
Abstract
Locusta migratoria (LM) is an edible insect recognized as a valuable source of high-quality proteins, essential amino acids, vitamins, and minerals. Metabolic dysfunction-associated steatotic liver disease (MASLD) is often associated with metabolic syndrome (MetS) and is characterized by excessive fat accumulation in the liver along with low-grade chronic inflammation. In addition, chronic metabolic dysfunction and hepatic lipid toxicity can induce acute liver injury (ALI). This study investigated the protective effects of LM hydrolysate against MASLD and the related liver pathophysiology. Mice were fed a high-fat, high-fructose, and high-cholesterol diet to induce MASLD, whereas ALI was induced using lipopolysaccharide (LPS) and d-galactosamine (Gal). LM hydrolysate reduced liver weight by decreasing hepatic ectopic fat accumulation and downregulating lipogenic gene expression in the liver. In addition, LM hydrolysate improved dyslipidemia by lowering the serum triglyceride and low-density lipoprotein cholesterol levels. LM hydrolysate also demonstrated hepatoprotective properties by reducing the protein levels of tumor necrosis factor-alpha and interleukin-6 while enhancing antioxidant capacity, thereby mitigating liver damage induced by LPS/Gal under MASLD-promoting conditions. Thus, LM hydrolysate significantly attenuated the pathological changes in the hepatic tissue. These findings suggest that LM hydrolysate has potential as a functional food or dietary supplement for managing MetS-related disorders and protecting against liver pathology.
INTRODUCTION
Edible insects have been a fundamental part of human diets and served as a promising food resource in various cultures for centuries, particularly in Africa, Asia, and Latin America. 1 The global practice of entomophagy underscores the importance of insects as a sustainable nutritional resource, and insects are often incorporated into traditional dishes and culinary practices. 2 Recently, interest in edible insects has grown in Western countries owing to their environmentally friendly characteristics and potential role in promoting sustainability. 3 Increased interest in entomophagy has been driven by increasing awareness of the environmental impact of conventional livestock farming, which contributes significantly to greenhouse gas emissions, deforestation, and water consumption. 4
Edible insects offer substantial environmental advantages over traditional livestock. For example, crickets require only one-sixth of the feed cattle need to produce an equivalent amount of dietary protein. 5 Relatively lower carbon production positions edible insects as a sustainable and eco-friendly alternative to conventional animal protein sources, aligning with global initiatives to reduce carbon footprints and promote sustainable food systems. 6 Furthermore, edible insect farming can be integrated into urban environments and small-scale operations, offering flexibility and accessibility beyond the reach of traditional livestock farming. 7 Breeding adaptability further enhances the suitability of insect farming to meet global challenges in food security and environmental conservation.
Edible insects are rich in essential amino acids, fatty acids, water-soluble vitamins, and minerals, making them a highly nutritious food source. 8 The nutrient density of edible insects makes insects an excellent option for addressing nutritional deficiencies and providing supplemental nutrients. In South Korea, 10 types of edible insects have been designated as promising future food sources and approved by the Ministry of Food and Drug Safety. 9 Among these, Locusta migratoria (LM) stands out as an exceptional protein source, containing up to 76% protein. 10 Being an herbivorous insect, LM is also rich in retinol, vitamins, carotenoids, and antioxidants, highlighting its potential as a functional food that can address metabolic diseases while supporting nutritional and environmental sustainability. 11 Furthermore, by-products of insect farming, such as frass, serve as high-quality fertilizers that improve soil quality and promote sustainable agriculture. 12 Notably, LM frass has been shown to significantly attenuate de novo adipogenesis in both in vivo and in vitro models. 13 These combined nutritional and environmental benefits position edible insects as a vital component of future food security strategies, holistically addressing health and sustainability challenges.
Metabolic syndrome (MetS) is a prevalent and complex condition characterized by a cluster of metabolic abnormalities, including central obesity, insulin resistance, dyslipidemia, and hypertension. 14 A meta-analysis involving 28 million individuals globally revealed that the prevalence of MetS can reach up to 31.4%, with higher rates being observed in middle- and high-income countries than in low-income regions. 15 In the United States, ∼34.7% of adults are affected, 16 whereas the prevalence rates in Europe, 17 Korea, 18 and China 19 are 13–35%, 31.3%, and 31.1%, respectively. The significant prevalence of MetS imposes considerable health care costs, including increased hospitalizations, greater utilization of medical services, and management of complications. 20 MetS is also associated with an increased risk of metabolic-associated steatohepatitis (MASH), a spectrum of histological abnormalities that can progress to fibrosis and cirrhosis, ultimately leading to advanced liver disease. 21 Excess ectopic fat accumulation in the liver plays a central role in MASH development, triggering inflammatory responses that cause hepatocellular damage and functional decline. 22 Chronic metabolic complications contribute to sustained liver damage and elevated risk of acute liver injury (ALI). 23
ALI, characterized by extensive hepatocyte necrosis and a rapid decline in liver function, is a life-threatening condition often caused by drugs and viruses. 24 Hepatitis B is the leading cause of ALI in Asian countries such as China and Korea, while the widespread use of acetaminophen contributes to its higher incidence rates in Western countries. 25 Prolonged progression of metabolic dysfunction-associated steatotic liver disease (MASLD) can lead to MASH, and if left unresolved, ALI may develop. 26 Lipid peroxidation caused by excessive free radicals results in severe cellular membrane damage, further exacerbating hepatic dysfunction. Early stage ALI can be managed with N-acetylcysteine, which replenishes liver glutathione (GSH) levels, neutralizes harmful free radicals, and reduces oxidative stress and inflammation. 27 Supplementation with vitamins C and E, along with plant-derived antioxidants, has shown promise in alleviating inflammation and oxidative stress, providing critical support in ALI management. 28 Effective management of ALI requires liver regeneration, which depends on adequate intake of high-quality proteins to facilitate hepatocyte repair and maintain immune function. 29
Both LM and LM hydrolysate, rich in high-quality amino acids, possess bioactive components with antioxidant, antibacterial, and anti-inflammatory properties. 30 LM consumption enhances intestinal protein digestion rates in rats, reduces hepatic and plasma lipoprotein expression and enzyme activity, and restores normal hepatic homeostasis. 31 Previous in vitro experiments demonstrated that LM enzymatic hydrolysate alleviated hepatic cytotoxicity and inflammatory responses induced by lipopolysaccharide/d-galactosamine (LPS/Gal), a well-established experimental model of ALI, in human hepatoma G2 (Hep G2) cells. 32 These findings underscore the potential of LM as a nutraceutical agent for managing MASLD and ALI.
