
Abstract
Isoliquiritigenin (ISL), a bioactive phytochemical derived from the root of Glycyrrhiza uralensis, is known to exert anticancer effects by modulating cancer cell functions, including proliferation, metastasis, and autophagy. However, the anticancer effects of ISL on cholangiocarcinoma (CCA) are limited. In this study, we investigated the anticancer effects of ISL on CCA cells (SNU-478 and HuCCT-1). In SNU-478 cells, ISL decreased the expression of glucose-regulated protein 78 (GRP78). ISL was also found to significantly reduce cell viability and colony area of both SNU-478 and HuCCT-1 cells. Furthermore, ISL treatment significantly increased the population of apoptotic cells, as observed through Annexin V/propidium iodide (PI) staining. PI staining showed that the ISL treatment induced significant cell cycle arrest compared to the control group, indicating that ISL-induced decreases in cell viability are associated with cell cycle arrest and apoptosis. ISL treatment also resulted in a significant decline in the mitochondrial membrane potential when observed through JC-10 staining. Mitochondrial superoxide staining revealed that ISL significantly upregulated the generation of mitochondrial reactive oxygen species (ROS). Acridine orange staining indicated that the ISL treatment upregulated lysosomal membrane permeability, which could be related to autophagy. These results suggest that ISL affects the proliferation and apoptosis of CCA cells, which might be related to its regulatory role in various cellular functions, including mitochondrial dysfunction, ROS balance, and autophagy. This study provides empirical evidence that ISL could be a potential candidate for CCA therapy.
INTRODUCTION
Cholangiocarcinoma (CCA) is a primary hepatobiliary cancer that may arise anywhere in the biliary tract. 1 CCA is commonly diagnosed at an advanced stage of cancer progression because it does not cause any noticeable symptoms in the early stages. Late diagnosis reduces access to effective treatment, leading to a poor prognosis and treatment outcome. 2 The incidence of CCA has been increasing worldwide, and despite notable advances in treatment, the survival rates have not improved significantly over the past few years. 3 Among the treatment options available for CCA, surgical resection has the highest cure rate. 4 Unfortunately, complete resection of the primary tumor is only possible in less than 35% of cases, and even after removal, the risk of recurrence is higher than 60%. 5,6 Chemotherapy includes the use of 5-fluorouracil (5FU), gemcitabine, and cisplatin for patients with unresectable CCA. However, the cure rate is poor. 7 Various alternate treatment strategies have been proposed for CCA, including combination therapy, immunotherapy, hormonal therapy, and targeted therapy. 8 However, the CCA treatments currently available are not adequately effective. Besides, patients experience a wide range of mild-to-severe adverse effects from these drugs. Therefore, finding potential candidate drugs with fewer adverse effects would benefit patients with limited options. Several natural compounds, including alkaloids and phenolic compounds, are being investigated for the treatment of CCA. 9,10
Phytochemicals, natural compounds derived from plants, exhibit bioactive properties, including antioxidant and anti-inflammatory effects and hormone regulation. 11 The potential mechanisms of phytochemicals include their role as substrates of biochemical reactions, cofactors of enzymatic reactions, enzyme inhibitors, and ligands that agonize or antagonize intracellular receptors. 12,13 These mechanisms highlight the therapeutic potential of phytochemicals, including their roles in preventing numerous diseases, including diabetes, cancer, cataracts, gastrointestinal disorders, and neurological disorders. 14 Consequently, extensive research on the health benefits of phytochemicals has led to the identification of compounds with high efficacy and fewer side effects for use in cancer therapy. 7,8,15
