
Abstract
Skeletal muscle health is essential for metabolic homeostasis, and dysregulated apoptosis in muscle cells can lead to muscle wasting and degenerative diseases. Although soybean-derived peptides are known for their bioactive properties, including antioxidant and anti-apoptotic effects, their impact on apoptosis regulation in skeletal muscle cells remains underexplored. This study aims to investigate the effects of low-molecular-weight soy peptide hydrolysate (SPH) on apoptosis and related markers in C2C12 myoblasts. SPH was prepared by enzymatic hydrolysis, and its antioxidant and anti-apoptotic activities were analyzed through in vitro assays (i.e., ABTS, DPPH). Cellular studies were used to evaluate SPH effects on ROS scavenging and apoptosis. Results show that SPH enhanced antioxidant activity, reduced ROS levels, and promoted cell proliferation in a dose-dependent manner. SPH notably decreased apoptosis under oxidative stress by down-regulating p53, c-Caspase-3, and Cyto-c, while promoting HO-1 expression, likely via AMPK activation. Importantly, SPH had no significant impact on inflammation-related proteins or MAPK activation. These findings suggest that SPH may have therapeutic potential against oxidative stress and apoptosis in muscle cells through intrinsic pathways, contributing to muscle health maintenance.
Skeletal muscle plays a fundamental role in metabolic homeostasis. The balance between cell survival and apoptosis in skeletal muscle is pivotal for its function. Dysregulated apoptosis in muscle cells can lead to pathological conditions, including muscle wasting, 1,2 degenerative diseases. 3 Understanding the molecular mechanisms underlying apoptosis in skeletal muscle is therefore crucial for advancing our knowledge of muscle physiology and the development of therapeutic interventions.
Soybean-derived functional peptides have garnered considerable attention for their potential health-promoting properties. 4 –6 These peptides, derived from enzymatic hydrolysis of soy proteins, exhibit a wide range of bioactive functions, including antioxidant, 7 anti-inflammatory, 8 and anti-apoptotic 9 effects. Their bioactivity is attributed to specific peptide sequences that interact with cellular signaling pathways, 9 making them promising candidates for nutritional and pharmaceutical applications. While there is existing evidence supporting the beneficial effects of soy-derived peptides in various cellular contexts, a comprehensive investigation into their impact on apoptosis and related signaling pathways in skeletal muscle cells is notably lacking.
The current literature lacks a comprehensive understanding of how low-molecular-weight soy peptide hydrolysate (SPH) modulates apoptosis in skeletal muscle cells. Given the importance of apoptosis in muscle physiology and the bioactive potential of soy peptides, this study aims to address this gap. Specifically, it investigates the effects of SPH on ROS scavenging, antioxidant activity, and apoptotic pathways in C2C12 myoblasts, offering insights into its potential therapeutic applications for skeletal muscle dysfunction.
A commercial isolated soy peptide (ISP, Beijing DuPont Co., Ltd., Beijing, China) diluted in 100 g/L distilled water was incubated with Alcalase (enzyme activity 2.4 AU/g, Novozymes, Bagsvaerd, Denmark) of 0.5% (w/w) at pH 8.5, 60°C for 24 h. Reaction products were filtered using filter paper (Whatman, 1001–0155), and low molecular weight fractions were separated using an ultrafiltration membrane with a molecular weight cut-off of 10 kDa (Millipore, Darmstadt, Germany). It was concentrated using an evaporator (Rotavapor R-210, BÜCHI Labortechnik, Flawil, Switzerland) and freeze-dried. SPH was provided by S Bio Co., Ltd. (Cheongju, Korea) and used as a test sample. Antioxidant activity was examined via in vitro assays and using the C2C12 murine myoblast cell line. In addition, expressions of apoptotic protein and related signaling molecules were measured using Western blotting. Detailed information for materials and methods is provided in Supplementary Data S1.
