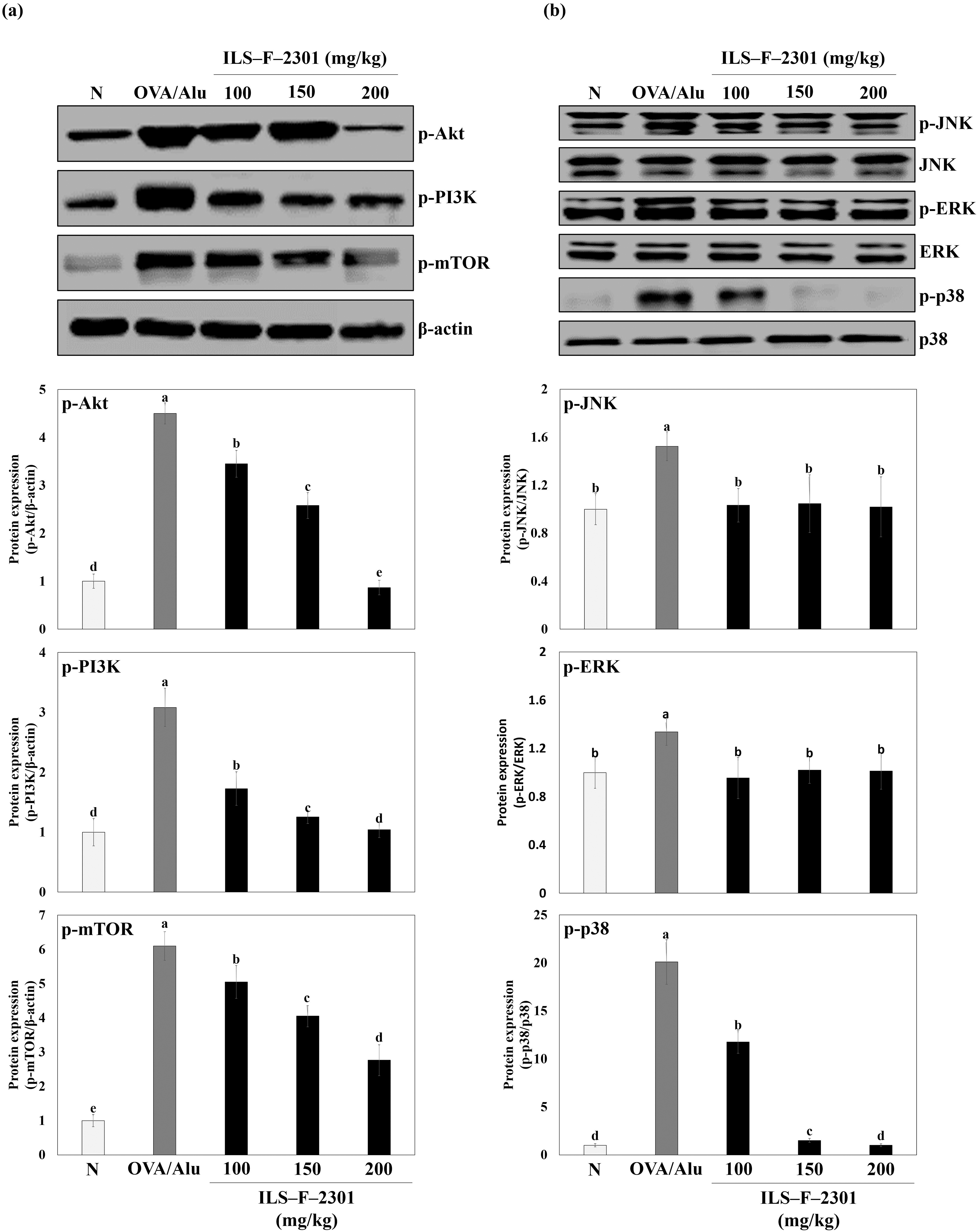
Th1 and Th2 differentiation mechanism ILS-F-2301 in OVA/Alu-sensitized mice according to dosing levels. Data are represented as the mean ± standard error of mean (SEM) from seven independent experiments, unless stated otherwise. Differences between treatment groups were evaluated using one-way ANOVA followed by Tukey–Kramer test, and P < .05 was considered to represent a statistically significant difference from OVA/Alu-sensitized mice, as determined by a one-way ANOVA. ILS-F-2301 group was administered 100, 150, or 200 mg/kg of ILS-F-2301. OVA, ovalbumin.