Glycyrrhiza uralensis, commonly known as licorice, one of the most popular herbs in Chinese traditional medicine, has been used to treat constipation and various diseases such as peptic ulcers, canker sores, and eczema. 16,17 Its root and rhizome have been used as an herbal medicine and as a natural sweetener. 18 It contains various phytochemicals, including glycyrrhizin, saponins, and isoquiritigenin (ISL). 19 ISL, derived from the root of Glycyrrhiza uralensis, has therapeutic properties, including anticancer, immune regulation, liver, and cardioprotective effects. 20 In recent years, the anticancer mechanisms of ISL have been studied in endometrial, breast, lung, and melanoma cancers. ISL exerts its anticancer effects by targeting the key cancer cell properties of cell proliferation, metastasis, autophagy, endoplasmic reticulum (ER) stress, and chemoresistance. ISL has been shown to induce cell cycle arrest in renal cancer via the reactive oxygen species (ROS) pathway. 21 In breast cancer, ISL was found to induce apoptosis and anti-proliferation via the Bcl-2-associated protein x (Bax), a pro-apoptotic member of the B-cell lymphoma/leukemia 2 (Bcl-2) family. 22 In ovarian cancer, ISL inhibited metastasis by reversing epithelial-to-mesenchymal transition. 23 ISL-induced apoptosis and autophagy by downregulating the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt)/mammalian target of rapamycin (PI3K/AKT/mTOR) pathway in hepatocellular carcinoma. 24 Whereas the anticancer effects of ISL have been demonstrated in various cancers, no studies have confirmed the anticancer effects of ISL in CCA.
This study investigated whether ISL reduces cell viability and proliferation in two CCA cell lines (SNU-478 and HuCCT-1). Then, we confirmed whether ISL plays a role in apoptosis and cell cycle arrest. Finally, we identified the effects of ISL on mitochondrial membrane potential (MMP), mitochondrial ROS, and autophagy to determine the pathways involved in the anticancer effects of ISL.
MATERIALS AND METHODS
Chemicals
ISL (Fig. 1A), isolated from the root of Glycyrrhiza uralensis as previously described, 25 was kindly provided by Prof. Dr. Yeon Hee Seong (Laboratory of Pharmacology, College of Veterinary Medicine, Chungbuk National University, Cheongju, Republic of Korea).

Effect of ISL on GRP78 protein levels in SNU-478 cells.
Cell cultures
SNU-478 cells were obtained from the Korea Cell Line Bank (Seoul, Republic of Korea), and HuCCT-1 cells were purchased from the Japanese Collection of Research Bioresources (Osaka, Japan). All cell lines were cultured in Roswell Park Memorial Institute (RPMI) 1640 medium (Gibco, Gaithersburg, MD, USA). The RPMI 1640 medium was supplemented with 10% fetal bovine serum (FBS, R&D Systems, Minneapolis, MN, USA), antibiotic–antimycotic solution (Gibco), and 1% HEPES buffer solution (Welgene, Gyeongsan, Republic of Korea). Subcultures were performed for subsequent experiments using 0.25% trypsin (Wellgene) at 70–80% cell density.
Cell viability assay
Cell viability was measured using the Quanti-MAX water-soluble tetrazolium salt (WST)-8 cell viability kit (Biomax, Seoul, Republic of Korea) according to the manufacturer’s instructions. SNU-478 cells (3 × 103 cells/well) and HuCCT-1 cells (4 × 103 cells/well) were seeded in 96-well plate cell culture dishes (Sarstedt, Nümbrecht, Germany). After 24 h, CCA cells were treated with various concentrations of ISL for 48 h. After incubation with WST solution for 30 min, the optical density was measured using a Neo2 hybrid multimode reader (Agilent, Santa Clara, CA, USA) at 460 nm.
Clonogenic assay
The clonogenic ability was evaluated by colony formation assay as described previously with slight modification. 26 The CCA cells were seeded at 500 cells/well in a 6-well cell culture plate (Sarstedt). After 24 h, CCA cells were treated with ISL (SNU-478: 0, 10, and 20 µM, HuCCT-1: 0, 20, and 40 µM) for 48 h. After incubation (SNU-478: 7 days, HuCCT-1: 9 days), the CCA cells were fixed with 4% paraformaldehyde (GeneAll, Seoul, Republic of Korea) and stained with 0.5% crystal violet (Sigma-Aldrich, Louis, MO, USA). Stained wells were photographed, and colony areas were measured by Image J software (National Institutes of Health, Bethesda, MD, USA).