In this study, we designed a comprehensive experimental scheme to evaluate the hepatoprotective effects of LM hydrolysate in animal models. MASLD was induced in C57BL/6J mice through an 8-week high-fat, high-fructose, and high-cholesterol (H) diet, with LM hydrolysate administered daily via oral gavage. Subsequently, ALI was induced by intraperitoneal injection of LPS and Gal under MASLD conditions. Biological indices were assessed to investigate the effects of LM hydrolysate, including body and organ weight changes, serum biochemical parameters [aspartate aminotransferase (AST), alanine transaminase (ALT), triglyceride (TG), low-density lipoprotein cholesterol (LDL-C), and high-density lipoprotein cholesterol (HDL-C)], hepatic histological analysis (inflammatory cell infiltration and vacuolization), and markers of inflammation, oxidative stress, and lipid metabolism. These indices provide a robust framework for determining the therapeutic potential of LM hydrolysate in mitigating liver pathology. The aim of this study was to evaluate whether LM hydrolysate can prevent the progression of MASLD induced by H diet and alleviate ALI induced by LPS/Gal, thereby assessing the potential of LM hydrolysate in managing metabolic or acute liver injuries.
MATERIALS AND METHODS
Preparation of LM hydrolysate
LM powder was obtained from three domestic insect farms in Korea in June 2022. A 10% (w/v) suspension was prepared by dissolving LM powder in distilled water. The pH of the suspension was adjusted to 7.0 before adding 1% (v/v) Neutrase® (Novozymes, Bagsvaerd, Denmark) for enzymatic hydrolysis. The mixture was then incubated at 50°C for 8 h. Enzymatic activity was terminated by heating the mixture to 90°C for 30 min. The hydrolysate was filtered through a 100-mesh sieve, freeze-dried, and stored at −70°C for use in animal experiments.
Animal experiments
All animal experiments were conducted in accordance with the guidelines approved by the Institutional Animal Care and Use Committee of Dankook University (IACUC No. DKU-23-018). Five-week-old male C57BL/6J mice (Raon Bio, Yongin, Korea) were housed under controlled conditions (temperature = 22 ± 2°C, relative humidity = 50–55%) with a 12-h light/dark cycle. The mice were acclimatized for 1 week with ad libitum access to their designated diets and filtered water.
Following acclimatization, the mice were randomly assigned to one of three groups (n = 22 per group): (1) low-fat (L) diet (D09100304), macronutrient-matched to the H diet, containing only 10% kcal from fat and lacking fructose and cholesterol supplementation; (2) H diet (D09100310), a MASH model diet containing 40% kcal from fat (primarily palm oil), 20% kcal from fructose, and 2% cholesterol; and (3) H diet supplemented with LM hydrolysate administered via oral gavage for 8 weeks. Mice in the L and H diet groups were administered Dulbecco’s phosphate-buffered saline (DPBS; Welgene, Gyeongsan, Korea) as a vehicle control. Dietary intake and body weight were measured and recorded at a consistent time on a weekly basis. The food efficiency ratio (FER) was calculated using the following formula: (total weight gain/total food intake) × 100. 33,34
After 8 weeks of dietary intervention, half of the mice in each group were randomly selected to receive intraperitoneal injection of either DPBS (control) or a combination of LPS (50 µg/kg; Sigma-Aldrich, St. Louis, MO) and Gal (400 mg/kg; Sigma-Aldrich) to induce ALI. The mice were sacrificed 5 h post-injection via thoracotomy under isoflurane anesthesia. Blood was collected through cardiac puncture, and the serum was separated by centrifugation at 3000 g at 4°C for 15 min. Liver and white adipose tissues (WATs) were harvested, weighed, and stored at −70°C for further analyses.
Serum biochemistry and lipid panel
Serum TG and total cholesterol (TC) levels were measured using commercial kits according to our previous research (TG and TC Assay Kits; Embiel, Gunpo, Korea). 35,36 HDL-C and LDL-C levels were analyzed using the EZ-HDL and LDL/VLDL Assay Kits (DoGenBio, Seoul, Korea). Cardiac risk factor (CRF) calculations were performed according to previous studies. 37,38 Serum cytokine levels were quantified using the Mouse High Sensitivity T Cell Multiplex Kit (Merck, Darmstadt, Germany). AST and ALT levels were assessed using AST and ALT assay kits (Embiel). 39,40
Histological examination of the liver
Liver tissues were fixed in 10% neutral-buffered formalin (Sigma-Aldrich), embedded in paraffin, and sectioned at 3–4 µm thickness using a microtome. Hematoxylin and eosin staining was performed, and histological images were captured at 20× magnification using an optical microscope (NEO Science, Daejeon, Korea). 41,42 Pathological changes were evaluated using Suzuki’s histological criteria on randomly selected microscopic images. 43
Hepatic lipid content
Hepatic lipids were extracted using the Bligh and Dyer method and validated using in-house protocols. 44,45
Enzyme-linked immunosorbent assay
Approximately 20 mg of the liver tissue was rinsed with cold DPBS to remove any residual blood. The tissue was homogenized in RIPA buffer (ATTO, Tokyo, Japan) containing cOmplete™ Protease Inhibitor Cocktail (Roche, Basel, Switzerland) and PhosSTOP™ tablets (Roche). The homogenate was centrifuged, and the supernatant was collected for analysis. Tumor necrosis factor-alpha (TNF-α) and interleukin (IL)-6 concentrations were measured using enzyme-linked immunosorbent assay kits (BioLegend, San Diego, CA) following the manufacturer’s instructions.