DISCUSSION
Guaijaverin has been reported to exhibit effective antiplaque and antidiabetic activity, but the antiallergic effect and modulatory effects on Th1 and Th2 of guaijaverin have yet to be fully investigated. In a previous study, we successfully identified seven compounds of ILS-F-2301 using HPLC-PDA analysis and confirmed that the active compounds of ILS-F-2301 were guaijaverin and EGCG. 8 Our present results also showed that GEC or ILS-F-2301 had synergistic antiallergy effects in DNP-HSA/DNP-IgE-mediated RBL-2H3 mast cells, OVA-treated EL-4T lymphocytes, and OVA/Alu-sensitized mice. However, we did not investigate the synergistic effect on the production of IL-12 (Th1 cytokine) of GEC and ILS-F-2301. Altogether, we confirmed the synergistic modulatory effect of GEC and ILS-F-2301 through the regulation of IL-12 production in vitro and in vivo.
We further investigated the synergistic antiinflammatory effect of ILS-F-2301 and GEC in LPS-stimulated Raw264.7 macrophages. NO is a critical signaling molecule that serves as a retrograde neurotransmitter and is known to be associated with neural development, immune response, angiogenesis, and is one of the vital features of inflammation. 22 NO is mediated by iNOS, and NO is released when iNOS is activated by cytokines. NO is an activating factor of COX-2. COX-2 is called prostaglandin endoperoxide synthase, and it is responsible for the biosynthesis of prostanoids, such as thromboxane and prostaglandins, from arachidonic acid. 26 COX-2 has also been shown to respond by mediating inflammatory reactions in humans. These overexpressed proteins are the main biomarkers for evaluating antiinflammatory activity. Figure 2b shows our results in investigating the synergistic effect of the inhibition of NO production by ILS-F-2301 and GEC in LPS-stimulated Raw264.7 macrophages. GEC synergistically inhibited the NO production by about 3.9% compared to the result obtained by EGCG plus LPS macrophages. Furthermore, ILS-F-2301 significantly inhibited NO production compared to LPS-stimulated macrophages. These data suggest that guaijaverin and EGCG were the main active compounds influencing the antiinflammatory effect. Subsequently, we confirmed the synergistic antiinflammatory mechanism of ILS-F-2301 and GEC. ILS-F-2301 and GEC significantly suppressed the iNOS and COX-2 protein, thereby confirming that the inhibition of NO, iNOS, and COX-2 protein expressions were related to guaijaverin and EGCG in ILS-F-2301 (Fig. 3).
Next, to investigate the synergistic production of IL-12 following ILS-F-2301 or GEC treatment, we evaluated IL-12 production using ELISA analysis. IL-12, which is well known as a Th1-related cytokine, is a major biomarker for evaluating Th1 and Th2 balance. 27 –29 ILS-F-2301 was shown to significantly increase IL-12 production compared to DNP-HSA/DNP-IgE-treated mast cells or OVA-treated lymphocytes, whereas GEC synergistically increased the IL-12 production compared to EGCG alone (Figs. 4 and 5). Moreover, the decrease in IL-12 by OVA/Alu was significantly increased by 50 mg/kg of ILS-F-2301 (Fig. 5c). These data suggest that guaijaverin and EGCG of ILS-F-2301 had an effect on Th1 cytokine production. In a previous study, we examined the modulatory effect of Th1 cytokine IFN-γ and Th2 cytokines IL-4, -5, and -13 of ILS-F-2301 and GEC in DNP-HSA/DNP-IgE-treated RBL-2H3 mast cell, OVA-treated EL-4T lymphocytes, and OVA/Alu-sensitized mouse. The Th1 cytokine IFN-γ was reduced by 250 μg/mL of ILS-F-2301 or 100 mg/kg of ILS-F-2301, whereas Th2 cytokines IL-4, -5, and -13 were inhibited in those groups. GEC also has a synergistic modulation effect on Th1 and Th2 compared to EGCG alone only.