Cell apoptosis assay
The types of cell death were evaluated using Alexa Fluor 488 Annexin V/Dead Cell Apoptosis Kit (Invitrogen, Waltham, MA, USA), according to the manufacturer’s instructions. SNU-478 cells (5 × 104 cells/well) and HuCCT-1 cells (8 × 104 cells/well) were seeded in 6-well cell culture plates (Sarstedt). After 24 h, CCA cells were treated with ISL (SNU-478: 0, 10, and 20 µM; HuCCT-1: 0, 20, and 40 µM) for 48 h. The cells were harvested in DPBS. After harvesting, CCA cells were stained with Annexin V and propidium iodide (PI). Samples were analyzed using the FACS CaliburTM instruments (BD Bioscience, San Diego, CA, USA), and data were analyzed with the FlowJo Software v. 10.8.1 (TreeStar, San Carlos, CA, USA).
Cell cycle analysis
Cell cycle distribution was evaluated using PI (Sigma-Aldrich). SNU-478 cells (5 × 104 cells/well) and HuCCT-1 cells (8 × 104 cells/well) were seeded in 6-well cell culture plates (Sarstedt). After 24 h, CCA cells were treated with ISL (SNU-478: 0, 10, and 20 µM; HuCCT-1: 0, 20, and 40 µM) for 48 h. The cells were harvested in DPBS. After harvesting, CCA cells were fixed with 70% ethanol for 2 h, and then ethanol was removed by centrifugation (879 g, 3 min). RNase A (Sigma-Aldrich, 10 µg/mL) was added and incubated at 37°C, non-CO2. After fixation, PI (50 µg/mL) was added and incubated for 1 h in the dark. Samples were analyzed using the FACS CaliburTM instruments (BD Bioscience), and data were analyzed with the FlowJo Software v. 10.8.1 (TreeStar).
MMP assay
MMP changes were analyzed using a JC-10 (Adipogen Life Sciences, San Diego, CA, USA) according to the manufacturer’s instructions with minor modifications. SNU-478 cells (4 × 103 cells/well) and HuCCT-1 cells (5 × 103 cells/well) were seeded in 96-well cell culture black plates (Corning, Somerville, MA, USA). After 24 h, CCA cells were treated with ISL (SNU-478: 0, 10, and 20 µM; HuCCT-1: 0, 20, and 40 µM) for 24 h. The cells were stained with JC-10 (5 µg/mL) for 10 min. After washing with DPBS, the stained cells were imaged with a LionheartTM FX Automated Live Cell Imager (BioTek, Winooski, VT, USA) and analyzed using Gen5 v 3.14.03 (Agilent).
Mitochondrial superoxide assays
Mitochondrial ROS were detected by the mitochondrial superoxide (MitoSOXTM) Red superoxide indicator kit (Invitrogen) according to the manufacturer’s instructions with slight modifications. SNU-478 cells (4 × 103 cells/well) and HuCCT-1 cells (5 × 103 cells/well) were seeded in 96-well cell culture black plates (Corning). After 24 h of incubation, CCA cells were treated with ISL (SNU-478: 0, 10, and 20 µM; HuCCT-1: 0, 20, and 40 µM) for 24 h. CCA cells were stained with MitoSOXTM (Invitrogen, 5 µM) and Hoechst 33342 (Sigma-Aldrich, 10 µg/mL) for 10 min. After washing with DPBS, the stained cells were imaged with a LionheartTM FX Automated Live Cell Imager (BioTek) and analyzed using Gen5 v 3.14.03 (Agilent).