Liver oxidative stress markers
Liver oxidative stress markers, including malondialdehyde (MDA), superoxide dismutase (SOD), GSH, and glutathione disulfide (GSSG), were quantified using commercial kits (MDA Assay Kit, Dojindo, Kumamoto, Japan; EZ-SOD and EZ-Glutathione Assay Kits, DoGenBio).
Quantitative polymerase chain reaction
Total RNA was extracted from the liver tissues using NucleoZol® reagent (Macherey-Nagel, Düren, Germany). Complementary DNA was synthesized from 2 µg of RNA using the Moloney murine leukemia virus Reverse Transcriptase Kit (Bioneer, Daejeon, Korea). Quantitative reverse transcription polymerase chain reaction (qRT-PCR) was performed using AccuPower® 2× GreenStar™ qPCR Master Mix (Bioneer) on CFX Connect Real-Time PCR Detection System (Bio-Rad, Hercules, CA). Primer sequences are listed in Table 1.
List of Primers Used in Quantitative Reverse Transcription Polymerase Chain Reaction
Acc, acetyl-coenzyme A carboxylase beta; Acox1, acyl-coenzyme A oxidase 1, palmitoyl; Cebpa, CCAAT/enhancer binding protein alpha; Cd36, CD36 molecule; Fasn, fatty acid synthase; Gapdh, glyceraldehyde-3-phosphate dehydrogenase; Hmox1, heme oxygenase 1; Il-1β, interleukin 1 beta; Il-6, interleukin 6; Srebp1, sterol regulatory element binding transcription factor 1; Tnf-α, tumor necrosis factor alpha.
Western blot analysis
Liver proteins were extracted using RIPA buffer supplemented with protease and phosphatase inhibitors. Protein concentrations were determined using the Pierce™ BCA Protein Assay Kit (Thermo Fisher Scientific, Waltham, MA). Equal amounts of protein (20 µg per lane) were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis, transferred to polyvinylidene fluoride membranes (Millipore, Burlington, MA), and blocked with EveryBlot Blocking Buffer (Bio-Rad). The membranes were incubated with primary antibodies overnight at 4°C, followed by incubation with secondary antibodies for 1 h. Proteins were visualized using the EZ-Western Lumi Pico Alpha Kit (DoGenBio) on iBright 750 Imaging System (Invitrogen, Waltham, MA). Band intensities were analyzed using the ImageJ software (version 1.8.0; NIH, Bethesda, MD). Detailed antibody information is presented in Table 2.
Antibodies for Western Blot Analysis
BiP, binding immunoglobulin protein; CST, Cell Signaling Technology (Danvers, MA); HO-1, heme oxygenase-1; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; p-NF-κB, phospho-NF-κB; p-SAPK/JNK, phospho-SAPK/JNK; SC, Santa Cruz (Dallas, TX); SAPK/JNK, stress-activated protein kinase/c-Jun NH(2)-terminal kinase.
Statistical analysis
Data are expressed as means ± standard deviations. One-way analysis of variance (ANOVA) was used to analyze the effects of dietary intervention, and two-way ANOVA was employed to evaluate the interaction between diet and LPS/Gal stimulation, followed by Tukey’s post hoc test. Statistical significance was set at P < .05. The analyses were performed using XLSTAT 2019 (Addinsoft Inc., Paris, France) or SPSS 26.0 (IBM Corp., Armonk, NY). Graphs were generated using GraphPad Prism (version 10.1.1; GraphPad Software, San Diego, CA). The results of two-way ANOVA for dietary intervention, LPS/Gal injection, and their interactions are summarized in Supplementary Table S1.
RESULTS
Effects of H diet, LPS/Gal challenge, and LM hydrolysate on body and organ weights
During the 8-week dietary intervention, mice fed H diet exhibited significantly higher body weights than did those fed L diet, with differences becoming apparent after 4 weeks of intervention (Fig. 1A), regardless of LM treatment. Despite the differences in body weight, the estimated daily food intake remained consistent across all groups (Fig. 1B). However, the FER was significantly higher in all H diet groups, independent of LM treatment (Fig. 1C).
