
Abstract
The aim of this study was to determine the anti-inflammatory and anti-endoplasmic reticulum (ER) stress effects of Aruncus dioicus var. kamtschaticus (ADK) extract on ARPE-19 cells. Pretreatment with ADK effectively mitigated thapsigargin (Tg)-induced increases in vascular endothelial growth factor protein secretion and intracellular calcium levels. Furthermore, pretreatment with ADK suppressed ocular ER stress-related protein expression in a dose-dependent manner, inhibited the loss of tight junctions, and suppressed interleukin-6 gene expression. Moreover, ADK pretreatment significantly prevented lipopolysaccharide-inducible proinflammatory cytokine gene expression at the transcription level and the phosphorylation of proteins involved in the mitogen-activated protein kinase-nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB) axis at the posttranslational level. Additionally, ADK extract enhanced antioxidant activity, as evidenced by increased heme oxygenase-1 protein expression and increased 2,2-diphenyl-1-picrylhydrazyl radical scavenging and ferric-reducing antioxidant power. In conclusion, ADK extract effectively protected ARPE-19 cells from ocular ER stress, inflammation, and oxidative stress, demonstrating its potential as a nutraceutical intervention for ocular diseases.
INTRODUCTION
Diabetes is a chronic medical condition characterized by the inability to produce sufficient insulin or use insulin effectively, leading to elevated glucose levels in the bloodstream. 1 When insulin production is insufficient or insulin is ineffective, glucose accumulates in the blood, resulting in hyperglycemia. 2 Persistent hyperglycemia may result in a range of serious health complications over time, including neuropathy, nephropathy, retinopathy, cerebrovascular accidents (strokes), and peripheral arterial disease. 3,4 These metabolic complications are well documented in the medical literature, which also emphasizes the critical importance of early detection and comprehensive management of diabetes to prevent these severe outcomes. 5,6 Early intervention, including lifestyle changes, medication, and regular monitoring of blood glucose levels, plays a crucial role in mitigating the risk of complications. 7 With the progressive nature of diabetes, without appropriate management, the likelihood of developing complications increases, significantly impacting quality of life and overall health. 8,9
Among the metabolic complications induced by diabetes, diabetic retinopathy (DR) is a particularly serious issue due to its direct threat to vision. 10 DR is characterized by increased blood vessel permeability, which leads to leakage, retinal detachment, and intraocular bleeding. 11,12 These pathological changes are primarily driven by ischemic angiogenesis, which is intensified by the increased production of vascular endothelial growth factor (VEGF) and the resultant destabilization of the retinal nervous system. 13 The induction of VEGF and the resulting neovascularization are processes that are also common in other degenerative ocular diseases, such as age-related macular degeneration (AMD), underscoring the similarities between these conditions in terms of the underlying pathological mechanisms and potential therapeutic interventions. 14 An in-depth understanding of these processes is urgently needed to facilitate the development of targeted therapeutic strategies to slow the progression of DR and similar ocular diseases, thereby preserving vision and improving the quality of life for individuals affected by these severe complications of diabetes. 15
Elevated VEGF levels within the eye are closely linked to increased ER stress, a relationship substantiated by recent studies. 16 The ER, a pivotal cellular organelle, not only is essential for protein synthesis and proper folding but also plays an integral role in regulating the concentration of intracellular calcium ([Ca2+]i) and the biosynthesis of vital molecules such as phospholipids and cholesterol. 17 –20 An aberrant increase in [Ca2+]i, often resulting from abnormal metabolic activity within the ocular environment, serves as a catalyst for VEGF expression. 21 The ocular induction of [Ca2+]i triggers the production of new blood vessels, paving the way for the development of serious ocular conditions such as DR and AMD. 22 Furthermore, the production of abnormal proteins within cells exacerbates ER stress, leading to the increased activation of unfolded protein reactions (UPRs) within the ER. 23 The induction of ER stress has the dual potential to either promote chaperone production, thereby helping maintain robust cellular homeostasis, or initiate cellular apoptosis. The persistent presence of ER stress within the retinal environment is a significant contributing factor to the onset of various ocular diseases, underscoring the complex interplay between cellular stress mechanisms and ocular health. 24,25 This intricate connection highlights the critical importance of understanding ER stress pathways to develop more effective therapeutic strategies for ocular diseases associated with elevated VEGF levels and ER stress.
In clinical practice, the ocular injection of anti-VEGF agents such as Lucentis, Eylea, and Avastin has become a cornerstone for curing abnormal VEGF expression within the eye. 26 These pharmacological treatment strategies effectively inhibit VEGF activity, which in turn markedly reduces cellular proliferation and angiogenesis. 27 However, the long-term administration of these intravitreal injections has been associated with a range of potential side effects, including retinal pigment epithelial tears, significant intraocular inflammation, the development of cataracts, vitreous hemorrhage, and an increase in intraocular pressure. 26,28 In response to these challenges, recent scientific endeavors have increasingly focused on exploring natural substances as a strategy to prevent ocular diseases and mitigate the adverse effects linked to anti-VEGF injections. 29,30 Among these natural alternatives, phytochemicals, particularly polyphenols, have shown significant potential in either preventing or decelerating the progression of various ocular diseases. Phenolic compounds are recognized for their ability to modulate physiological processes, offering a promising avenue for the development of novel therapeutic strategies aimed at enhancing ocular health and reducing reliance on anti-VEGF therapies. 15,31,32
Aruncus dioicus var. kamtschaticus (ADK), a member of the Aruncus genus, is a medicinal herb that is rich in polyphenols (including flavonoids, tannins, and phenolic acids), monoterpenoids, and cyanogenic glycosides known as prunasin, as identified by Fusani et al. (2016). 33 The medicinal applications of ADK are impressive, ranging from its demonstrated antithrombotic properties in human red blood cells, which underscores its potential in circulation-related health issues, 34 to its role in supporting cognitive functions observed in C57BL/6 mice that have developed obesity from a high-fat diet, where ADK showed a notable impact on cognitive enhancement through behavioral experiments. 35 Additionally, ADK has exhibited significant anti-aging effects in human skin fibroblast models exposed to ultraviolet (UV)-B radiation, suggesting its utility in dermatological applications aimed at mitigating skin aging. 36 The antitumor effects of ADK have also been highlighted in studies involving Jurkat T cells, revealing a promising avenue for cancer research. 37 Furthermore, the efficacy of ADK extends to the prevention of acute renal injury in models of ischemia–reperfusion injury in SD rats, indicating its protective effects against renal damage. 38 Collectively, these findings not only underscore the multifaceted physiological activities of ADK but also highlight its potential as a versatile natural remedy in various health contexts, ranging from cardiovascular and cognitive health to dermatology, oncology, and nephrology. However, to date, there is little evidence regarding whether ADK can prevent pathophysiology in ocular diseases. In this study, we investigated whether pretreatment with ADK extract, which is known for various physiological activities, can prevent ER stress induced by thapsigargin (Tg) treatment in human retinal pigment epithelial (ARPE-19) cells and inflammatory reactions induced by lipopolysaccharide (LPS).
