
Abstract
Inflammation is a crucial response to harmful stimuli, but its chronic activation contributes to various diseases, including inflammatory bowel disease, osteoarthritis, and neurological disorders. While nonsteroidal anti-inflammatory drugs are widely used as anti-inflammation drugs, their extended usage often results in severe side effects, emphasizing the need for safer alternatives. Therefore, it is of the greatest importance to identify and discover new anti-inflammatory agents that exhibit a reduced incidence of adverse side effects. This study investigates the anti-inflammatory potential of methanol extracts from eight native Vietnamese plant species. These extracts were screened for their ability to inhibit nitric oxide production and pro-inflammatory cytokine expression in lipopolysaccharides-stimulated RAW264.7 macrophages. Among the tested extracts, those derived from Huberantha luensis (Pierre) Chaowasku and Ancistrocladus tectorius (Lour.) Merr. demonstrated notable reductions in NO, TNF-α, interleukin (IL)-1β, and IL-6 levels. Further analysis revealed that these extracts are abundant in polyphenols and flavonoids, compounds recognized for their anti-inflammatory effects. Furthermore, these extracts exerted their effects by inhibiting the kappa-light-chain-enhancer of activated B cells and mitogen-activated protein kinase signaling pathways, as evidenced by reduced phosphorylation of the proteins. These results suggest that the methanol extracts obtained from H. luensis and A. tectorius possess considerable potential in paving the way towards the innovative development of new therapeutic approaches aimed at alleviating chronic inflammation.
INTRODUCTION
Inflammation is a complex physiological response that, when triggered repeatedly by pollutants or poorly regulated acute inflammation, can lead to chronic inflammation, posing a serious threat to human health. 1,2 Persistent inflammatory responses are known to contribute to a number of diseases, including inflammatory bowel disease, osteoarthritis, atopic dermatitis, chronic obstructive pulmonary disease, and various neurological disorders. 3 Nonsteroidal anti-inflammatory drugs (NSAIDs) are commonly used to manage inflammation-related conditions by inhibiting cyclooxygenase (COX)-2, thereby reducing the production of prostaglandin E2 (PGE2) synthesis. 4 However, long-term use of NSAIDs can result in severe and potentially life-threatening side effects. 5 Therefore, there is a critical need to find alternative therapies that are effective and have fewer side effects.
Lipopolysaccharides (LPS), a key structural component of Gram-negative bacteria outer membranes, activate macrophages by engaging toll-like receptor (TLR)4, thereby initiating innate immune response. 6 LPS exposure stimulates pro-inflammatory mediators, including tumor necrosis factor (TNF)-α, interleukin (IL)-1β, IL-6, and nitric oxide (NO). 7,8 In addition, LPS activates signaling pathways such as extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinases (JNK), and p38 mitogen-activated protein kinase (MAPK), which lead to the activation of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), a key transcription factor involved in inflammation. 9 Without stimulation, NF-κB is sequestered in the cytoplasm by its inhibitor, IκBα. 10 Upon LPS stimulation, the IκB kinase (IKK) complex becomes activated, leading to the phosphorylation, ubiquitination, and degradation of IκBα. 11 This process allows NF-κB to translocate into the nucleus, where it regulates the expression of genes involved in inflammatory responses. 12 Additionally, NF-κB activation is regulated by MAPKs, including JNK, p38 MAPK, and ERK. Targeting these signaling pathways presents a promising approach to managing inflammation.
The use of medicinal plants for treating various diseases is deeply rooted in traditional medicine practices worldwide. Vietnam is one of the most biodiverse countries, with over 13,500 documented species of vascular plants. 13 Although traditional Vietnamese medicine has long used a diverse range of plant species to treat diseases, the number of well-studied plant species remains limited. 14 This study reports the screening of eight native Vietnamese plant species. We tested methanol extracts from these plants and identified promising potential anti-inflammatory agents. We then investigated two promising plant extracts identified through the screening, focusing on their inhibitory effects on the NF-κB and MAPK signaling pathways.