Lysosomal membrane permeability assay
Acridine orange (Sigma-Aldrich) staining was performed to detect lysosomal membrane permeability according to the manufacturer’s instructions with slight modifications. SNU-478 cells (4 × 103 cells/well) and HuCCT-1 cells (5 × 103 cells/well) were seeded in 96-well cell culture black plates (Corning). After 24 h of incubation, CCA cells were treated with ISL (SNU-478: 0, 10, and 20 µM; HuCCT-1: 0, 20, and 40 µM) for 24 h. CCA cells were stained with acridine orange hydrochloride solution (2 µg/mL) for 20 min. After washing with DPBS, the stained cells were imaged with a LionheartTM FX Automated Live Cell Imager (BioTek) and analyzed using Gen5 v 3.14.03 (Agilent).
Western blot analysis
The SNU-478 cells were treated with ISL (0, 10, and 20 µM) for 24 h. Total protein was extracted with PRO-PREP protein extraction solution (iNtRON Biotechnology, Seongnam, Republic of Korea). The concentration of proteins was measured using a bicinchoninic acid reagent (Sigma-Aldrich). Approximately 50 µg of protein lysates were loaded in 12% sodium dodecyl sulfate-polyacrylamide gel, and electrophoresis was conducted. Loaded proteins were transferred to a polyvinylidene fluoride membrane (Bio-Rad, Berkeley, CA, USA), and the membrane was blocked by 5% skim milk for 2 h. The membrane was incubated with primary antibodies against glucose-regulated protein 78 (GRP78, 1:1000, sc-13539, Santa Cruz, Dallas, Texas, USA), Bax (1:1000, 4223S, Cell Signaling Technology, Danvers, MA, USA), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH, 1:1000, ABS16, Millipore, Burlington, MA, USA) overnight at 4°C. Horseradish peroxidase-conjugated goat anti-rabbit IgG (H + L) (Bio-Rad, 1:3000) or goat anti-rat IgG (H + L) (Invitrogen, 1:5000) were used as secondary antibodies. Target proteins were detected using LuminoGraph 2 (ATTO Corporation) with SupersignalTM West Femto or Pico (1:4 mix, Rockford, IL, USA). All protein expression levels were normalized to the GAPDH expression levels and normalized by Image J software.
Statistical analysis
All experiments were performed at least three independent times, and all data were presented as means ± standard deviations (SD). The statistical significance of data was determined by one-way analysis of variance followed by a post hoc Dunnett’s test using the GraphPad Prism 5.01 software (GraphPad Software Inc., San Diego, CA, USA). The P < .05 was considered statistically significant.
RESULTS
ISL downregulated the GRP78 protein expression in SNU-478 cells
To determine whether ISL downregulates GRP78 expression in CCA cells, we used Western blotting to measure GRP78 protein levels in ISL-treated SNU-478 cells (Fig. 1B). The ISL (20 µM) treatment significantly decreased the levels of GRP78 protein expression in SNU-478 cells compared with those of the control group (Fig. 1C).
ISL decreased the cell viability and proliferation of CCA cells
To evaluate the effect of ISL on the viability of CCA cells, a WST assay was performed on the SNU-478 and HuCCT-1 cells. ISL treatment for 48 h decreased cell viability, and the half-maximal inhibitory concentration (IC50) values of ISL for the SNU-478 cells and HuCCT-1 cells were 30.16 and 49.22 µM, respectively (Fig. 2A, B). A clonogenic assay was performed to confirm the effect of ISL on proliferation by evaluating the ability of a single cell to grow into a colony (Fig. 2C, E). ISL treatment (at 10 and 20 µM for SNU-478 cells, and 20 and 40 µM for HuCCT-1 cells) significantly reduced the colony area when compared with the control group (Fig. D, F). These results indicate that ISL decreased the cell viability and proliferation of CCA cells.