MATERIALS AND METHODS
Extraction of ADK
ADK was sourced from Gangwondo, South Korea, in 2009. The dried powder was mixed with 80% ethanol at a ratio of 1500 mL per 100 g of sample. This mixture was extracted three times, each for 3 h, using a heating mantle (MS-265; Mtops, Seoul, Korea) equipped with a reflux condenser and maintained at 65°C. The resulting solution was then filtered through qualitative filter paper (No. 2; Advantec, Tokyo, Japan). The filtrate was concentrated using a rotary vacuum evaporator (NVC-1100; EYELA, Tokyo, Japan) at 40°C to remove the ethanol. Subsequently, the concentrate was dried under vacuum, and the solid yield was calculated. To prevent oxidation, the ADK extract was stored at −70°C.
Cell culture
The ARPE-19 cell line (CRL-2302) was obtained from the American Type Culture Collection (Manassas, VA, USA) and cultured in Dulbecco’s modified Eagle’s medium (DMEM/F12; Gibco, Carlsbad, CA, USA) supplemented with 10% fetal bovine serum (Biowest, Nuaillé, France) and 1% gentamicin (Gibco). Cultures were maintained in an incubator at 37°C with 5% CO2 (Vision Scientific Co. Ltd., Daejeon, Korea).
Cell viability assay
Cell viability was assessed using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT; Biosesang, Seoul, Korea) assay. ARPE-19 cells were seeded at a density of 2.0 × 104 cells per well in a 96-well plate (SPL Life Sciences Co. Ltd.). To evaluate the cytotoxicity of ADK, cells were treated with various concentrations of ADK (0, 10, 25, 50, 100, 200, and 500 μg/mL) for 24 h, with 1% DMSO serving as the control. The subsequent MTT assay was executed according to the described method and formula.
25
Western blot analysis
ARPE-19 cells were seeded in 6-well plates (SPL Life Sciences Co. Ltd., Pocheon, Korea) at a density of 5.0 × 105 cells per well and cultured for 48 h. To initiate an anti-ER stress response, ARPE-19 cells were pretreated with various concentrations of ADK (0, 25, 50, 100, and 200 μg/mL) for 3 h before exposure to 1 μM Tg (Sigma Aldrich, St. Louis, MO, USA) for 24 h. To initiate an anti-inflammatory response, ARPE-19 cells were treated with 1 μg/mL LPS (Sigma Aldrich) for 2 h after pretreatment with ADK.
Extraction, quantification, separation, and transfer of protein from treatments were executed using previously described methods. 25,39 –42 The membranes were blocked with 5% skim milk (BD Difco, Franklin Lakes, NJ, USA) for 1 h at room temperature, followed by overnight incubation at 4°C with primary antibodies. After washing, the membranes were incubated with secondary horseradish peroxidase-conjugated antibodies at room temperature for 1 h. Blots were developed using a chemiluminescent reagent (Thermo Fisher Scientific) and visualized on a Davinch Western™ Imaging system (Davinch-K, Seoul, South Korea). Quantification was performed using ImageJ software version 1.53k (National Institutes of Health, Bethesda, MD, USA). Each band was normalized to the total protein amount of glyceraldehyde 3-phosphate dehydrogenase (GAPDH), which served as an internal loading control. The primary and secondary antibodies used for western blotting are summarized in Supplementary Table S1.
Quantitative RT-PCR analysis
Total RNA extraction, cDNA synthesis, and quantitative reverse transcription polymerase chain reaction (qRT-PCR) analysis were performed following peer-reviewed methods. 43 –47 The relative gene expression was normalized to that of 36B4 (acidic ribosomal phosphoprotein P0 [RPLP0]), which showed no significant variation with treatment. The PCR primers used are listed in Supplementary Table S2.
Measurement of intracellular calcium ([Ca2+]i)
For [Ca2+]i measurements, ARPE-19 cells were pretreated with various concentrations of ADK (0, 25, 50, 100, and 200 μg/mL) for 3 h before exposure to 1 μM Tg for 24 h. Subsequent experimental procedures were carried out as previously described. 25
Enzyme-linked immunosorbent assay
To determine VEGF protein levels, VEGF secretion into the culture medium was quantified using an enzyme-linked immunosorbent assay (ELISA) kit according to the manufacturer’s instructions (Abcam, London, UK). Initially, cells were pretreated with various concentrations of ADK (0, 25, 50, 100, and 200 μg/mL) for 3 h before exposure to 1 μM Tg for 24 h. After centrifugation, the medium was analyzed at 450 nm using a microplate reader (SpectraMax iD3, Molecular Devices). To explore the role of mitogen-activated protein kinase kinase (MEK) in this process, cells were first pretreated with 20 μM U0126, a selective inhibitor of MEK1/2, for 30 min before exposure to ADK.
Antioxidant capacity
Total flavonoid content (TFC) and total phenolic content (TPC) were measured using validated methods from peer-reviewed literature 48,49 Quercetin (Sigma Aldrich) was used as the reference substance for TFC, and the results are expressed in terms of quercetin equivalents (mg QE/g). Gallic acid (Sigma Aldrich) was used as the reference substance for TPC, and the results are expressed in terms of gallic acid equivalents (μg GAE/g). Additionally, antioxidant activities were assessed through 2,2-diphenyl-1-picrylhydrazyl (DPPH) radical scavenging activity, Trolox equivalent antioxidant capacity (TEAC), and ferric-reducing antioxidant power (FRAP) according to established protocols. 50,51
Statistical analysis
All the statistical analyses were conducted using SPSS version 25.0 (Statistical Package for the Social Sciences, SPSS Inc., Chicago, IL, USA). The data are presented as means ± standard deviations (SDs). Significant differences were determined using one-way analysis of variance (ANOVA) followed by Tukey’s post hoc test for multiple comparisons. Differences were considered statistically significant at P < .05 and are indicated by an asterisk or lowercase letters above the respective bars. 48
RESULTS
Cellular toxicity of ADK extract
To determine the optimal concentration of ADK extract for pretreating ARPE-19 cells, cell viability was assessed using the MTT assay (Supplementary Fig. S1). The cells remained viable up to a concentration of 200 μg/mL ADK. However, treatment with 500 μg/mL ADK significantly decreased cell viability, resulting is a survival rate of 85% (P < .001; Supplementary Fig. S1). Consequently, the highest concentration of ADK extract that did not significantly reduce cell survival (200 μg/mL) was selected for subsequent experiments involving ARPE-19 cells.