MATERIALS AND METHODS
Preparation of methanol extracts from Vietnamese plants
Table 1 shows the list of plants studied. Methanol extracts from the leaves and branches of Vietnamese plants were supplied by the Korea Research Institute of Bioscience and Biotechnology. To produce crude extracts, the plant leaves and branches were combined with 99.9% MeOH. The resulting methanol extracts were then filtered and evaporated at 40°C under lower pressure.
List of Studied Plants
Cell culture
The RAW264.7 murine macrophage cell line was purchased from Korea Cell Line Bank (Seoul, Korea). Cells were cultured in DMEM medium supplemented with 10% Fetal Bovine Serum (Gibco, Gaithersburg, MD), and 1% penicillin/streptomycin solution (Welgene, Gyeongsan, Korea) at 37°C in an incubator with an atmosphere comprising 5% CO2.
Cell viability assay
Cell viability was assessed using the MTT assay. RAW264.7 cells were cultured on 96-well plates at a density of 1 × 104 cells/well. After 24 h incubation, the cells were treated with various concentrations of the Vietnamese plant methanol extracts for 24 h, and MTT solution (0.5 mg/mL) was added for 2 h. The supernatant was removed, and the produced formazan was solubilized in 100 µL of dimethyl sulfoxide (Sigma-Aldrich, St. Louis, MO). The absorbance of the resulting solution was assessed at a wavelength of 595 nm.
Measurement of nitric oxide production
The Griess reagent system was employed to quantitatively determine the concentration of NO produced within the cell culture medium. Cells were cultivated on 24-well plates at a density of 1 × 105 cells/well. Following a 24 h incubation, the cells were treated with LPS at a concentration of 500 ng/mL, concurrently with the application of a Vietnamese plant methanol extract at a concentration of 50 µg/mL for 24 h. After centrifuging the cell culture medium at 16,000 g for 3 min, 50 μL of a 1% sulfanilamide solution in 5% phosphoric acid was added to the supernatant. Then, 50 μL of a 0.1% N-(1-Napthyl)ethylenediamine dihydrochloride solution. The absorbance of the resulting mixture was assessed at a wavelength of 540 nm. A nitrite standard curve (0–100 μM) was used to calculate NO concentration.
Total polyphenol assay (TPC)
200 μL of Vietnamese plant methanol extract was reacted with 10 μL of 20% Na2CO3 and 10 μL of 2 N Folin-Ciocalteu solution for 30 min in the dark. The absorbance of the resulting mixture was assessed at a wavelength of 700 nm. The results were expressed as tannic acid equivalence value (TAE).
Total flavonoid content (TFC)
1 mL of Vietnamese plant methanol extract was mixed with 75 μL of 5% NaNO2 and the mixture was left in the dark. After 5 min, 150 μL of 10% aluminum chloride solution was added and incubated for 6 min. Next, 500 μL of 1 M sodium hydroxide was added into the mixture. After standing at room temperature for 15 min, the absorbance of the resulting mixture was assessed at a wavelength of 510 nm. The results were expressed as quercetin equivalent values (QE).
Real-time PCR
Total RNA isolation was executed utilizing a commercially available kit (Favorgen Biotech, Taipei, Taiwan). The Nanodrop One Microvolume UV-Vis Spectrophotometer (Thermo Fisher scientific, Waltham, MA) was employed to quantify RNA concentration, and 1 µg of total RNA was reverse transcribed into cDNA using AccuPower CycleScript RT PreMix (Bioneer, Daejeon, Korea). The quantification of TNF-α, IL-1β, and IL-6 expression levels was accomplished through real-time PCR, utilizing the PowerUp SYBR Green Master Mix (Thermo Fisher Scientific) and detection system (Applied Biosystems). The quantification of the expression levels of TNF-α, IL-1β, and IL-6 was assessed relative to the control gene GAPDH, and the relative mRNA expressions were determined by the 2-ΔΔCt method. The sequences of the primers used for real-time PCR amplification are listed in Table 2.