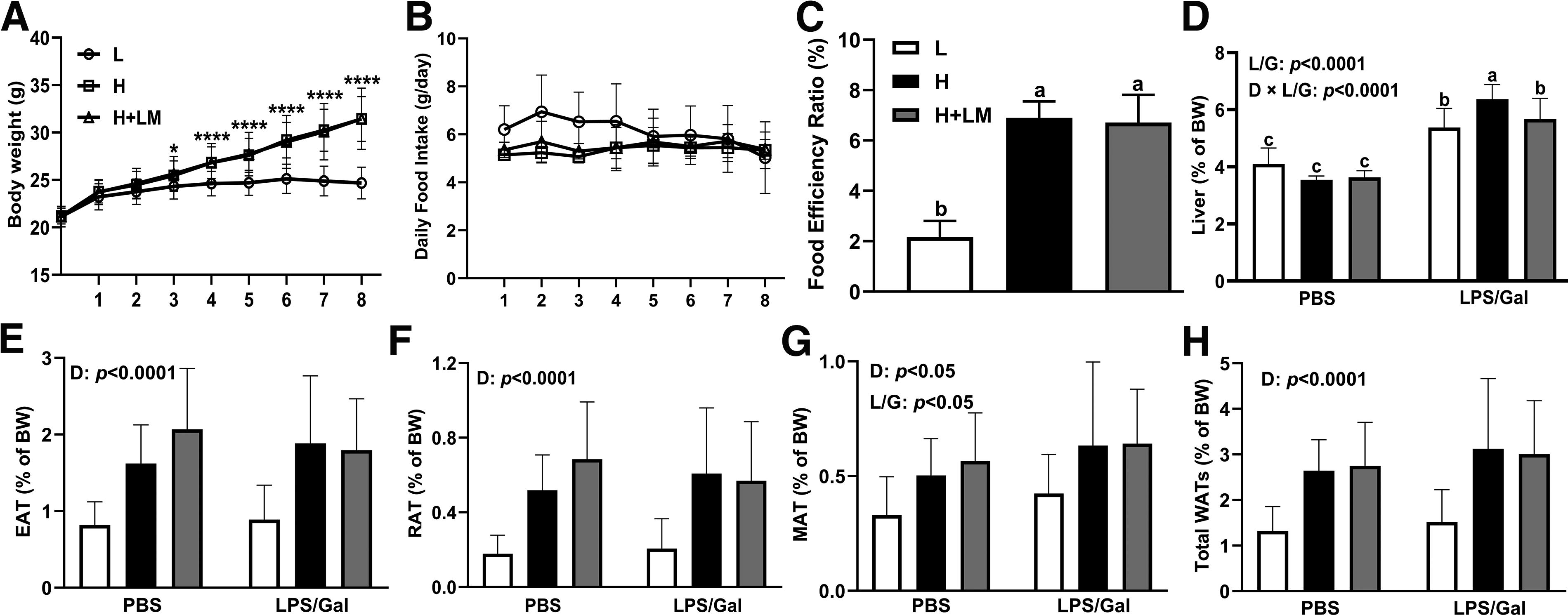
Effects of an H diet and LM hydrolysate on the BW change, daily food intake, FER, liver, and adipose tissue weights in C57BL/6J mice. Five-week-old male C57BL/6J mice were fed either an L or an H diet for 8 weeks. Concurrently, mice on the H diet received LM hydrolysate by gavage and were then treated with DPBS or LPS (50 µg/kg)/Gal (400 mg/kg).
Given the increase in body weight, the liver and WAT weights were examined. LPS/Gal intervention increased liver weight by 1.55-fold, and liver weight was significantly elevated in mice fed the H diet under LPS/Gal conditions compared with that in mice fed the L diet or receiving the LM intervention (dietary × LPS/Gal effect; P < .0001). Although our supplementary data showed that the H diet increased the absolute liver weight in mice (Supplementary Fig. S1; dietary effect; P < .0001), we attributed this change to the increased body weight caused by the H diet, with the primary cause of liver weight increase being the LPS/Gal intervention. Notably, LM hydrolysate reduced liver weight to levels comparable with those in L diet-fed mice (Fig. 1D), suggesting that LM hydrolysate may alleviate MASLD and mitigate secondary damage caused by LPS/Gal (LPS/Gal effect; P < .0001).
In terms of WATs, H dietary intervention significantly increased epididymal adipose tissue (EAT, dietary effect; P < .0001), retroperitoneal adipose tissue (RAT, dietary effect; P < .0001), mesenteric adipose tissue (MAT, dietary effect; P < .05), and total WATs weight (dietary effect; P < .0001) by 2.06-, 2.92-, 1.50-, and 2.03-fold, respectively (Fig. 1E–H). LPS/Gal increased MAT weight (LPS/Gal effect; P < .05) by approximately 1.29-fold (Fig. 1G). However, LM hydrolysate did not significantly alter WATs weight in either the dietary or LPS/Gal intervention group.
Effects of H diet, LPS/Gal challenge, and LM hydrolysate on serum hepatic functional parameters and lipid profile
Abnormal serum AST and ALT levels are indicative of liver injury. In this study, LPS/Gal significantly increased ALT (LPS/Gal effect; P < .0001) and AST (LPS/Gal effect; P < .0001) levels by 110-fold and 1.46-fold, respectively (Fig. 2A, B). Interestingly, although neither H diet nor LM hydrolysate intervention affected ALT levels (Fig. 2A), AST levels (dietary effect; P < .05) were reduced by 1.14-fold in H diet-fed mice compared with those in L diet-fed mice (Fig. 2B). Short-term intervention with the H diet did not cause severe inflammation or fibrosis that could affect liver biomarkers, whereas LPS/Gal did significantly induce ALI.