Effects of ADK extract on Tg-induced VEGF secretion and [Ca2+]i
We investigated the effects of Tg on VEGF protein secretion and [Ca2+]i concentration in ARPE-19 cells pretreated with ADK extract (Fig. 1). VEGF secretion, a key factor in angiogenesis, was approximately 1.6 times greater in the Tg-treated group than in the control group. However, pretreatment with ADK prevented VEGF protein secretion by approximately 26–36% compared with that in the Tg-treated group (Fig. 1A). To assess whether the reduction in VEGF secretion due to pretreatment with ADK extract in response to Tg treatment is related to MEK expression, we treated ARPE-19 cells with the MEK inhibitor U0126 (20 μM). Pretreatment with U0126 led to a decrease in VEGF secretion of approximately 58% compared with VEGF secretion in the Tg-treated group, suggesting that the decrease in VEGF protein secretion induced by ADK extract is linked to the inhibition of MEK expression. There was a 2.3-fold increase in the [Ca2+]i concentration (Fig. 1B) in the Tg-treated group compared with the [Ca2+]i concentration in the control group. Pretreatment with ADK extract at concentrations of 25, 50, 100, and 200 μg/mL resulted in reductions in [Ca2+]i concentrations of 1%, 10%, 22%, and 28%, respectively, compared with the [Ca2+]i concentration in the Tg-treated group, with statistically significant reductions observed at concentrations of 100 and 200 μg/mL.

Preventive effects of ADK pretreatment on Tg-induced VEGF secretion and [Ca2+]i. ARPE-19 cells were cultured for 48 h, pretreated with ADK (0–200 μg/mL) for 3 h, and then treated with Tg (1 μM) for 24 h.
Effects of ADK extract on the Tg-induced expression of ER stress-related proteins
To assess the protective effects of ADK on ER stress induced by Tg in ARPE-19 cells, we analyzed the expression levels of ER stress-related proteins via western blot analysis (Fig. 2 and Supplementary Fig. S2). Calnexin expression increased by 1.32-fold following Tg treatment (Fig. 2 and Supplementary Fig. S2A). However, pretreatment with ADK extract at concentrations of 100 and 200 μg/mL resulted in reductions in calnexin protein expression of 20.49% and 44.56%, respectively, compared with calnexin protein expression after Tg treatment alone (Fig. 2 and Supplementary Fig. S2A). Calnexin, acting as a chaperone, plays an essential role in protein folding and quality control, ensuring that only properly folded and assembled proteins are allowed to progress through the secretory pathway. 52 Specifically, calnexin retains N-linked glycoproteins within the ER that are either unfolded or not fully assembled. 53 Similarly, Ero1-Lα expression increased by 1.39-fold with Tg treatment but decreased by 14.28% and 25.54% with 100 and 200 μg/mL ADK extract pretreatment, respectively (Fig. 2 and Supplementary Fig. S2B). Ero1-Lα is crucial in the hypoxia-inducible factor 1-driven pathway, enhancing disulfide bond formation and VEGF release under low oxygen conditions. 54
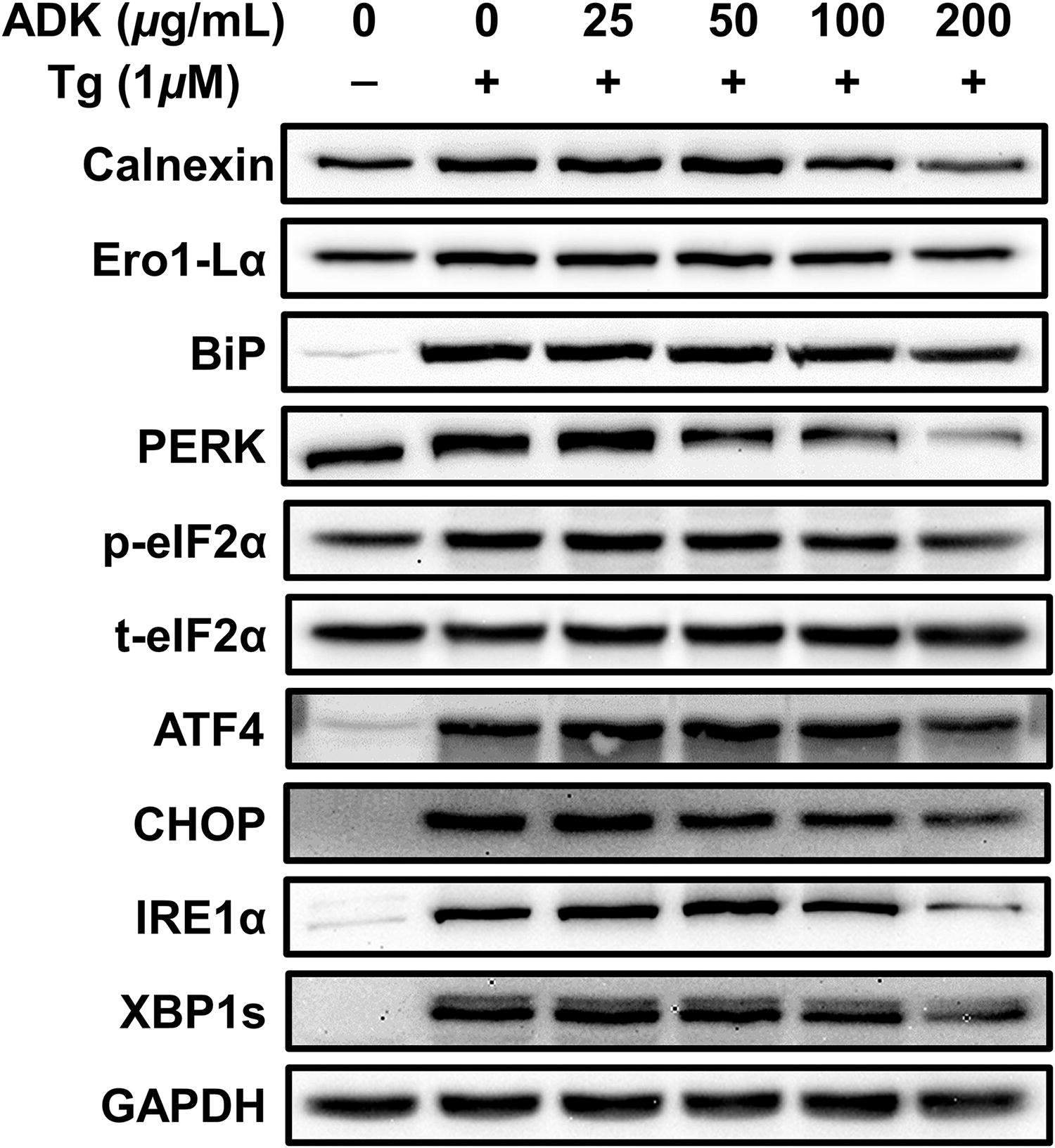
Preventive effects of ADK pretreatment against Tg-induced UPR-related protein expression. ARPE-19 cells were cultured for 48 h, pretreated with ADK (0 ∼ 200 μg/mL) for 3 h, and then treated with Tg (1 μM) for 24 h. The panels show the protein expression of calnexin, Ero1-Lα, BiP, PERK, p-eIF2α, t-eIF2α, ATF4, CHOP, IRE1α, XBP1s, and GAPDH. GAPDH was used as an internal standard for calnexin, Ero1-Lα, BiP, PERK, ATF4, CHOP, IRE1α, and XBP1s. t-eIF2α was used as an internal standard for p-eIF2α. ATF4, activating transcription factor 4; BiP, binding immunoglobulin protein; CHOP, C/EBP homologous protein; eIF2α, eukaryotic initiation factor 2 alpha; Ero1-Lα, Ero1-like protein alpha; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; IRE1α, inositol-requiring enzyme-1 alpha; p-eIF2α, phospho-eukaryotic initiation factor 2 alpha; PERK, protein kinase RNA-like endoplasmic reticulum kinase; t-eIF2α, total-eukaryotic initiation factor 2 alpha; XBP1s, spliced form of x-box binding protein 1.