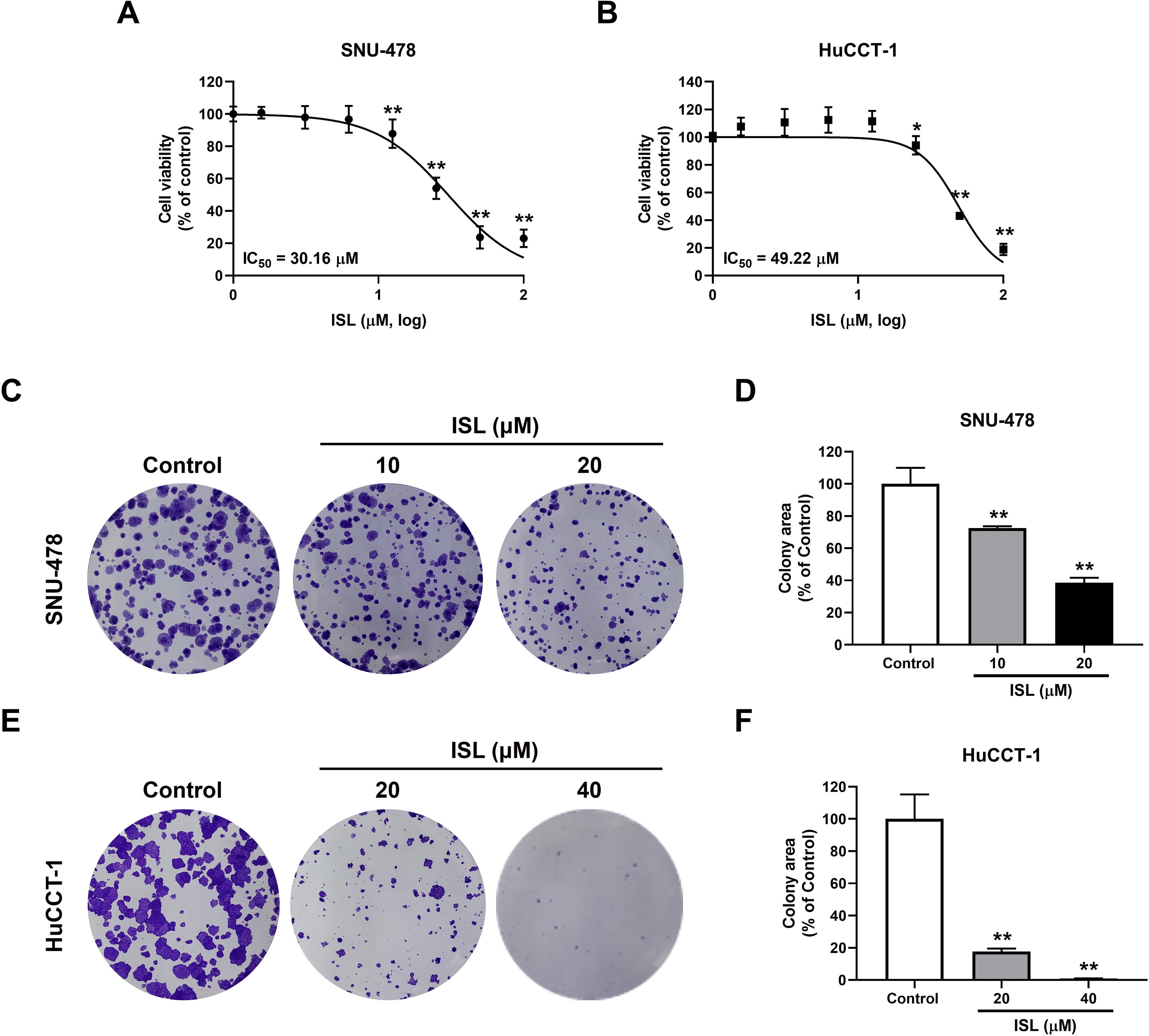
Effect of ISL on the cell viability and colony-forming capacity in CCA cells. The cell viability and IC50 value of
ISL increased the apoptotic death of CCA cells
To determine whether ISL-mediated apoptotic death occurs in CCA cells, an Annexin V/PI staining assay was performed (Fig. 3A, C). ISL treatment (at 20 µM for SNU-478 cells and 20 and 40 µM for HuCCT-1 cells) significantly increased the Annexin V-positive population compared to the control group (Fig. 3B, D). In addition, we evaluated the changes in the levels of an apoptosis-related protein, Bax (Fig. 3E). ISL (20 µM) significantly increased the Bax expression levels compared with the control (Fig. 3F). These results indicate that ISL increased the apoptotic death of CCA cells.