In the present study, effects of low-molecular-weight (<1.5 kDa) SPH on apoptosis and related markers were examined. ISP and SPH were compared throughout the study. First, the effects of nontoxic concentrations of each sample on ROS scavenging effect and cellular antioxidant activity were tested (Supplementary Table S1; Fig. 1). Both ISP and SPH showed dose-dependent radical scavenging effects measured by the ABTS assay (Fig. 1A), but there was no effect in the DPPH assay (Supplementary Table S1). The discrepancy in radical scavenging activity between ISP and SPH in the DPPH and ABTS assays stems from differences in assay conditions. The hydrophobic DPPH radical in an organic medium interacts poorly with hydrophilic peptides, while the ABTS radical, soluble in both aqueous and organic media, interacts more effectively. 10 Before the cellular antioxidant activity assay, appropriate concentrations of H2O2 and the effects of ISP and SPH on cell viability were determined. A cytotoxicity test using 200, 400, 600, 800, and 1000 μM of H2O2 showed 88%, 71%, 61%, 56%, and 45% cell viability, respectively (Supplementary Fig. S1). Based on these results, 600 μM was used for cell viability and antioxidant assays, whereas 300 μM was used for other protein markers. Higher oxidative stress (600 μM) was applied to assess the protective effects of ISP and SPH more clearly, while moderate stress (300 μM) allowed for specific evaluation of apoptosis pathways, minimizing necrosis or nonspecific cell death that could interfere with protein expression analysis. All concentrations of SPH (0.4, 0.6, 0.8, and 1 mg/mL) stimulated cell proliferation (Fig. 1B). In cellular antioxidant activity, 0.2, 0.4, and 0.6 mg/mL of SPH decreased cellular ROS levels in C2C12 cells against AAPH. The ROS level of 0.8 mg/mL was not altered, and 1 mg/mL of SPH showed prooxidant activity, which might be due to stimulated cell proliferation (Fig. 1C). However, none of the ISP significantly changed cellular ROS level (Fig. 1C). Thus, the highest dose showing cellular antioxidant activity (i.e., 0.6 mg/mL) was utilized for further analyses. Based on the fact that cell proliferation was increased, the anti-apoptosis effect of SPH was examined in C2C12 myoblasts; SPH significantly suppressed hydrogen peroxide-induced apoptosis, which was assessed by cPARP (Fig. 2A and B). Since p53 plays a pivotal role in apoptosis by orchestrating the expression of pro-apoptotic and anti-apoptotic genes, regulating the balance between survival and cell death in response to cellular stresses, 11 first, p53 and p-p53 expressions were examined. As expected, protein expression of p53 was notably down-regulated in SPH-treated cells and p-p53 showed a trend of decrease (Fig. 2A and C). Subsequently, representative proteins involved in intrinsic and extrinsic apoptosis were measured. Fas/FADD related signaling was not altered (Fig. 2A and D). There was no change in active Caspase-8, but c-Caspase-3 was decreased despite the marginal significance (Fig. 2A and E), indicating that SPH may affect the intrinsic apoptosis pathway. Although change in Bax was not significant, Cyto-c (P = .08) was down-regulated in SPH-treated cells (Fig. 2A and F). One of the important pro-apoptotic genes upregulated by p53 is Bax that is a member of the Bcl-2 family, thereby disrupting the mitochondrial membrane leading to the release of cytochrome c into the cytosol. 12 It is thought that anti-apoptosis by SPH might be mediated by other Bcl-2 family members besides regulating Bax (e.g., PUMA, Bcl-2), which are also well known to contribute to apoptosis. 13 Thus, SPH is likely suppressing apoptosis via the intrinsic pathway despite the null effect on Bax. The anti-apoptosis effect of SPH is consistent with the promoting effect of SPH on cell proliferation shown in Figure 1B.