Compared with that of the control, the expression of binding immunoglobulin protein (BiP), the major ER chaperone, increased 12.17-fold after Tg treatment. However, ADK extract treatment caused a concentration-dependent reduction in BiP protein expression, with significant suppression starting at a concentration of 50 μg/mL. Compared with Tg treatment alone, treatment with 200 μg/mL ADK led to a 33.83% decrease in BiP expression (Fig. 2 and Supplementary Fig. S2C). Additional research was conducted to explore whether pretreatment with ADK could reduce the protein expression of protein kinase R-like endoplasmic reticulum kinase (PERK), p-eukaryotic translation initiation factor 2 (eIF2) α, activating transcription factor 4 (ATF4), and C/EBP homologous protein (CHOP) (Fig. 2 and Supplementary Fig. S2C–G). The expression of PERK, a membrane protein of the ER that is activated by ER stress due to the presence of misfolded proteins, increased by 1.17-fold after Tg treatment. However, pretreatment with 100 and 200 μg/mL ADK resulted in reductions of 17.28% and 57.45%, respectively, with the greatest suppression observed with 200 μg/mL ADK (Fig. 2 and Supplementary Fig. S2D).
The phosphorylation of eIF2α, which was 1.59-fold greater after Tg treatment, was notably reduced by 16.45% and 37.51% after treatment with 100 and 200 μg/mL ADK extract, respectively (Fig. 2 and Supplementary Fig. S2E). eIF2α phosphorylation induced by PERK leads to ATF4 activation, which suppresses translation initiation and reduces global protein synthesis. 55,56 The level of ATF4, which plays a proapoptotic role under conditions of excessive ER stress, increased 17.28-fold in response to Tg treatment. Significant reductions were observed starting at 50 μg/mL ADK extract, with decreases of 20.05%, 25.83%, and 44.69% at concentrations of 50, 100, and 200 μg/mL ADK, respectively (Fig. 2 and Supplementary Fig. S2F). The expression of CHOP, which increased by 23.12-fold with Tg treatment, was substantially reduced starting at 50 μg/mL ADK, with reductions of 20.73%, 42.62%, and 58.65% at 50, 100, and 200 μg/mL ADK, respectively (Fig. 2 and Supplementary Fig. S2G).
ADK pretreatment significantly mitigated Tg-induced UPRs; therefore, we hypothesized that it would also inhibit the expression of inositol-requiring enzyme 1 (IRE1), an ER stress sensor and cell fate executor, 57 along with its downstream target, X-box binding protein 1 (XBP1s). IRE1α protein expression was 10.65-fold greater in Tg-treated ARPE-19 cells than in control ARPE-19 cells. However, pretreatment with ADK extract at concentrations of 50, 100, and 200 μg/mL led to notable reductions in IRE1α protein expression, with decreases of 9.83%, 25.84%, and 60.78%, respectively, compared with IRE1α protein expression after Tg treatment alone (Fig. 2 and Supplementary Fig. S2H). Additionally, XBP1s protein expression, which was 15.12-fold higher in Tg-treated ARPE-19 cells than in control cells, decreased by 12.79% and 36.48% at ADK extract concentrations of 100 and 200 μg/mL, respectively (Fig. 2 and Supplementary Fig. S2I).
Effect of ADK extract on mRNA expression alterations induced by Tg treatment
According to our previous findings, ADK pretreatment significantly prevents both UPRs and the secretion of VEGF proteins. Consequently, we hypothesized that ADK pretreatment would also inhibit the expression of ER stress-related gene expressions. To further explore the anti-ER stress effects of ADK, we examined the mRNA levels of VEGFα, the tight junction protein zonula occludens-1 (ZO-1), and the inflammatory cytokine interleukin-6 (IL-6) in ARPE-19 cells treated with Tg (Fig. 3).

Preventive effects of ADK pretreatment against Tg-induced ER stress-related mRNA expression. ARPE-19 cells were cultured for 48 h, pretreated with ADK (0–200 μg/mL) for 3 h, and then treated with Tg (1 μM) for 24 h. The graphs show the ocular gene expression of VEGFα, ZO-1, and IL-6
VEGFα mRNA expression increased by 5.97-fold in ARPE-19 cells following Tg treatment, but pretreatment with ADK extract at concentrations of 25, 50, 100, and 200 μg/mL resulted in reductions of 29%, 37%, 33%, and 30%, respectively, compared with VEGFα mRNA expression after Tg treatment alone (Fig. 3A). ZO-1 mRNA expression decreased by 24% in ARPE-19 cells following Tg treatment. Pretreatment with ADK extract led to a concentration-dependent recovery of ZO-1 mRNA expression, with levels comparable to those of the control group at pretreatment concentrations of 100 and 200 μg/mL (Fig. 3B).
Considering the expected decrease in ZO-1 mRNA expression and subsequent increase in inflammation-related gene expression, IL-6 mRNA expression was also evaluated. IL-6 mRNA expression increased by 5.01-fold in ARPE-19 cells upon Tg treatment. However, pretreatment with ADK extract at concentrations of 25, 50, 100, and 200 μg/mL led to significant reductions in IL-6 mRNA expression of 44%, 41%, 45%, and 57%, respectively, compared with IL-6 mRNA expression after Tg treatment alone (Fig. 3C).