Effect of ISL on the apoptotic cell death of CCA cells. Representative flow cytometry plot of
ISL inhibited cell cycle progression in CCA cells
For cell cycle analysis, we performed PI staining to identify the proportion of cells in each of the three interphase stages of the cell cycle (Fig. 4A, C). ISL treatment (at 20 µM for SNU-478 cells, and 40 µM for HuCCT-1 cells) significantly increased the G2/M phase distribution compared with the control (Fig. 4B, D). ISL treatment at 20 and 40 µM significantly increased the S-phase distribution specifically in HuCCT-1 cells compared to the control. These results suggest that ISL treatment induced cell cycle arrest in CCA cells.

Effect of ISL on cell cycle arrest in CCA cells. Representative images of PI staining in
ISL decreased the MMP level in CCA cells
MMP is an overall indicator of mitochondrial function. Cancer cells have abnormally high MMP, associated with various cellular functions, including metastasis. 27 To evaluate the effects of ISL on MMP, we performed JC-10 staining (Fig. 5A, C). In normal cells, JC-10 concentrates in the mitochondrial matrix, forming red fluorescent aggregates. In apoptotic and necrotic cells, JC-10 diffuses out of the mitochondria and becomes a green fluorescent monomer. A decreasing JC-10 red/green fluorescence ratio indicates a decrease in MMP. The ISL treatment (at 10 and 20 µM for SNU-478 cells and 40 µM for HuCCT-1 cells) significantly decreased the fluorescence intensity of the JC-10 dimers compared with the control (Fig. 5B, D). These results indicate that the ISL treatment induced mitochondrial dysfunction of the CCA cells by decreasing the MMP levels.

Effect of ISL on mitochondrial membrane potential (MMP) in CCA cells. MMP after treatment of ISL was assessed by JC-10 staining. Representative images of JC-10 dye in
ISL upregulated mitochondrial ROS in CCA cells
Since mitochondrial dysfunction is associated with the upregulation of oxidative stress, 28,29 we evaluated the effect of ISL on mitochondrial ROS levels using MitoSOXTM staining (Fig. 6A, C). ISL treatment (at 20 µM for SNU-478 cells and 40 µM for HuCCT-1 cells) significantly increased the MitoSOX fluorescence intensity compared with the controls (Fig. 6B, D). These results indicate that the ISL treatment might induce mitochondrial oxidative stress, leading to mitochondrial dysfunction.

Effect of ISL on mitochondrial reactive oxygen species (ROS) in CCA cells. The content of mitochondrial ROS was determined by MitoSOX staining. Representative images of MitoSOX in
ISL upregulated autophagy in CCA cells
Numerous stresses, including hypoxia, intracellular ROS levels, bacteria, and chemical drugs, regulate autophagy. 30,31 We determined whether ISL upregulated the induction of autophagy using acridine orange staining (Fig. 7A, C). ISL treatment (at 20 µM for SNU-478 cells and 20 and 40 µM for HuCCT-1 cells) significantly increased acridine orange-red fluorescence intensity compared with the control (Fig. 7B, D). These results indicate that the ISL treatment upregulated the autophagy of CCA cells.