Primer Sequences Used for Real-Time PCR
Western blotting
For comprehensive extraction of total protein, cell lysis was achieved using radio-immunoprecipitation assay buffer for conjunction and the Pierce™ Protease and Phosphatase Inhibitor Mini Tablets, EDTA-Free (Thermo Fisher Scientific). The samples were centrifuged at 4°C and 16,000 g for 20 min to obtain the supernatant. Subsequently, an aliquot of 20 μg of protein was subjected to 10% SDS-PAGE separation and transferred to a polyvinylidene fluoride membrane. To attenuate nonspecific binding, the membrane was subjected to a blocking procedure with 5% skim milk at room temperature for 1 h. The membrane was then incubated with primary antibodies over a longer period of time overnight at 4°C. The study’s primary antibodies used were acquired from Cell Signaling Technology (Danvers, MA).
Statistical analysis
IMB SPSS software was used to perform the statistical analysis of the experimental data (version 23.0 Armonk, NY, USA: IBM Corp.). Three separate experiments were used to calculate the means ± standard deviations, which is how the data are presented. The t-test was used to assess group difference. A P value < 0.05 was accepted as statistically significant, and values were indicated as *P < .05, **P < .01, and ***P < .001.
RESULTS
Effect of methanol extracts on the viability in RAW264.7 cells
To evaluate the cytotoxic potential of the methanolic extracts of the eight plant species, cells were incubated with each extract (12.5–100 µg/mL) for 24 h. As shown in Figure 1, cytotoxicity was observed at a concentration of 100 µg/mL, leading to the use of 50 µg/mL in subsequent experiments.

Effect of Vietnamese native plant extracts on cell viability in RAW264.7 cells. Cell viability was determined by MTT assay. Samples were treated at the indicated concentration for 24 h. Values represent the mean ± SD of three experiments. *P < .05; **P < .01; ***P < .001 versus CON group.
Inhibitory effects of methanol extracts on NO production in LPS-induced RAW264.7 cells
To evaluate the inhibitory effects of the methanolic extracts from the eight plant species on LPS-induced NO production, cells were incubated with 50 µg/mL of each extract for 24 h. Notably, LPS treatment produced 14.03 ± 3.03 µM NO, and Huberantha luensis (Pierre) Chaowasku, Merremia boisiana (Gagnep.) Ooststr., Melaleuca cajuputi Roxb., Ancistrocladus tectorius (Lour.) Merr., Memecylon lilacinum Zoll. & Mor., and Diospyros venosa Wall. significantly reduced NO production compared to the LPS group. Especially, H. luensis, A. tectorius, and D. venosa showed decreases of 83.6%, 71.9%, and 68.8%, respectively, compared to the LPS group (P < .01)(Fig. 2).

Effect of Vietnamese native plant extracts on NO production in LPS-induced RAW264.7 cells. NO production was determined by Griess assay. Samples were treated at a concentration of 50 µg/mL and LPS at a concentration of 500 ng/mL for 24 h. Values represent the mean ± SD of three experiments. *P < .05; **P < .01 versus LPS group, ##P < .01 versus CON group. LPS, lipopolysaccharide.
Effects of methanol extracts on pro-inflammatory cytokine production in LPS-induced RAW264.7 cells
To further investigate the potential regulatory effects of methanol extracts from the eight plants on LPS-induced inflammatory gene expression, the gene expressions of pro-inflammatory cytokines TNF-α, IL-1β, and IL-6 were examined. As shown in Figure 3, H. luensis and A. tectorius significantly reduced TNF-α and IL-6 expression compared to the LPS group. In addition, methanolic extracts from seven plants, excluding Scleropyrum wallichianum var. mekongense (Gagnep.) Lecomte, significantly downregulated IL-1β expression. Based on their pronounced inhibitory effects on NO production and pro-inflammatory cytokine levels, the extracts of H. luensis (HME) and A. tectorius (AME) were selected for further investigation.