Effects of ADK extract on the LPS-induced phosphorylation of mitogen-activated protein kinase proteins
In our previous findings, ADK pretreatment significantly mitigated Tg-induced ocular pathophysiological responses; therefore, we logically hypothesized that ADK pretreatment would also prevent ocular inflammation. Notably, the pretreatment of ARPE-19 cells with ADK suppressed the increase in the expression of inflammatory IL-6 mRNA induced by Tg, suggesting the potential effectiveness of ADK in inhibiting inflammatory responses. Consequently, we examined the changes in mitogen-activated protein kinase (MAPK) protein expression in ARPE-19 cells after LPS treatment (Fig. 3, Fig. 4).
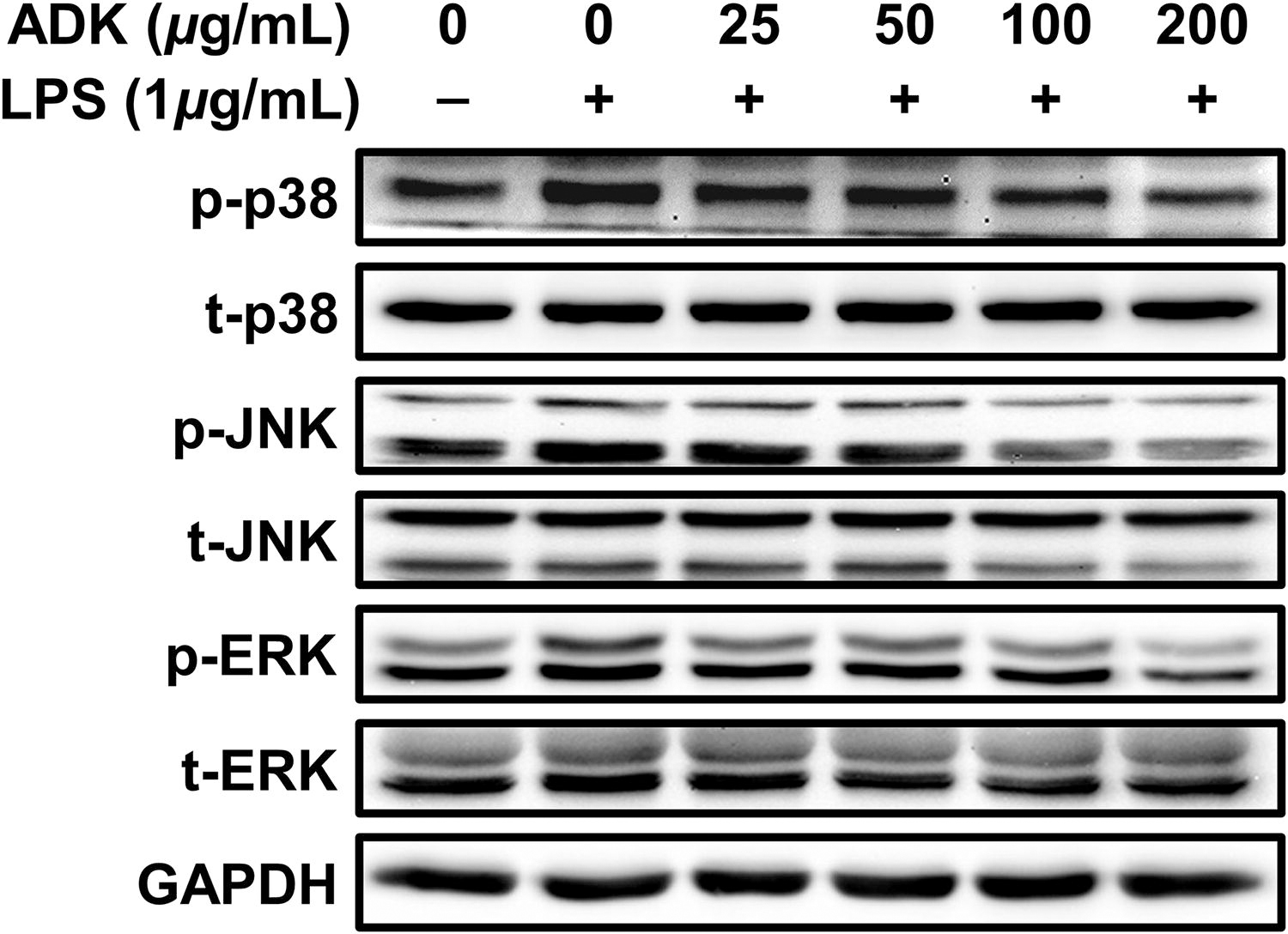
Preventive effects of ADK pretreatment against LPS-induced phosphorylation of MAPKs. ARPE-19 cells were cultured for 48 h, pretreated with ADK (0–200 μg/mL) for 3 h, and then treated with LPS (1 μg/mL) for 2 h. The panels show the protein expression of p38, JNK, and ERK. LPS, lipopolysaccharide; p-p38, phospho-p38; p-ERK, phospho-extracellular signal-regulated kinase; p-JNK, phospho-c-jun n-terminal kinase; t-p38, total-p38; t-ERK, total-extracellular signal-regulated kinase; t-JNK, total-c-jun n-terminal kinase.
A concentration-dependent trend was observed in the inhibition of MAPK protein phosphorylation upon pretreatment with the ADK extract (Fig. 4). Specifically, phosphorylation of the p38 protein increased by 1.26-fold in ARPE-19 cells after LPS treatment, whereas after pretreatment with ADK, p38 protein phosphorylation levels were 1.09-, 1.00-, 0.85-, and 0.70-fold different (a concentration-dependent decrease) than p38 protein phosphorylation levels in the control cells (Fig. 4 and Supplementary Fig. S3A). Phosphorylation of the c-jun n-terminal kinase (JNK) protein increased by up to 1.53-fold after LPS treatment, whereas after pretreatment with ADK extract, JNK protein phosphorylation levels were 1.18-, 1.02-, 0.83-, and 0.74-fold different (a concentration-dependent decrease) than JNK protein phosphorylation levels in control cells (Fig. 4 and Supplementary Fig. S3B). Similarly, phosphorylation of the extracellular signal-regulated kinase (ERK) protein increased by 1.27-fold after LPS treatment, whereas after pretreatment with ADK, ERK protein phosphorylation levels were 0.96-, 0.92-, 0.70-, and 0.37-fold lower (a concentration-dependent decrease) than ERK protein phosphorylation levels in control cells (Fig. 4 and Supplementary Fig. S3C).