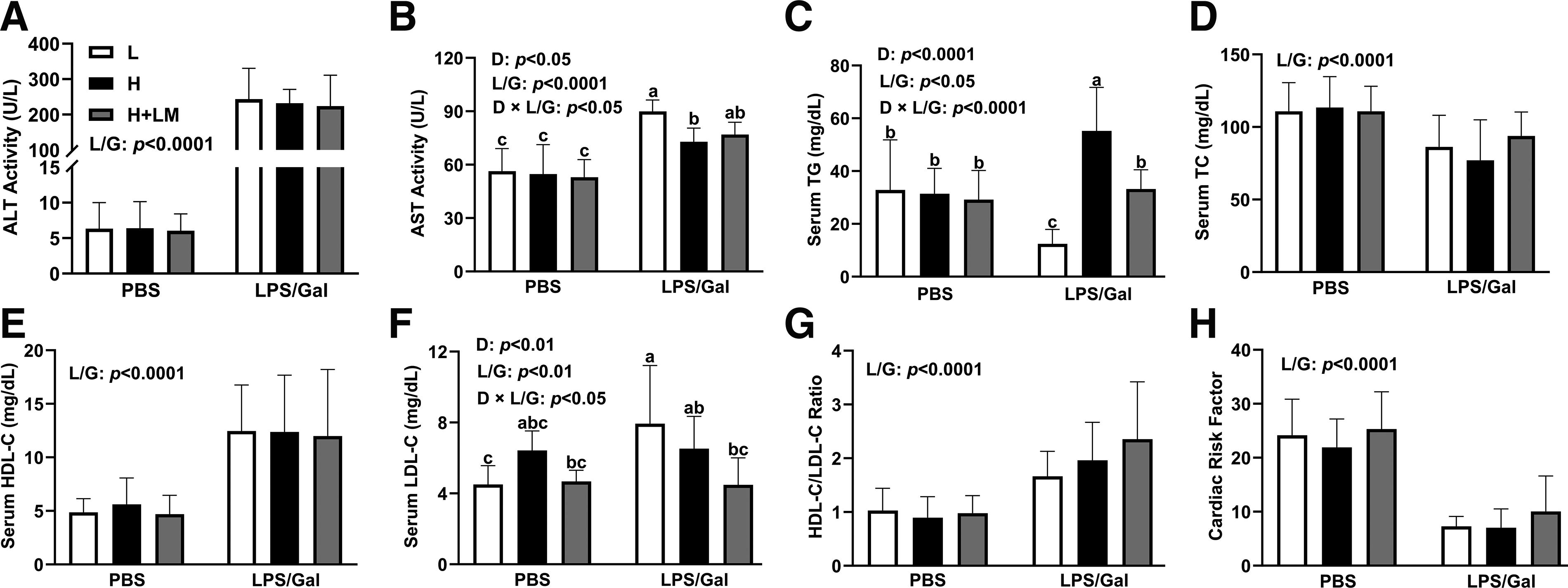
Effects of an H diet, LM hydrolysate, and LPS/Gal challenge on serum hepatic function parameters and lipid profiles in C57BL/6J mice. Five-week-old male C57BL/6J mice were fed either an L diet or an H diet for 8 weeks. Concurrently, mice on the H diet received LM hydrolysate by gavage and were then treated with DPBS or LPS (50 μg/kg)/Gal (400 mg/kg).
Dyslipidemia is closely associated with the development of MASLD. Dietary intervention significantly altered the serum TG (dietary effect; P < .05) and LDL-C (dietary effect; P < .01) levels. Furthermore, coadministration of LPS/Gal also increased the serum TG (LPS/Gal effect; P < .0001) and LDL-C levels (LPS/Gal effect; P < .01) by 1.19-fold and 1.26-fold, respectively. Importantly, experimental mice fed the H diet combined with LPS/Gal injection exhibited a 2.35-fold increase in TG levels (dietary × LPS/Gal effect; P < .0001) compared with those in L diet-fed mice, whereas LM hydrolysate reduced TG levels (dietary × LPS/Gal effect; P < .0001) by 1.51-fold in the H diet group (Fig. 2C). Similarly, LM hydrolysate lowered serum LDL-C levels (dietary × LPS/Gal effect; P < .05) by 1.38-fold compared with those in both the L and H diet groups (Fig. 2F).
LPS/Gal intervention influenced serum lipid levels, increasing HDL-C (LPS/Gal effect; P < .0001) by 2.37-fold and reducing TC (LPS/Gal effect; P < .0001) (Fig. 2D, E). This resulted in a higher HDL/LDL ratio (LPS/Gal effect; P < .0001) and reduced CRF values (LPS/Gal effect; P < .0001) (Fig. 2G, H). However, LM hydrolysate did not significantly affect the LPS/Gal-induced changes in either TC or HDL-C levels (Fig. 2D, E). These results suggest that LM hydrolysate may alleviate dyslipidemia by reducing TG and LDL-C levels, thereby mitigating the risk of MASLD. However, LM hydrolysate exhibited limited protective effects against ALI induced by LPS/Gal.
Effects of H diet, LPS/Gal challenge, and LM hydrolysate on serum cytokines and chemokines
LPS/Gal-induced ALI is accompanied by significant inflammatory responses, leading to elevated cytokine and chemokine levels. 46 Serum levels of TNF-α, IL-6, IL-1 beta (IL-1β), IL-10, and monocyte chemoattractant protein-1 increased by 33-, 1,500-, 23.5-, 41.5-, and 381-fold, respectively (Fig. 3A–E), compared with those in control mice, with all targets showing significance (LPS/Gal effect; P < .0001). These increases were solely attributable to LPS/Gal and were unaffected by dietary intervention or LM hydrolysate. Thus, LM hydrolysate did not alleviate the systemic inflammatory response associated with LPS/Gal-induced ALI.

Effects of an H diet, LM hydrolysate, and LPS/Gal challenge on serum cytokines and chemokines levels. Five-week-old male C57BL/6J mice were fed either an L diet or an H diet for 8 weeks. Concurrently, mice on the H diet received LM hydrolysate by gavage and were then treated with DPBS or LPS (50 μg/kg)/Gal (400 mg/kg).
Effects of H diet, LPS/Gal challenge, and LM hydrolysate on hepatic histology
Given the observed reduction in liver weight with LM hydrolysate, histological changes were examined. Before LPS/Gal injection, the liver appearance and histology were unaffected by the diet or LM hydrolysate intervention (Fig. 4A, B). After LPS/Gal injection, gross liver observations revealed pronounced redness, which was attenuated in LM hydrolysate-treated mice (Fig. 4A). Histologically, LPS/Gal caused extensive inflammatory cell infiltration and vacuolization, particularly in H diet-fed mice. LM hydrolysate reduced both inflammation and vacuolization (Fig. 4B). Liver histological scores increased by 3.39-fold with LPS/Gal treatment (LPS/Gal effect; P < .0001) and 1.61-fold with H diet (dietary effect; P < .01). LM hydrolysate reduced liver histological scores (dietary effect; P < .01) by 1.95-fold compared with those in the H diet group (Fig. 4C). These findings suggest that LM hydrolysate alleviates MASLD-associated liver damage and reduces the susceptibility to secondary damage caused by LPS/Gal.