Th1/Th2 balance is known to be a major factor in the induction of allergenic diseases, and T cells rely on mTOR signaling to maintain immune and metabolic signals in appropriate states. 23,30,31 mTOR is a serin-threonine-specific protein kinase belonging to the PI3K family that plays a central role in the body, particularly in affecting the expression of mTOR, which activates CD4+ T cells to induce Th1, Th2, and Th17 cell differentiation and under allergic conditions. 32 The overexpressions of p-Akt, p-PI3K, and p-mTOR protein were shown to be regulated by 250 μg/mL of ILS-F-2301 (Fig. 6a, lanes 2 and 5). Moreover, GEC synergistically inhibited the p-Akt, p-PI3K, and p-mTOR protein expression compared to EGCG only (Fig. 6a, lanes 7 and 8).
Next, we investigated the mechanism of Th1 and Th2 cell differentiation of GEC or ILS-F-2301 in DNP-HSA/DNP-IgE-treated mast cells using Western blot analysis. Several studies are currently examining ways to improve allergic and inflammatory reactions by regulating the activity of Th1/Th2 cells. 33 –39 ILS-F-2301 was shown to significantly increase p-STAT1 and T-bet (Th1-related protein), and it also suppressed p-STAT6 and GATA3 (Th2-related protein) compared to DNP-HSA/DNP-IgE-sensitized mast cells (Fig. 6b, lanes 2 and 5). GEC also had a synergistic modulatory effect through STAT1/T-bet and STAT6/GATA3 pathways compared to EGCG alone (Fig. 6b, lanes 7 and 8), thus confirming that guaijaverin and EGCG of ILS-F-2301 were the main active compounds in the antiallergenic effect. In a previous study, we confirmed the Th1 and Th2 modulatory effect of ILS-F-2301 as well as its active molecules guaijaverin and EGCG using OVA-treated EL-4T lymphocytes through the STAT1/T-bet and STAT6/GATA3 signaling pathway. Our findings showed that ILS-F-2301 effectively modulated the downexpressed STAT1/T-bet protein and upexpressed STAT6/GATA3.
Following our in vivo experiments, we examined the PI3K/mTOR signaling and MAPK signaling effects of ILS-F-2301. MAPKs regulate the expression of Th2 cytokines. 40 –42 Many signaling pathways in eosinophils are activated by the Th2 cytokine IL-5, including MAPKs, and the combination of these kinases and transcription factors stimulates eosinophil differentiation, survival, degranulation, and adhesion. 43 This is characteristic of a late allergenic reaction, where the main symptom is nasal congestion. The group receiving 100 mg/kg of ILS-F-2301 showed significant suppression of p-Akt, p-PI3K, and p-mTOR compared to the OVA/Alu-treated group (Fig. 7a, lanes 2 and 3). Furthermore, overexpressed p-JNK, p-ERK, and p-p38 proteins by OVA/Alu were suppressed by the administration of 100 mg/kg of ILS-F-2301 (Fig. 7b, lanes 2 and 3). Taken together, these data confirmed that ILS-F-2301 had effective antiallergenic activities through the modulation of Th1 and Th2 cell differentiation.
CONCLUSIONS
In the present work, for the first time, we demonstrate the synergistic antiinflammatory antiallergenic effects of GEC and its application example, ILS-F-2301, which are natural sources that can serve as functional foods to ameliorate AR symptoms by modulating the allergen-mediated STAT6/GATA3 and IL-12/STAT1/T-bet signaling pathways. Our findings are expected to help explain the anti-AR activity related to the combination of guaijaverin and EGCG in ILS-F-2301. We also anticipate that, if adopted by the food industry, ILS-F-2301 would show efficacy in preventing or ameliorating the symptoms of allergic rhinitis, but clinical trials are still needed to confirm efficacy in humans.
Footnotes
AUTHORS’ CONTRIBUTIONS
S.-H.P.: Conceptualization (lead); Investigation (equal); Formal analysis (lead); Supervision (equal); Visualization (lead); Writing—original draft (equal); Methodology (lead); Project administration (lead); Resources (lead); Writing review and editing (lead). P.Y.J.: Writing—original draft (supporting); Software (supporting); investigation (supporting). K.-Y.K.: formal analysis (supporting); Software (supporting). K.J.S.: Formal analysis (supporting).
DATA AVAILABILITY STATEMENT
All data are reported in the article.
ETHICS APPROVAL STATEMENT
All the procedures were performed according to the guidelines approved by the National Institutes of Health. All animal experiments were approved by the SEMI Institutional Animal Care and Use Committee (SEMI-23-011).
AUTHOR DISCLOSURE STATEMENT
The authors declare they have no conflicts of interest.
FUNDING INFORMATION
This research received no external funding.