Effect of Vietnamese native plant extracts on TNF-α, IL-1β, and IL-6 mRNA expression in LPS-induced RAW264.7 cells. The mRNA expression was determined by real-time PCR. Samples were treated at a concentration of 50 µg/mL for 24 h and LPS (500 ng/mL) were treated for 3 h. Values represent the mean ± SD of three experiments. *P < .05; **P < .01; ***P < .001 versus LPS group. LPS, lipopolysaccharide.
Phytochemical contents in HME and AME
The TPC and TFC of HME and AME were assessed using colorimetric methods. According to Table 3, the TPC and TFC of HME were determined to be 339.52 ± 0.48 µg TAE/mg sample and 380.37 ± 8.98 µg QE/mg sample, respectively. Additionally, the TPC and TFC of AME were found to be 451.77 ± 4.11 µg TAE/mg sample and 906.30 ± 42.53 µg QE/mg sample, respectively.
Total Polyphenol Content and Total Flavonoid Content of HME and AME
Effect of HME and AME on iNOS and COX-2 expressions in LPS-induced RAW264.7 cells
iNOS and COX-2 are involved in the synthesis of the inflammatory mediators NO and PGE2, respectively. The investigation of iNOS and COX-2 protein expression revealed that both HME and AME inhibited iNOS expression in a dose-dependent manner, while COX-2 expression was not affected (Fig. 4).

Effect of HME and AME on the expression of inflammatory mediator proteins in LPS-induced RAW264.7 cells. Samples were treated with the indicated concentration for 24 h and LPS (500 ng/mL) were treated for 24 h. After harvesting, iNOS and COX-2 protein expression was examined by western blotting. β-actin was used as a loading control. Values represent the mean ± SD of three experiments. *P < .05; **P < .01; ***P < .001 versus LPS group.
Effect of HME and AME on the NF-κB and MAPKs pathways in LPS-induced RAW264.7 cells
Phosphorylation of the IκB and NF-κB p65 represents a crucial event in the transcriptional activation and subsequent translocation of NF-κB. Without stimulation, NF-κB is sequestered in the cytoplasm due to its binding to IκB. 15 After stimulation with LPS, IκB undergoes phosphorylation and is subsequently degraded by IκB kinase, facilitating translocation of the released NF-κB into the nucleus. Consequently, we assessed NF-κB and IκBα proteins after treatment with HME and AME. Our results showed that the phosphorylation of p65 was decreased in LPS-induced RAW264.7 cells upon treatment with HME and AME (Fig. 5B). LPS treatment triggered the degradation of IκBα, a process that was significantly inhibited by treatment with HME and AME (Fig. 5C). It was observed that p-IκBα levels were increased after LPS administration. However, this increase was significantly inhibited by HME and AME (Fig. 5D). Furthermore, significant reductions in the expression levels of ERK and JNK proteins were detected compared to the LPS group (Fig. 5E–F).