Effects of ADK extract on the LPS-induced phosphorylation of IκB and nuclear factor kappa-light-chain-enhancer of activated B cells proteins
Pretreatment of ARPE-19 cells with ADK significantly inhibited the LPS-induced phosphorylation of p38, JNK, and ERK in a posttranslational manner (Fig. 4 and Supplementary Fig. S3). Consequently, follow-up experiments were performed to determine whether ADK pretreatment decreased the phosphorylation of IκB-nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB) axis. After treating ARPE-19 cells with LPS, the phosphorylation levels of the IκB and NFκB proteins increased by 2.49- and 1.28-fold, respectively (Fig. 5 and Supplementary Fig. S4A/B). However, compared with LPS treatment alone, pretreatment with ADK extract at concentrations of 50, 100, and 200 μg/mL decreased the phosphorylation of IκB by 31%, 45%, and 49%, respectively (Fig. 5 and Supplementary Fig. S4A). Additionally, the phosphorylation of NFκB decreased in a concentration-dependent manner by pretreatment with ADK extract at concentrations of 25, 50, 100, and 200 μg/mL, with NFκB phosphorylation levels 1.17-, 1.04-, 0.78-, and 0.64-fold different (a concentration-dependent decrease), respectively, than the NFκB phosphorylation levels in control cells (Fig. 5 and Supplementary Fig. S4B).
Effect of ADK extract on the LPS-induced changes in inflammatory-related gene expression
Pretreatment with ADK significantly inhibited the LPS-induced phosphorylation of components of the MAPK-NFκB axis (Fig. 4 and 5). Consequently, we investigated whether ADK extract also suppresses the expression of proinflammatory cytokine gene expressions (Fig. 6). The expression level of TNFα mRNA increased by 2.16-fold after LPS treatment in ARPE-19 cells. However, compared with the control, pretreatment with ADK extract at concentrations of 25, 50, 100, and 200 μg/mL significantly suppressed TNFα mRNA expression by 0.97-, 0.85-, 0.95-, and 0.93-fold, respectively (Fig. 6A). Similarly, IL-1β mRNA expression increased by up to 4.43-fold after LPS treatment, while pretreatment with ADK extract at concentrations of 25, 50, 100, and 200 μg/mL significantly reduced IL-1β mRNA expression by 64%, 57%, 62%, and 69%, respectively (Fig. 6B). Compared with that in LPS-treated ARPE-19 cells, IL-6 mRNA expression, which increased by 2.80-fold after LPS treatment, was suppressed in cells pretreated with ADK extract, decreasing by 39%, 30%, 29%, and 57%, respectively (Fig. 6C). Finally, monocyte chemoattractant protein 1 (MCP-1) mRNA expression increased by 4.98-fold after LPS treatment, while pretreatment with ADK extract at concentrations of 25, 50, 100, and 200 μg/mL significantly suppressed MCP-1 mRNA expression by 55%, 43%, 60%, and 74%, respectively (Fig. 6D). Thus, ADK extract significantly decreased both the phosphorylation of components of the MAPK-NFκB axis and the expression of proinflammatory cytokine genes.

Preventive effects of ADK pretreatment against LPS-induced phosphorylation of IκB-NFκB. ARPE-19 cells were cultured for 48 h, pretreated with ADK (0–200 μg/mL) for 3 h, and then treated with LPS (1 μg/mL) for 2 h. p-IκBα, phospho-nuclear factor of kappa light polypeptide gene enhancer in B cells inhibitor alpha; p-NFκB, phospho-nuclear factor kappa-light-chain-enhancer of activated B cells; t-IκBα, total-nuclear factor of kappa light polypeptide gene enhancer in B cells inhibitor alpha; t-NFκB, total-nuclear factor kappa-light-chain-enhancer of activated B cells.

Preventive effects of ADK pretreatment against LPS-inducible proinflammatory cytokines. ARPE-19 cells were cultured for 48 h, pretreated with ADK (0–200 μg/mL) for 3 h, and then treated with LPS (1 μg/mL) for 2 h. The graphs show the gene expression of the inflammatory cytokines TNFα, IL-1β, IL-6, and MCP-1
Intercellular antioxidant activity of ADK extract
ADK is known to be rich in functional materials 58 that may confer antioxidative effects. Therefore, we investigated the antioxidant activity of ADK following the treatment of cells with LPS. ARPE-19 cells were pretreated with ADK extract and then exposed to LPS, after which we examined the protein expression of heme oxygenase-1 (HO-1), which is regulated by the antioxidant response element (ARE), in cell lysates. Antioxidant activities were also assessed in the cell culture medium (Fig. 7 and Supplementary Fig. S5).
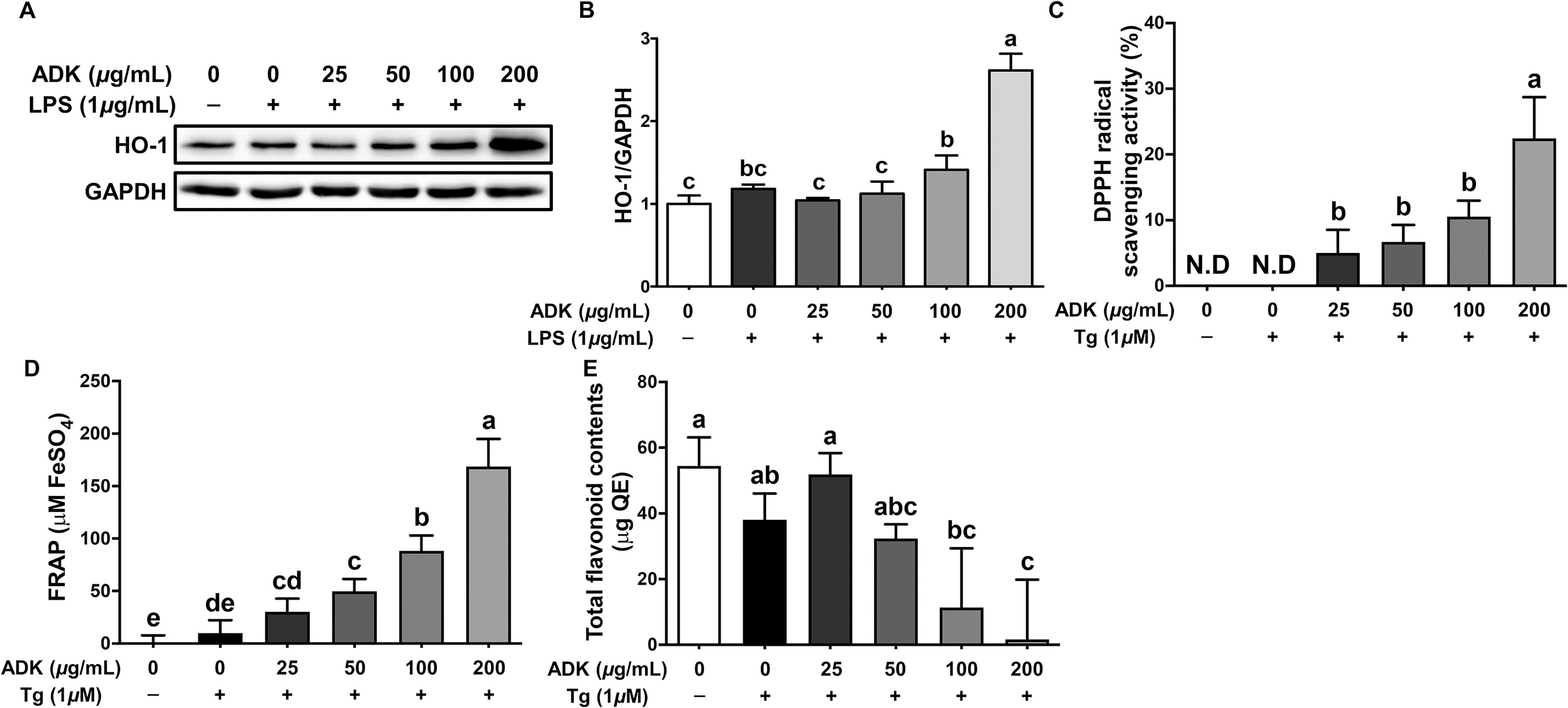
Antioxidant activity of ADK in ARPE-19 cells. ARPE-19 cells were cultured for 48 h, pretreated with ADK (0–200 μg/mL) for 3 h, treated with LPS (1 μg/mL) for 2 h or treated with Tg (1 μM) for 24 h. The graphs and panels show the protein expression of HO-1 and GAPDH