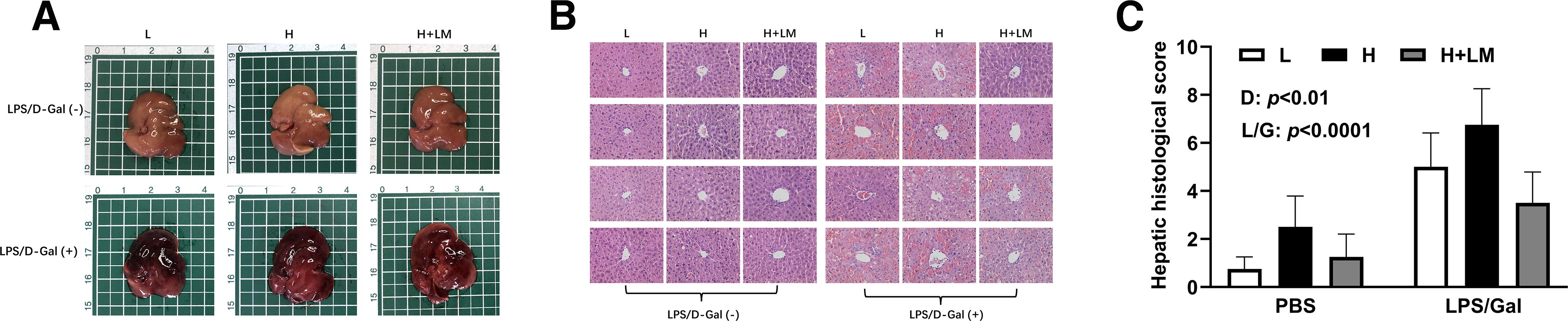
Effects of an H diet, LM hydrolysate, and LPS/Gal challenge on hepatic histology in C57BL/6J mice. Five-week-old male C57BL/6J mice were fed either an L diet or an H diet for 8 weeks. Concurrently, mice on the H diet received LM hydrolysate by gavage and were then treated with DPBS or LPS (50 μg/kg)/Gal (400 mg/kg).
Effects of H diet, LPS/Gal challenge, and LM hydrolysate on hepatic lipids and cytokines
To explore the inflammatory and lipid-modulating effects of LM hydrolysate, liver cytokine and lipid levels were analyzed. Dietary intervention increased hepatic TNF-α levels (dietary effect; P < .0001) by 1.21-fold, independent of LPS/Gal (Fig. 5A). LPS/Gal elevated IL-6 levels (LPS/Gal effect; P < .0001) by 2.53-fold, with H diet further increasing IL-6 levels (dietary effect; P < .01) by 1.37-fold compared with those in L diet-fed mice. After LPS/Gal administration, LM hydrolysate reduced IL-6 (dietary × LPS/Gal effect; P < .01) to levels similar to those in L diet-fed mice (Fig. 5B). In terms of hepatic lipids, the liver lipid weight was approximately 1.08-fold higher in mice fed the H diet than in mice fed the L diet, while LM hydrolysate lowered liver lipid levels (dietary effect; P < .01) by 1.25-fold compared with those in H diet-fed mice (Fig. 5C). These results suggest that LM hydrolysate reduces hepatic fat accumulation and inflammation, thereby alleviating MASLD.
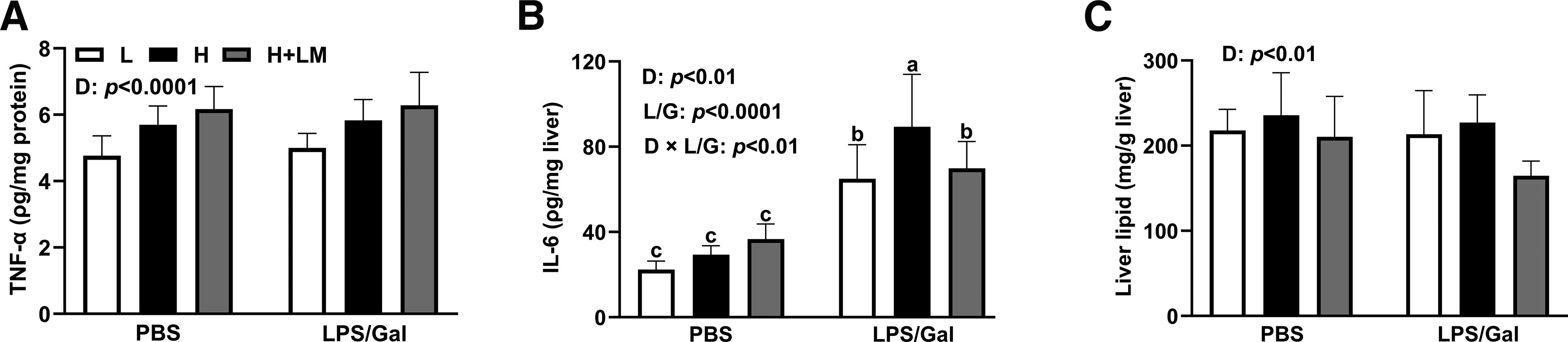
Effects of an H diet, LM hydrolysate, and LPS/Gal challenge on hepatic lipid profiles in C57BL/6J mice. Five-week-old male C57BL/6J mice were fed either an L diet or an H diet for 8 weeks. Concurrently, mice on the H diet received LM hydrolysate by gavage and were then treated with DPBS or LPS (50 μg/kg)/Gal (400 mg/kg).