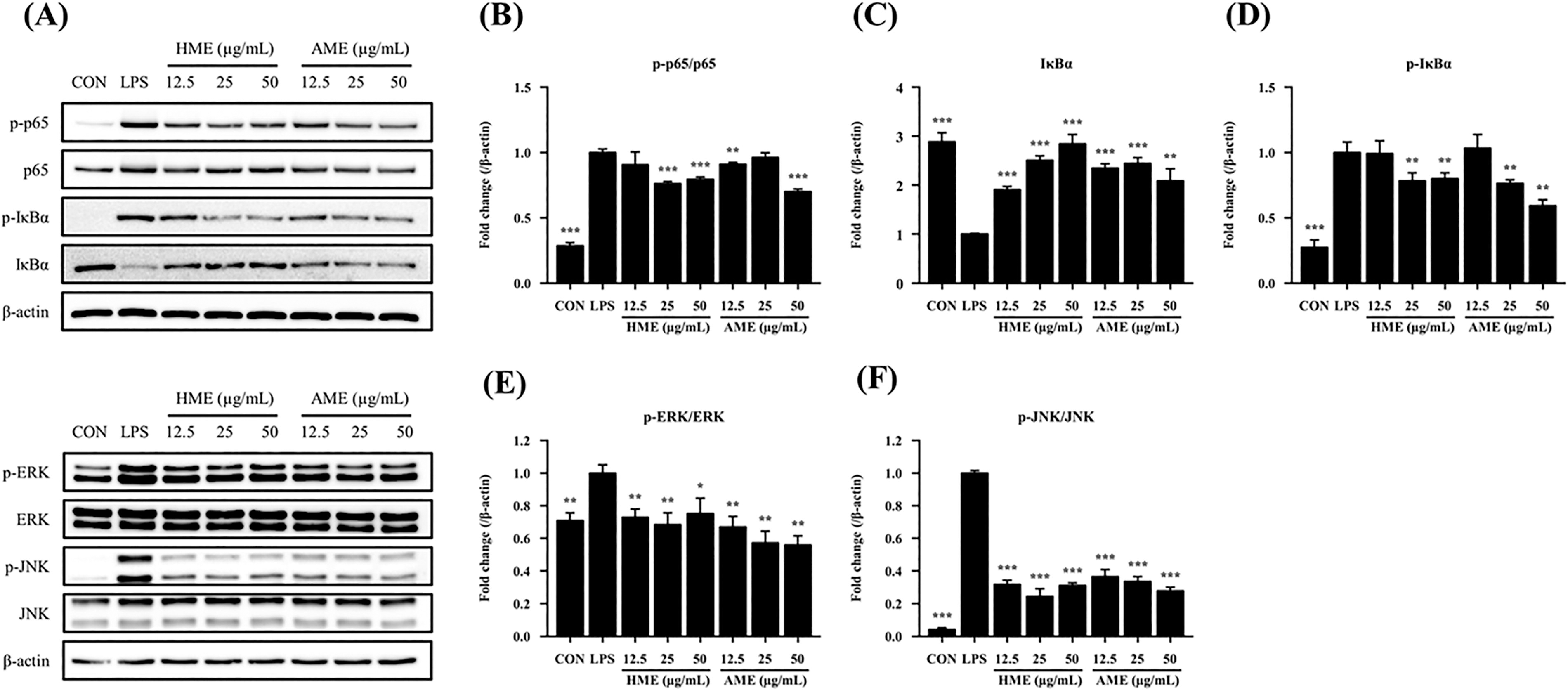
Effect of HME and AME on the expression of NF-κB and MAPK signaling pathway proteins in LPS-induced RAW264.7 cells.
DISCUSSION
Inflammation is a multifaceted biological response activated by various stimuli, including bacterial infections and environmental factors. 16 During inflammation progression, activation of macrophages can result in the secretion of various inflammatory mediators and cytokines. 17 Excessive expression of these inflammatory factors, such as NO, TNF-α, IL-1β, and IL-6, can trigger cellular inflammatory responses that lead to tissue damage. 18,19 These values are often considered indicators of the severity of inflammation. 20 Therefore, anti-inflammatory agents typically inhibit the excessive production of these mediators.
We first screened the inhibitory effect of eight native Vietnamese plants on the production of NO, TNF-α, IL-1β, and IL-6 in LPS-stimulated macrophages. NO is an important inflammatory mediator produced through the catalytic action of iNOS. 21 NO plays an essential role in the immune system, and its overproduction can lead to various inflammatory diseases. 22 In addition, TNF-α, IL-1β, and IL-6 are important inflammatory markers that are often present in elevated levels of inflammatory diseases. 23 These cytokines play a pivotal role in the inflammatory cascade by stimulating the synthesis of additional inflammatory mediators, facilitating immune responses, and ultimately resulting in cellular apoptosis and tissue injury. 24,25 Figures 2 and 3 demonstrate that HME and AME significantly suppressed the excessive production of NO, TNF-α, IL-1β, and IL-6 at a concentration of 50 µg/mL and significantly reduced TNF-α and IL-6 among the eight Vietnamese plants. This indicates that HME and AME might possess anti-inflammatory properties through the reduction of these inflammatory factors.