Compared with LPS treatment alone, pretreatment with ADK extract at concentrations of 100 and 200 μg/mL increased the protein expression of HO-1 by 1.19- and 2.21-fold, respectively (Fig. 7A/B). Given that this increase in HO-1 protein expression was associated with antioxidant activity, we further investigated the impact of ADK extract pretreatment on antioxidant activity in the cell culture medium. Concentration-dependent increases in DPPH radical scavenging activity were observed following pretreatment with ADK extract. Specifically, the highest DPPH radical scavenging activity, 22.4%, was observed with pretreatment with ADK extract at a concentration of 200 μg/mL (Fig. 7C). Additionally, FRAP indicated an increase in the amount of FeSO4 in a concentration-dependent manner after treatment with ADK extract, indicating greater reducing power (Fig. 7D). The TFC did not significantly differ with or without Tg treatment or pretreatment with ADK extract at concentrations of 25 and 50 μg/mL. However, a decreasing trend was observed with pretreatment with ADK extract at concentrations of 100 and 200 μg/mL (Fig. 7E), suggesting a potential association with the cellular utilization of TFC. Neither TEAC nor TPC were significantly different after Tg treatment or treatment with ADK extract (Supplementary Fig. S5A/B). Thus, it is inferred that the antioxidant activity induced by pretreatment with ADK extract in this study was less related to polyphenols and the reducing power of TEAC.
DISCUSSION
The aim of this study was to investigate the preventive effects of pretreatment with ADK extract on ER stress induced by Tg and inflammatory responses induced by LPS in ARPE-19 cells. Pretreatment with ADK significantly inhibited the Tg-induced increase in the [Ca2+]i concentration and the expression of VEGF and UPR-related proteins (calnexin, Ero1-Lα, BiP, PERK, p-eIF2α, ATF4, CHOP, IRE1α, and XBP1s). Pretreatment with ADK extract also effectively suppressed the increase in VEGFα mRNA expression and prevented the decrease in tight junction ZO-1 mRNA expression. Additionally, ADK pretreatment significantly prevented the mRNA expression of the inflammatory cytokine IL-6 mRNA expression, which was induced by ER stress due to Tg exposure. Furthermore, ADK pretreatment exhibited an anti-inflammatory effect on ARPE-19 cells by significantly preventing the phosphorylation of components of the MAPK-IκB-NFκB axis in a posttranslational manner and significantly inhibiting the mRNA expression of inflammatory cytokines (TNF-α, IL-1β, IL-6, and MCP-1) associated with NFκB activation in a transcriptional manner. Moreover, ADK pretreatment significantly increased cellular antioxidative capacity by elevating the protein levels of the antioxidant HO-1, which is associated with AREs. Additionally, increases in antioxidant activity and content were observed in the cell culture supernatant containing ADK.
Pretreatment with ADK inhibited the Tg-induced increase in UPR-related protein expression in ARPE-19 cells. Prolonged ER stress, triggered by diabetic complications, leads to neovascularization, the expression of inflammatory genes, and oxidative stress. 56,59 –61 Elevated ER stress activates UPRs, resulting in the significant upregulation of the expression of proteins such as the Ero1-Lα, calnexin, and BiP, within the ER. 30,54,62 BiP dissociates from the ER membrane-bound proteins IRE1, PERK, and ATF6 in response to ER stress, thereby activating these proteins. Activated IRE1α protein splices XBP1, ultimately activating the transcription of genes related to the downstream apoptotic pathway. 30,52 The pretreatment of ARPE-19 cells with ADK extract significantly inhibited the Tg-induced increase in the protein expression of Ero1-Lα, calnexin, and BiP. 63 Additionally, pretreatment with ADK extract dose-dependently suppressed the Tg-induced increase in the expression of IRE1α and XBP1s. 64 Tg treatment induced ER stress, leading to the increased expression of PERK pathway proteins, including PERK, p-eIF2α, ATF4, and CHOP. However, pretreatment with ADK extract effectively suppressed the expression of these proteins.
The pretreatment of ARPE-19 cells with ADK significantly suppressed the Tg-induced increase in VEGF protein secretion and [Ca2+]i. At the cellular level, Tg selectively inhibits sarco/ER Ca2+-ATPase, leading to elevated VEGF expression and [Ca2+]i. Based on a study in which linalool, a monoterpenoid compound found in ADK, was administered to Swiss albino mice and inhibited UV-B-induced VEGF expression, it can be inferred that the inhibition of VEGF protein secretion by ADK extract may be associated with the presence of linalool. 65,66 Additionally, not only linalool but also other polyphenolic compounds may significantly contribute to the inhibition of ER stress by acting as reagents for anti-ER stress. ADK pretreatment not only prevented Tg-induced VEGF secretion but also inhibited UPRs in ARPE-19 cells. Our findings suggest that ADK may function as a potential MEK inhibitor. Although VEGF expression is regulated by multiple signaling inducers, there is a known link between the activation of the MEK/ERK pathway and increased VEGF expression. Experimental observations show that U0126 treatment significantly decreased VEGF secretion, similar to the effect observed with ADK extract treatment, supporting our research hypothesis that ADK may suppress VEGF expression, at least in part, by inhibiting the ocular MEK/ERK pathway. Therefore, pretreatment with ADK extract prevented the increase in VEGF expression in Tg-treated ARPE-19 cells by partially suppressing the MEK/ERK pathway.