Effect of ISL on autophagy in CCA cells. The content of autophagy was determined by acridine orange staining. Representative images of acridine orange dye in
DISCUSSION
Accumulating research indicates that GRP78 is involved in several aspects of tumor development, including cell proliferation, autophagy, chemosensitization, and apoptosis. 32 –34 GRP78 is a chaperone protein located in the ER membrane of eukaryotic cells. It plays an important role in protein folding and maturation and regulates ER stress. ER stress is commonly activated in cancer cells due to the accumulation of misfolded proteins caused by genetic mutations and genomic rearrangements. 35 The unfolded protein response overexpresses chaperone proteins, including GRP78, which play a role in the adaptation to ER stress. 36 Thus, within the tumor microenvironment, GRP78 acts as a stress sensor and is frequently upregulated. Inhibition of GRP78 increases ER stress, autophagy, and cell death. 37,38 ISL is known to upregulate ER stress through GRP78 inhibition in various cancers, including ovarian, breast, and gastric cancers. 39,40 In the current study, ISL decreased GRP78 protein levels in CCA cells. ISL-mediated inhibition of GRP78 might exert anticancer effects by modulating ER stress in CCA.
ER stress is known to downregulate cancer cell proliferation through cell cycle arrest. 41 Previous studies have demonstrated that ISL treatment arrests the G2/M phase in cervical and ovarian cancer cells, 42,43 and also increases markers of DNA replication damage, including γ-H2A histone family member X foci formation. 43 In the present study, ISL-induced cell cycle arrest in both S and G2/M phases in CCA cells might be associated with GRP78 inhibition. Intriguingly, in the current study, ISL-induced G2/M phase arrest in SNU-478 cells, whereas both S and G2/M phase arrests were observed in HuCCT-1 cells. This discrepancy suggests that the response to ISL is cell line-dependent, possibly due to differences in cell cycle regulation or ISL sensitivity. Further research is needed to clarify the underlying mechanisms of these differences, particularly focusing on cell cycle regulators and DNA damage response pathways. Additionally, ISL decreased the clonogenic ability of CCA cells, indicating that ISL exerts its anticancer effect through downregulation of CCA proliferation. A previous study revealed that cell cycle arrest by ER stress is associated with the cyclin-dependent kinase (CDK) family. 41 In liver cancer cells, ISL has been shown to reduce the proliferation and colony formation capacity by inhibiting the CDK family. 44 Further studies are needed to investigate whether ISL regulates the cell cycle through CDK modulation in CCA.
Accumulating evidence implicates ER stress-induced mitochondrial dysfunction in cancer cells. 45 The ER plays an important role in regulating protein folding, lipid synthesis, and calcium homeostasis, and ER dysfunction leads to mitochondrial dysfunction. 46,47 Mitochondrial dysfunction induces decreased adenosine triphosphate (ATP) production and impaired cellular function through decreases in MMP. 48 Treatment with ISL was observed to increase apoptosis through MMP reduction in lung, ovarian, and pancreatic cancers. 49,50 In the current study, ISL decreased MMP and mitochondrial ROS in CCA cells, suggesting that ISL could induce mitochondrial dysfunction through the ER stress pathway.
Mitochondria are involved in a variety of cellular functions, including ion homeostasis, ROS regulation, and programmed cell death. 51 Cellular stress signals, including persistent ROS, are involved in mitochondrial dysfunction through the regulation of the proteins that affect MMP permeability, including Bcl-2 family proteins. 52,53 Prior evidence indicates that ISL induces mitochondrial dysfunction through ER stress in various cancers, including cervical, melanoma, and prostate cancer. 54 –56 Consistent with previous findings, the current study demonstrated that ISL induces mitochondrial ROS and mitochondrial dysfunction in CCA cells, suggesting that its anticancer effects are mediated through mitochondrial targeting.