Effects of H diet, LPS/Gal challenge, and LM hydrolysate on hepatic gene expression
The observed decreases in IL-6 levels and hepatic lipid contents in LM hydrolysate-treated mice suggest potential mechanisms through which LM hydrolysate mitigates MASLD. To explore these mechanisms, we analyzed the messenger RNA levels of genes associated with inflammation and lipid metabolism in the liver. LPS/Gal significantly increased the gene expression of Tnf-α (LPS/Gal effect; P < .0001), Il-1β (LPS/Gal effect; P < .0001), and Il-6 (LPS/Gal effect; P < .0001) by 8.95-, 96-, and 53.2-fold, respectively (Fig. 6A–C). Additionally, H diet elevated the expression of these genes by 1.85-(dietary effect; P < .05), 1.67-(dietary effect; P < .01), and 1.71-fold (dietary effect; P < .01), respectively (Fig. 6A–C). Notably, LM hydrolysate intervention reduced Tnf-α gene expression (dietary × LPS/Gal effect; P < .01) to levels comparable to those in L diet-fed mice under LPS/Gal condition, and similar reductions were observed for Il-6 (dietary × LPS/Gal effect; P < .01) (Fig. 6A, C). These results are consistent with previously observed reductions in IL-6 protein levels in the liver (Fig. 5B).

Effects of an H diet, LM hydrolysate, and LPS/Gal challenge on liver gene expression. Five-week-old male C57BL/6J mice were fed either an L diet or an H diet for 8 weeks. Concurrently, mice on the H diet received LM hydrolysate by gavage and were then treated with DPBS or LPS (50 μg/kg)/Gal (400 mg/kg). Relative hepatic
Furthermore, LM hydrolysate enhanced the expression of Hmox1 [encoding heme oxygenase-1 (HO-1)] in the absence of LPS/Gal injection, increasing it by 1.90-fold (dietary effect; P < .0001) compared with that in the L diet group and 2.48-fold (dietary effect; P < .0001) compared with that in the H diet group (Fig. 6D). After LPS/Gal injection, LM hydrolysate also enhanced Hmox1 gene expression by 2.99- and 1.99-fold compared with that in the L diet group (dietary × LPS/Gal effect; P < .0001) and H diet group (dietary × LPS/Gal effect; P < .0001), respectively. The current findings suggest that LM hydrolysate boosts antioxidant capacity and hence reduces inflammatory responses and prevents MASLD progression.
We also investigated the expression of genes regulating lipid metabolism. H diet reduced sterol regulatory element binding transcription factor 1 (Srebp1) expression (dietary effect; P < .0001) by 4.19-fold and fatty acid synthase (Fasn) expression (dietary effect; P < .001) by 3.1-fold, regardless of LPS/Gal intervention (Fig. 6E, G). To our surprise, the expressions of anabolic hepatic genes related to lipid metabolism were significantly attenuated in the H diet group compared with those in the L diet group. Although this reduction in lipogenic gene expression was unexpected, it may be attributed to compensatory responses aimed at counteracting the onset of fatty liver. In contrast, LPS/Gal stimulation increased acetyl-coenzyme A carboxylase beta (Acc) (LPS/Gal effect; P < .0001) and Fasn expression (LPS/Gal effect; P < .0001) by 29.5- and 25.7-fold, respectively, but reduced CCAAT/enhancer binding protein alpha (Cebp/α) gene expression (LPS/Gal effect; P < .0001) by 6.08-fold (Fig. 6F–H). This upregulation of lipid synthesis genes suggests a hepatic response to inflammatory damage, whereas reduction in Cebp/α expression indicates impaired hepatocyte regeneration.
Interestingly, LM hydrolysate reduced LPS/Gal- and H diet-induced Acc expression (dietary × LPS/Gal effect; P < .0001) by 2.26-fold compared with that in the H diet group (Fig. 6F). Furthermore, LPS/Gal increased the gene expression of the CD36 molecule (Cd36) (LPS/Gal effect; P < .0001) and acyl-coenzyme A oxidase 1, palmitoyl (Acox1) (LPS/Gal effect; P < .0001) (related to beta-oxidation) by 7.35- and 1.70-fold, respectively, whereas H diet elevated Acox1 gene expression (dietary effect; P < .01) by 1.82-fold, independent of LM intervention (Fig. 6I, J). These results suggest that LPS/Gal-induced ALI increases hepatic lipid demand, whereas LM hydrolysate primarily reduces Acc gene expression, alleviates hepatic fat accumulation, and prevents the pathological progression of MASLD.
Effects of H diet, LPS/Gal challenge, and LM hydrolysate on the expressions of proteins related to inflammation, endoplasmic reticulum stress, and oxidative stress
To examine the preventive effects of LM hydrolysate on MASLD, we assessed the expressions of proteins related to inflammation, oxidative stress, and endoplasmic reticulum (ER) stress. LPS/Gal increased the phosphorylation of nuclear factor kappa B (NF-κB) (LPS/Gal effect; P < .05) and c-Jun N-terminal kinase (JNK) (LPS/Gal effect; P < .05) by 1.67- and 1.80-fold, respectively, regardless of dietary intervention or LM hydrolysate treatment (Fig. 7A–C). Although LM hydrolysate reduced inflammatory cytokine levels in the liver, it did not significantly affect LPS/Gal-induced phosphorylation of NF-κB or JNK.