A logical extension of this study was to explore how ADK pretreatment affects the expression of inflammatory mRNAs (i.e., ZO-1 and IL-6) in ARPE-19 cells exposed to Tg, in conjunction with its anti-ER stress effects. Our findings clearly showed that the pretreatment of ARPE-19 cells with ADK extract significantly prevented the Tg-induced decrease in ZO-1 mRNA expression and the increase in IL-6 mRNA expression (Fig. 3B/C). Prolonged ER stress in cells is positively correlated with heightened inflammatory responses. 67 According to research by Ma et al. (2016), Tg treatment leads to a reduction in the expression of ZO-1 and occludin in ARPE-19 cells. Moreover, the decrease in tight junction proteins due to inflammation could lead to weakened cell-to-cell adhesion. 68 Therefore, it is postulated that the ER stress induced by Tg treatment in ARPE-19 cells would increase inflammatory responses, resulting in reduced ZO-1 expression. Since the pretreatment of ARPE-19 cells with ADK inhibited the Tg-induced increase in mRNA expression of the inflammatory cytokine IL-6 (Fig. 3C), it is speculated that ADK extract may suppress overall inflammatory responses.
Based on the preceding anti-ER stress effects of ADK, we hypothesized that pretreatment with ADK extract could mitigate the inflammatory responses induced by LPS treatment in ARPE-19 cells. After LPS exposure, notable increases in the posttranslational phosphorylation of MAPKs (JNK, ERK, and p38) and IκB-NFκB (Figs. 4 and 5) were observed. Other studies have also demonstrated the anti-inflammatory effects of ADK. Zhang and Kim (2014) reported the anti-inflammatory effects of ADK extract on murine macrophages. 36 Additionally, hyperoside (a quercetin derivative), a flavonoid compound found in ADK, has been reported to suppress renal inflammation via intraperitoneal administration in a diabetic nephropathy mouse model. 69 Another flavonoid compound in ADK, astragalin, has been reported to inhibit the phosphorylation of MAPKs and NFκB 70 in a posttranslational manner. Therefore, the inhibition of MAPK and IκB-NFκB phosphorylation by pretreatment with ADK extract in this study is presumed to be associated with the anti-inflammatory properties of these flavonoid components documented in previous studies. 69,70 The nuclear translocation and phosphorylation of NFκB protein induce the expression of inflammatory cytokine genes such as TNF-α, IL-6, IL-1β, and MCP-1. 71,72 In this study, upon LPS treatment of ARPE-19 cells, the mRNA expression of cytokines increased; however, pretreatment with ADK extract significantly inhibited the mRNA expression of inflammatory cytokines, suggesting that the nuclear translocation and phosphorylation of NFκB protein were suppressed (Fig. 5).
Based on various studies indicating that ADK extract possesses antioxidant activity, 73,74 we investigated whether pretreatment with ADK extract also has antioxidant effects on ARPE-19 cells (Fig. 7). Increased ER stress and inflammatory responses in cells can induce oxidative stress by promoting the excessive production of reactive oxygen species. 75 The pretreatment of ARPE-19 cells with 200 μg/mL ADK extract significantly increased the level of the HO-1 protein (Fig. 7A/B). The nuclear translocation of Nrf2 leads to the increased expression of antioxidant-related biological factors such as HO-1 and NAD(P)H quinone oxidoreductase 1 by binding with AREs. 76 –78 The pretreatment of ARPE-19 cells with ADK extract resulted in a concentration-dependent increase in DPPH radical scavenging activity and reducing power in the culture supernatant, as measured by FRAP analysis (Fig. 7C/D). TFC exhibited a significant decreasing trend with increasing concentrations of ADK extract (Fig. 7E), while TEAC and TPC did not significantly change with increasing concentrations of ADK (Supplementary Fig. S5A/B). Youn et al. (2012) reported that the ethanol extract of ADK contained 122.60 mg/g TPC and 36.80 mg/g TFC. 58 In this study, there was no difference in the TPC in the cell culture supernatant containing ADK extract, while the TFC tended to decrease. The decrease in TFC observed with increasing concentrations of ADK may be associated with an increase in the absorption and utilization rate of TFC in ARPE-19 cells; however, further in-depth studies are needed to understand the specific utilization rate and metabolism of TFC.
Our findings are captivating but have certain limitations. We used ARPE-19 cells to evaluate the effectiveness of ADK extract based on the hypothesis that ADK extract can be absorbed through the gastrointestinal tract and delivered to the retina without further modification. However, the components of ADK extract delivered to retinal cells may differ from those used in this study. After absorption and transport from small intestinal epithelial cells, ADK extract is likely metabolized into various forms. Additionally, since ADK extract consists of multiple unknown compounds, we have not fully identified which components have anti-inflammatory and ER stress effects. Furthermore, it is unclear whether the concentrations (25–200 μg/mL) of ADK extract used in this study are clinically applicable. Therefore, the results from our preliminary study might not yet support clinical trials of ADK extract due to incomplete information on its chemical composition. Despite these experimental limitations, our initial results clearly indicate a promising approach for preventing ocular disturbances through nutritional interventions with potential functional compounds. Our future research will focus on identifying the effective components within ADK extract. However, further studies are needed to understand the detailed characteristics of the absorption, distribution, metabolism, and excretion of these potential functional compounds in retinal cells.
CONCLUSION
The current study clearly demonstrated that pretreatment with ADK extract effectively prevented retinal ER stress, inflammatory responses, and oxidative stress in ARPE-19 cells. ADK extract significantly inhibited Tg-induced increases in intracellular calcium levels, VEGF secretion, and UPR expression, including calnexin, Ero1-Lα, and BiP. The presence of ADK also suppressed the expression of inflammatory cytokines such as IL-6 and enhanced cellular antioxidative capacity by elevating HO-1 protein levels. The inhibition of the MAPK-IκB-NFκB pathway and NFκB-mediated transcriptional regulation further highlighted the anti-inflammatory properties of ADK. Despite these promising results, future research is needed to identify the specific bioactive components in ADK extract and to understand their absorption, distribution, metabolism, and excretion in human retinal pigment cells.
Footnotes
AUTHORS’ CONTRIBUTIONS
Conceptualization: S.J., M.D., Y.J.K., J.-H.L., and J.-H.H. Methodology: S.J. and K.S. Data curation: S.J., M.D., K.S., Y.J.K., J.-H.L., and J.-H.H. Writing—original draft preparation: S.J., M.D., K.S., Y.J.K., J.-H.L., and J.-H.H. Writing—review and editing: S.J., M.D., K.S., Y.J.K., J.-H.L., and J.-H.H. Supervision: J.-H.L. and J.-H.H. Project administration: J.-H.L. and J.-H.H. Funding acquisition: J.-H.H. All authors have read and agreed to the published version of the article.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
No conflicts of interest to disclose.
SUPPLEMENTARY MATERIAL
Supplementary Table S1
Supplementary Table S2
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.