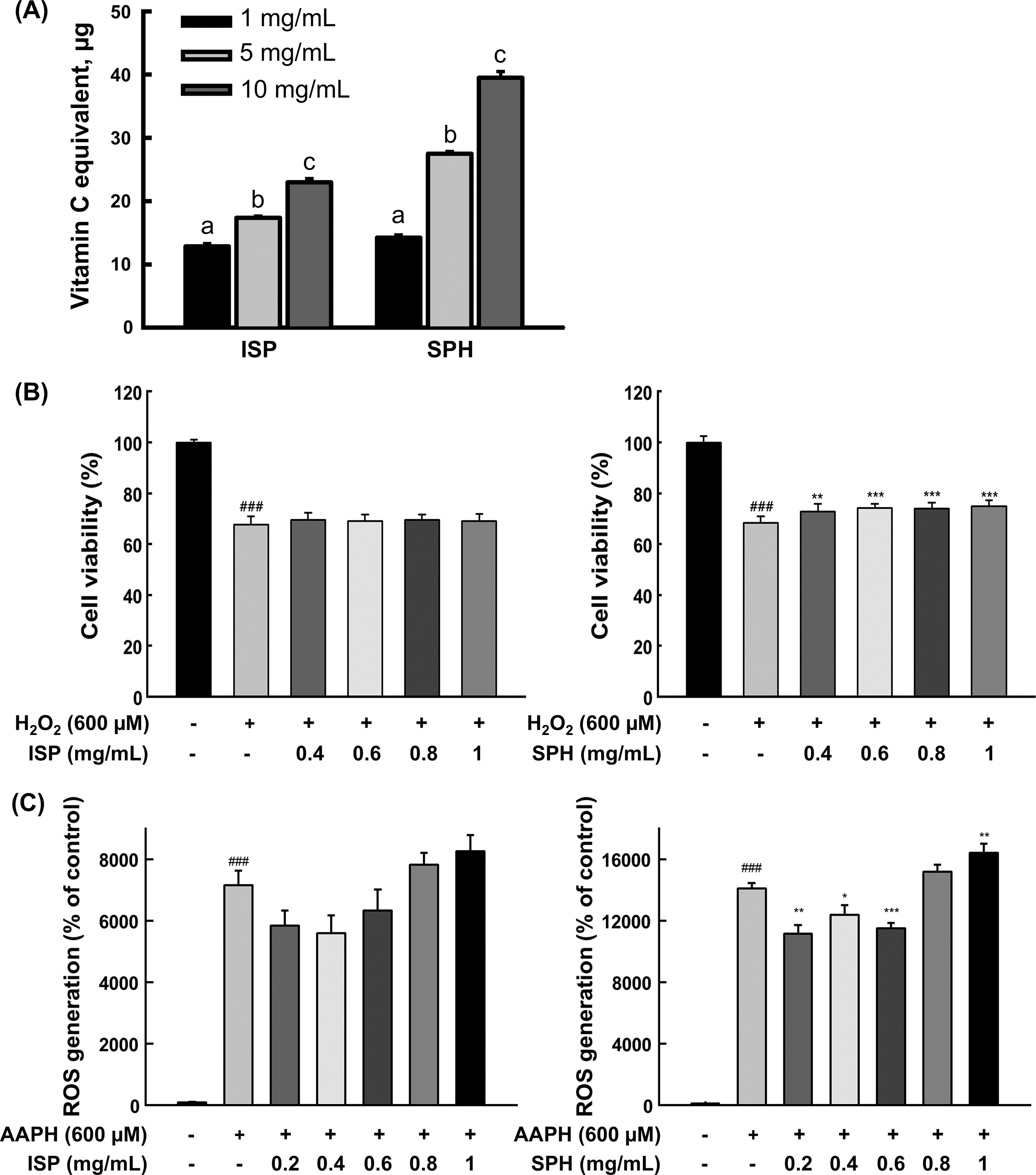
Antioxidant effect of ISP and SPH.