Polyphenols, which are abundantly present in plants, contribute to pharmacological activities of bioactive compounds. 26,27 They are classified into flavonoids and non-flavonoids based on their chemical structure. 28 Flavonoids, a major class of polyphenols commonly consumed in the human diet, exhibit significant anti-inflammatory activity. 29 The ability of these natural compounds to modulate the expression of inflammatory factors such as iNOS and cyclooxygenase contributes to the regulation of inflammatory signaling. 30 iNOS, an enzyme responsible for NO synthesis, is a crucial marker for detecting inflammatory responses. 31,32 The regulation of iNOS expression is considered an important target for treating inflammatory diseases. 33 COX enzymes are crucial in inflammatory response, with COX-2 acting as an inducible enzyme that catalyzes the conversion of arachidonic acid to prostaglandins and plays a key role in inflammation. 34 Therefore, reducing or inhibiting COX-2 activity is an effective strategy to control inflammation. The significant total content of polyphenols and flavonoids observed in HME and AME (Table 3) and the markedly lower iNOS expression in LPS-treated cells following HME and AME treatment (Fig. 4) suggest that these polyphenolic compounds may confer anti-inflammatory effects by downregulating iNOS protein expression.
NF-κB is a nuclear transcription factor that is integral to the regulation of transcription in inflammation. 35 Under non-inflammatory conditions, NF-κB is typically sequestered in the cytoplasm through its association with IκBα. 32 Upon stimulation by LPS, IκBα is phosphorylated by upstream kinases such as IKKβ, leading to its degradation and subsequent release of NF-κB, which translocates to the nucleus and further exacerbates inflammation by promoting the release of inflammatory cytokines, including TNF-α, IL-1β, and IL-6. 12,17,36 Thus, the phosphorylation levels of IκBα and NF-κB p65 are crucial for the transcriptional activation of genes encoding inflammatory mediators and cytokines. 37 The findings of our study indicate that the administration of LPS led to a marked increase in the phosphorylated form of p-IκBα, while concurrently resulting in a notable decrease in total IκBα when compared to the control group. This suggests that LPS stimulation markedly enhances the phosphorylation and subsequent degradation of IκBα (Fig. 5C–D). In contrast, treatment with HME and AME effectively inhibited both the phosphorylation and degradation of IκBα, as well as significantly attenuated the LPS-induced phosphorylation of p65.
In addition to NF-κB, anti-inflammatory therapy can target the MAPK signaling pathway, which is another important regulator of LPS-induced inflammatory responses in macrophages. The MAPK family consists of three subgroups, which are p38, JNK, and ERK. 38 Upon inflammatory stimulation, MAPKs are phosphorylated, leading to the phosphorylation and activation of various intracellular targets. 39 According to our findings, ERK and JNK phosphorylation were increased by LPS treatment compared to the control group, whereas phosphorylation was decreased by HME and AME treatment (Fig. 5E–F). These results suggest that HME and AME may have anti-inflammatory effects by inhibiting both NF-κB and MAPK signaling pathways.
In summary, our study supports the potential use of HME and AME as alternative or complementary therapies for inflammatory diseases and provides a basis for the development of more effective anti-inflammatory agents.
Footnotes
AUTHORS’ CONTRIBUTIONS
S.J.O.: Methodology, investigation, writing—original draft, visualization. Y.S.J.: Conceptualization, methodology, validation, formal analysis. E.-R.K., J.I.K., H.K., Y.K., C.W.P., and H.J.: Investigation. T.B.T. and T.T.N.: Resources. I.-K.H.: Writing—review and editing, supervision, and project administration.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This study was supported by the KRIBB Initiative Program of the Republic of Korea (KGM1172511) and project ĐTĐL.CN-72/22 in Vietnam.