ER stress causes mitochondrial dysfunction and increases mitochondrial ROS production. 21,55,57 ROS play an important role in regulating cellular processes, including cell proliferation, death, and progression. 29 Redox homeostasis acts as a signal in cancer cells, allowing proliferation, but an imbalance in ROS induces cell death through mitochondrial damage. 57 ISL acts as an antioxidant enzyme and upregulates ROS, which promotes apoptosis, as observed in pancreatic cancer cells. 58 In this study, ISL significantly decreased the MMP and upregulated mitochondrial ROS in CCA cells. These findings suggest a potential relationship with the inhibition of the GRP78 protein.
Mitochondrial dysfunction due to persistent ROS and reduction in MMP increases apoptosis. 59 ISL induces apoptosis through the regulation of the Bcl-2 family proteins in breast cancer and osteosarcoma. 22,60 Apoptosis is a regulated and highly conserved process of cell death that involves self-destruction. It is an essential process in multicellular organisms, preventing carcinogenesis by removing potentially harmful cells with DNA damage. 61 Various stress signals, including heat shocks, ROS, and hypoxia, trigger cell death. 62 Exposure to chemicals such as pesticides and anticancer drugs triggers cell death, a process that is generally mediated by ROS. In an earlier study in cancer cells, ISL-induced apoptosis by regulating apoptosis-associated proteins, including Bcl-2 and Bax. 63 Similarly, in this study, ISL significantly upregulated apoptosis in CCA cells, possibly due to ROS upregulation.
Autophagy is a homeostatic response in eukaryotic cells that regulates the removal of proteins and damaged organelles. 47,64 Autophagy is upregulated via various stresses, including the accumulation of unfolded proteins, ROS, and mitochondrial dysfunction. 65 –67 Autophagy upregulated by persistent stress causes cell death by increasing the permeability of the lysosomal membranes. 68,69 In this study, ISL effectively upregulated apoptosis, mitochondrial ROS, and autophagy in CCA cells, suggesting its potential link between autophagy and the regulation of CCA cell death. However, further mechanistic studies are needed to confirm ISL-induced autophagy in CCA cells.
Although the current study provides valuable insight into the anticancer effects of ISL in CCA, further research is needed to elucidate the underlying mechanism more precisely. Future studies exploring various pathways, including cell cycle regulation, ER stress, and autophagy, will help uncover the molecular mechanisms involved. Additionally, in vivo experiments, including pharmacokinetic and pharmacodynamic evaluations, are essential for assessing the clinical therapeutic potential of ISL. Investigating the interactions between ISL and existing therapeutic agents could also provide valuable information for developing a new therapeutic intervention to enhance treatment efficacy. Taken together, these studies will enhance our understanding of ISL’s role in CCA and may contribute to developing novel treatment strategies.
In conclusion, this study demonstrates that ISL treatment downregulates GRP78 in CCA cells, which in turn induces cell cycle arrest, a decrease in MMP, excessive mitochondrial ROS generation, and autophagy. These findings suggest that ISL exerts its anticancer effects through mitochondrial dysfunction-induced apoptosis, providing scientific evidence that ISL could be a potential therapeutic agent for treating CCA.
Footnotes
AUTHORS’ CONTRIBUTIONS
Conceptualization: H.K.L. and K.C.C. Methodology: S.H.B., H.N., and H.K.L. Validation: S.H.B. Formal analysis: S.H.B. Investigation: S.H.B. and H.N. Data curation: H.K.L. and K.C.C. Writing—original draft: S.H.B. Writing—review and editing: H.K.L. and K.C.C. Visualization: S.H.B. Supervision: K.C.C. Project administration: H.K.L. and K.C.C. Funding acquisition: K.C.C.
AUTHOR DISCLOSURE STATEMENT
The authors do not have any conflicts of interest to declare.
DATA AVAILABILITY STATEMENT
All data generated or analyzed in this study are included in this published article.
FUNDING INFORMATION
This study was supported by the “
SUPPLEMENTARY MATERIAL
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.