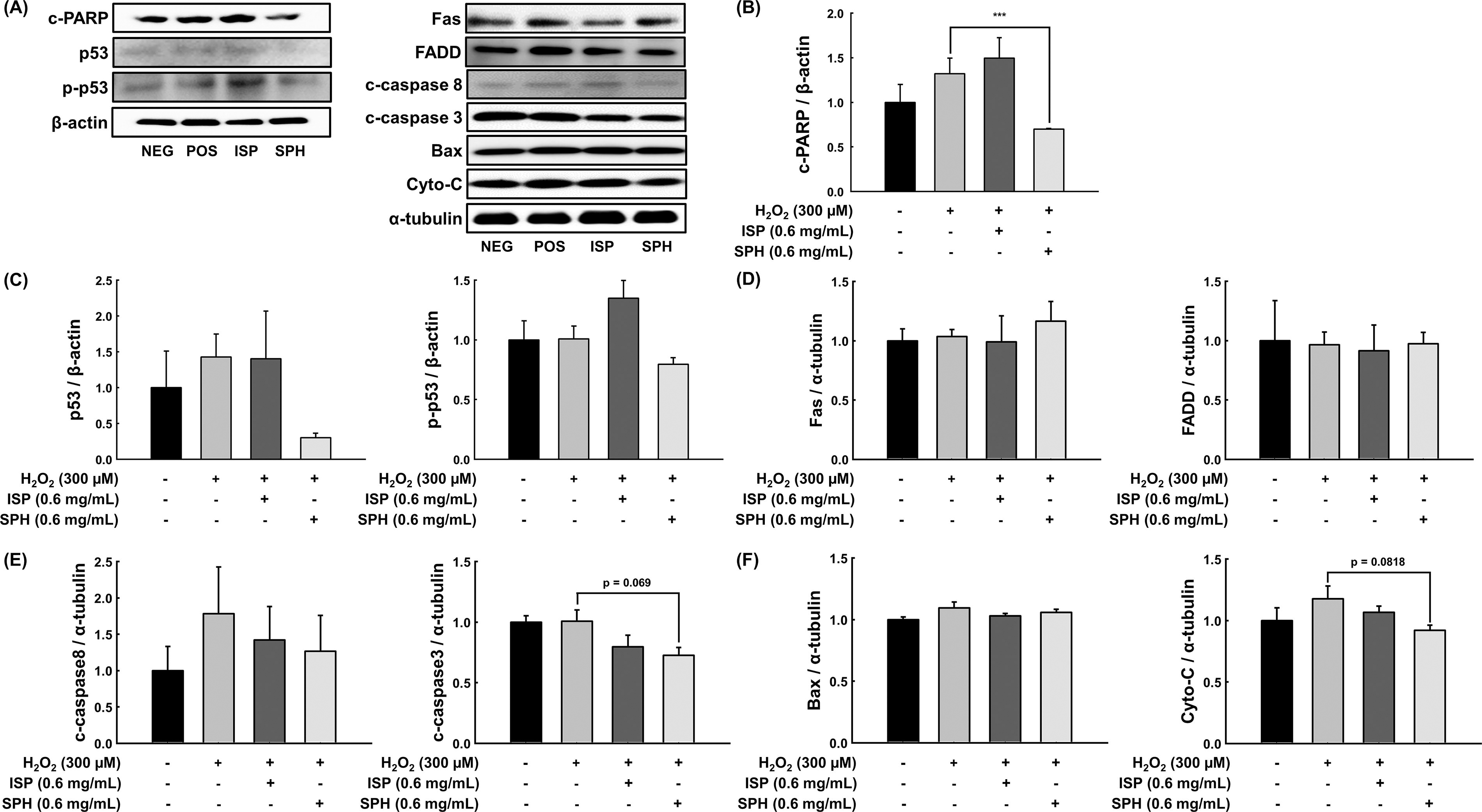
Effect of ISP and SPH on apoptosis-related markers
ROS is one of the main triggers of p53-mediated apoptosis. Activation of phase II antioxidant enzymes is a protective mechanism in response to ROS-induced oxidative damages, and HO-1 is one of the enzymes. 14,15 Hydrogen peroxide increased HO-1 expression, and SPH further elevated HO-1 in C2C12 myoblast (Fig. 3). The increased HO-1 in the hydrogen peroxide group compared with the negative control is likely a cellular defensive response to ROS. Next, we questioned how SPH increased HO-1. Of the multiple factors stimulating HO-1 gene expression, a relationship between HO-1 and AMPK is noteworthy. Multiple studies have demonstrated that activated AMPK stimulates the Nrf2/HO-1 signaling axis, specifically HO-1. 16,17 In that regard, p-AMPK and AMPK protein expression levels were examined in C2C12 cells. SPH was treated with C2C12 myoblasts while differentiating, and p-AMPK/AMPK was remarkably increased, whereas ISP did not change it (Fig. 4). It is possible that low molecular weight peptide in SPH may have an AMPK agonistic effect. In fact, there is evidence that low molecular weight (300–500 Da) peptides derived from soybean act as agonists of AMPK. 18 It should be noted that HO-1 expression and AMPK activation studies were carried out in different experimental conditions. HO-1 expression and AMPK activation were assessed under different experimental conditions. HO-1 expression in H2O2-treated myoblasts evaluated the protective role of SPH against oxidative stress by mitigating damage and upregulating HO-1. Conversely, AMPK activation in myotubes without H2O2 examined whether SPH directly activates AMPK in differentiated, metabolically active cells, independent of oxidative stress. While the lack of a direct mechanistic link remains a limitation of this study, our findings suggest that SPH-induced AMPK activation may prime cells for oxidative stress defense (e.g., HO-1 induction), warranting further investigation.
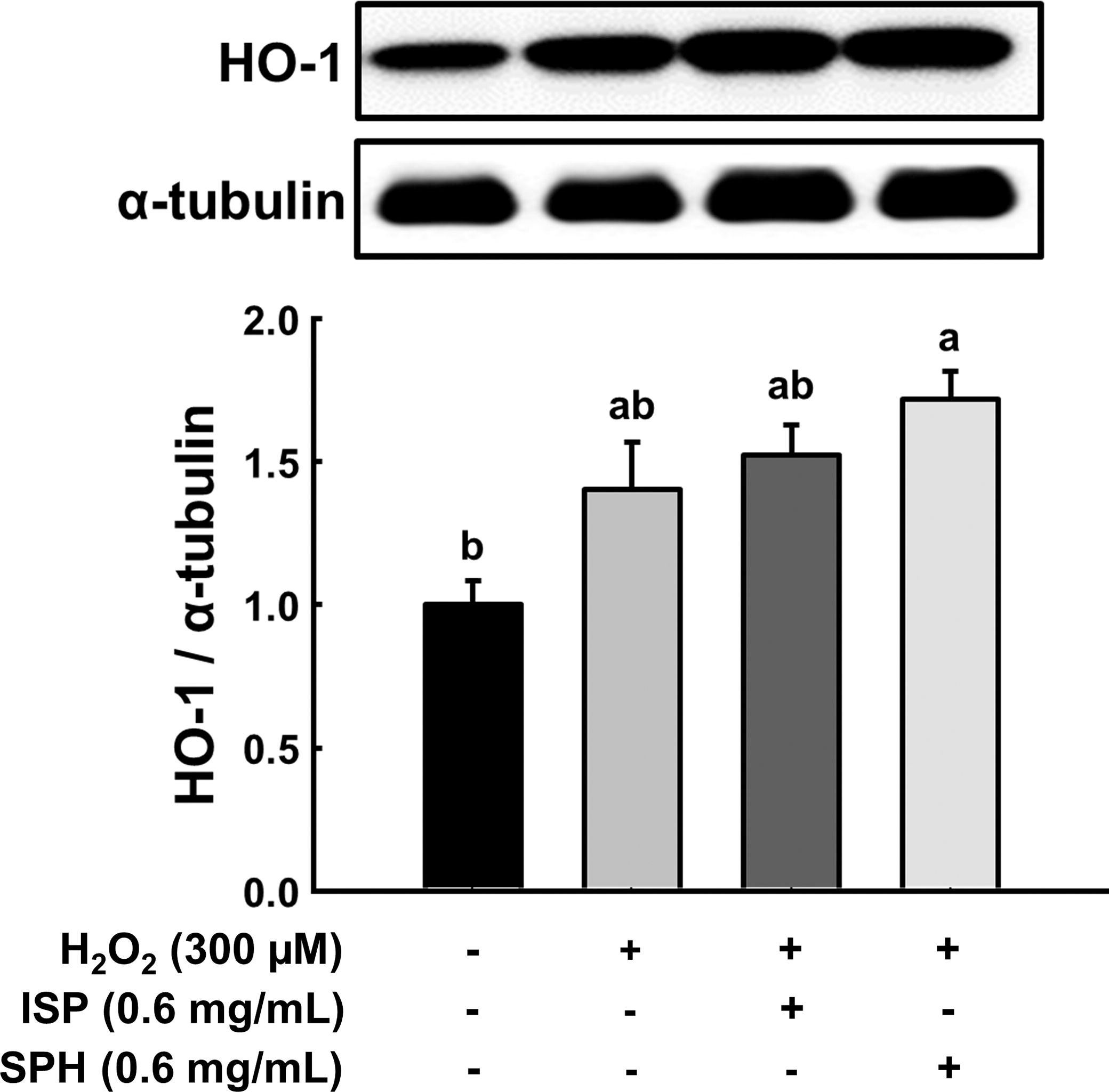
Effect of ISP and SPH on phase II antioxidant enzyme against H2O2. Representative band images from immunoblot analysis of HO-1 and their quantification. Different letters indicate statistical differences at P < .05. H2O2, hydrogen peroxide; ISP, isolated soy peptide; SPH, soy peptide hydrolysate.
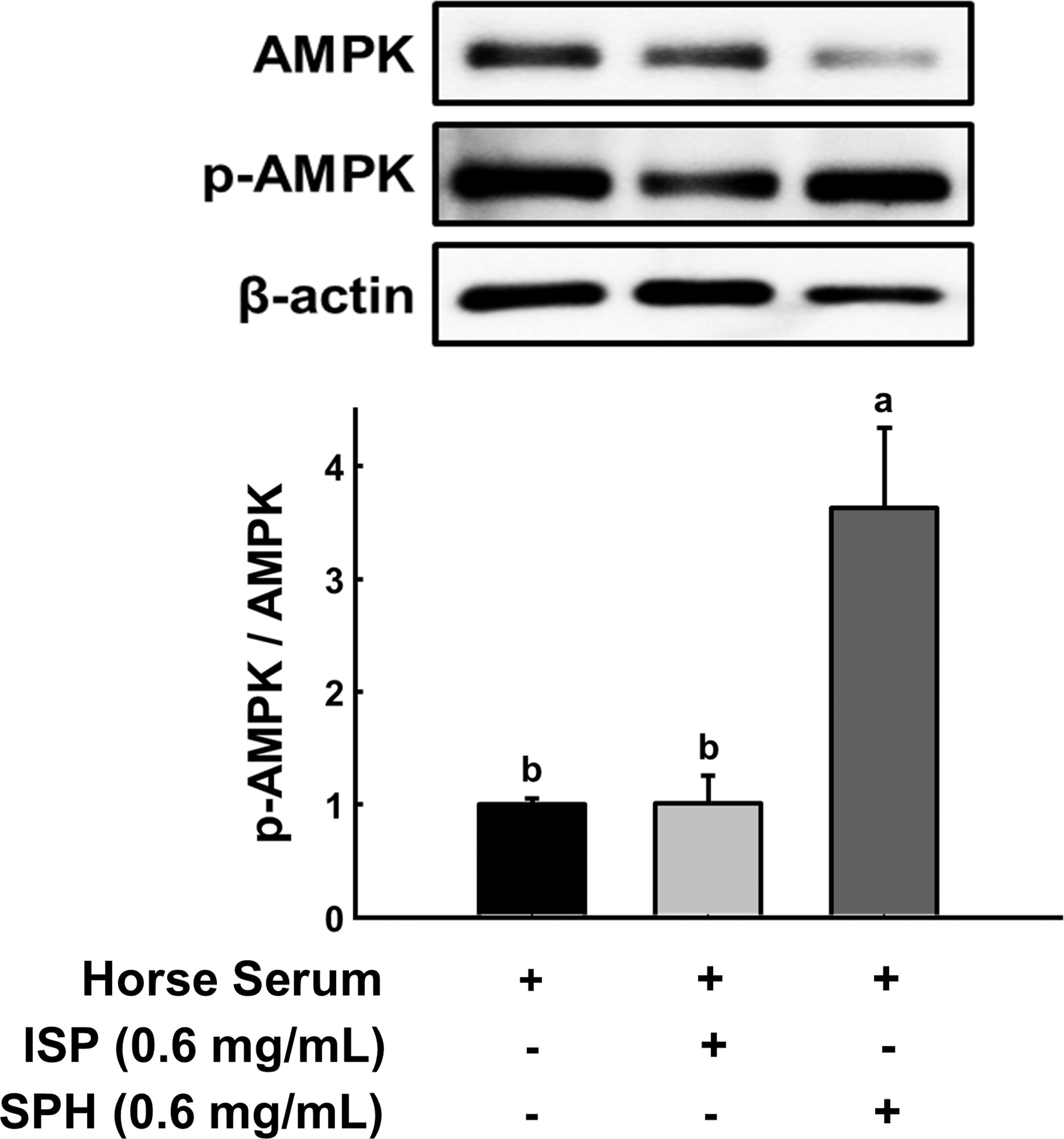
Effect of ISP and SPH on AMPK during differentiation to myotube. Representative band images from immunoblot analysis of total AMPK and p-AMPK and their quantification. Different letters indicate statistical differences at P < .05. ISP, isolated soy peptide; SPH, soy peptide hydrolysate.
Apoptosis can also be impacted by inflammation. It is well established that the ROS-mediated MAPK/NF-κB axis induces inflammation via multiple factors (e.g., adhesion molecules, inflammatory cytokines). 19 Related, key inflammatory cytokines (i.e., COX-2, p65 NF-κB, IL-1β, TNF-α) were also assessed (Supplementary Fig. S2). In addition, MAPK activation (i.e., JNK, ERK, p38) was examined (Supplementary Fig. S3). However, neither ISP nor SPH changed any of the proteins except for some marginal decrease in p-JNK, p-p65, and IL-1β.
In conclusion, this study demonstrates that low-molecular-weight SPH significantly influence cellular responses, particularly apoptosis and related mechanisms. SPH showed a dose-dependent radical scavenging effect, enhanced cellular antioxidant activity, and promoted cell proliferation at nontoxic concentrations. SPH exhibited anti-apoptotic effects by suppressing H2O2-induced apoptosis, downregulating p53, and reducing Cyto-c and c-Caspase-3 levels, though Bax expression was unaffected. In addition, SPH increased HO-1 expression, likely through AMPK activation, contributing to protection against ROS-induced damage. However, SPH had minimal effects on inflammation-related proteins and MAPK activation. While the mechanistic link between SPH and AMPK activation remains unclear, further investigation is warranted. These findings suggest SPH as promising bioactive compounds with antioxidant and anti-apoptotic properties, with potential applications in muscle disease models involving the AMPK/HO-1 axis-mediated apoptosis.
Footnotes
AUTHORS’ CONTRIBUTIONS
J.H.P.: Conceptualization, validation, writing—review and Editing, Supervision; W.H.: Methodology, formal analysis, data curation, writing—original draft, Visualization; W.H. and J.H.P.: Investigation, resources.
AUTHOR DISCLOSURE STATEMENT
The authors have declared no conflicts of interest.
FUNDING INFORMATION
This study was supported by research fund from
SUPPLEMENTARY MATERIAL
Supplementary Data S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.