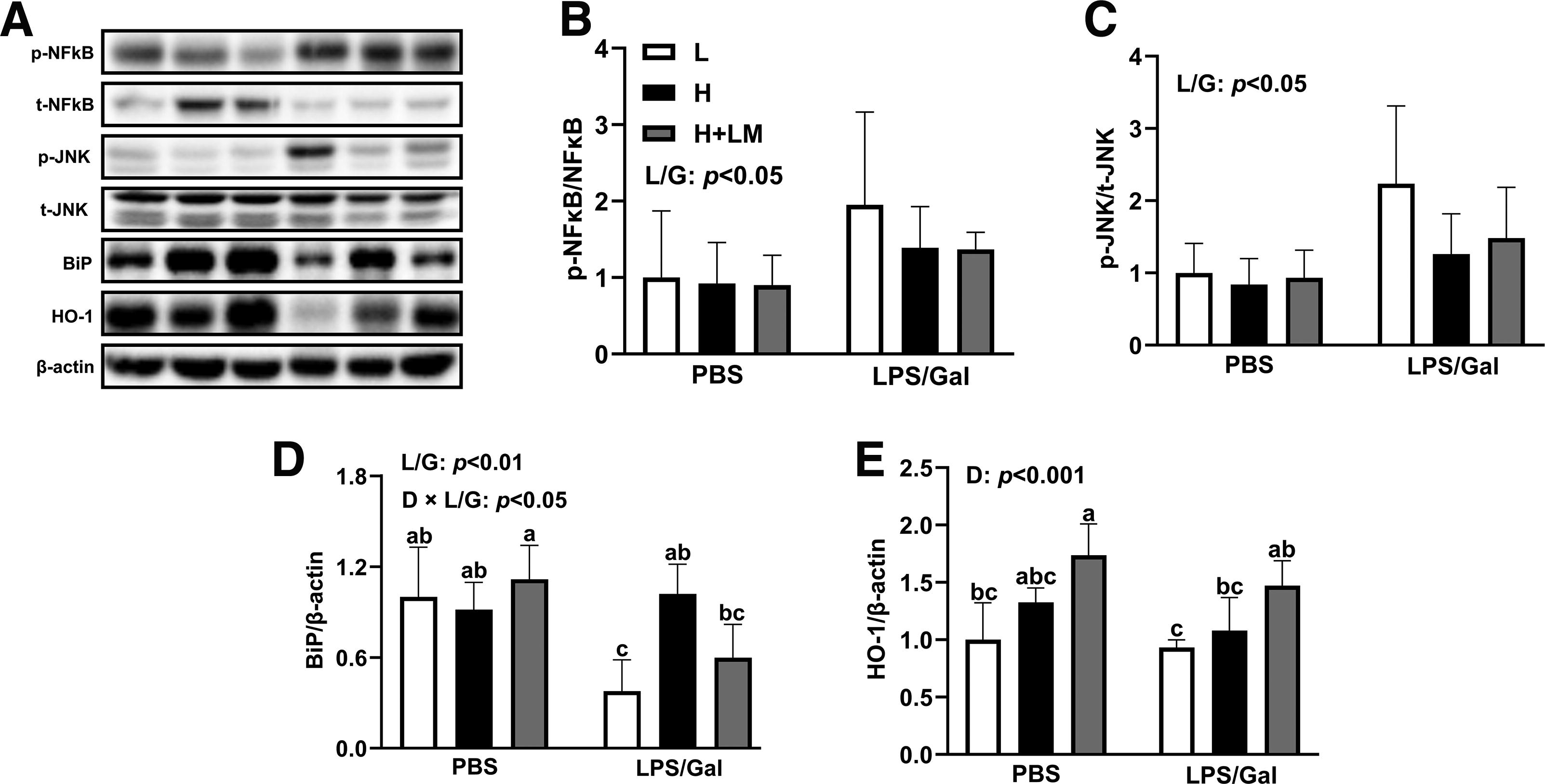
Effects of an H diet, LM hydrolysate, and LPS/Gal challenge on protein expression of inflammation, oxidative stress, and ER stress in C57BL/6J mice. Five-week-old male C57BL/6J mice were fed either an L diet or an H diet for 8 weeks. Concurrently, mice on the H diet received LM hydrolysate by gavage and were then treated with DPBS or LPS (50 μg/kg)/Gal (400 mg/kg).
LPS/Gal stimulation decreased the overall expression of binding immunoglobulin protein (BiP) (LPS/Gal effect; P < .01), a marker of ER stress. However, the combination of H diet and LPS/Gal intervention increased BiP expression (dietary × LPS/Gal effect; P < .05) by 2.7-fold compared with that in L diet-fed mice (Fig. 7A, D). LM hydrolysate treatment reduced BiP expression (dietary effect; P < .001) by 1.7-fold compared with that in the H diet group with LPS/Gal coadministration (Fig. 7A, D). Importantly, LM hydrolysate significantly increased HO-1 protein expression (dietary effect; P < .001) by 1.66- and 1.33-fold compared with that in the L diet and H diet groups, respectively, independent of LPS/Gal intervention (Fig. 7A, E). These findings suggest that LM hydrolysate alleviates MASLD progression by enhancing the antioxidant capacity and reducing ER stress, although its effects on inflammation in ALI are limited.
Effects of H diet, LPS/Gal challenge, and LM hydrolysate on liver oxidative stress markers
To further investigate the antioxidant effects of LM hydrolysate, we analyzed markers of oxidative stress in the liver. LPS/Gal significantly increased MDA levels by 4.57-fold (Fig. 8A) and decreased SOD, GSH, and GSSG levels by 1.36-, 2.69-, and 4.23-fold, respectively (Fig. 8B–D). These changes were independent of dietary intervention, with all targets showing significance (LPS/Gal effect; P < .0001). H diet of LM further reduced GSH (dietary effect; P < .0001) and GSSG (dietary effect; P < .0001) levels by 1.55- and 1.63-fold, respectively, regardless treatment (Fig. 8C, D). Notably, LM hydrolysate increased hepatic SOD levels (dietary effect; P < .01) by 1.29-fold compared with those in the H diet group (Fig. 8B). Although LM hydrolysate did not directly alter GSH or GSSG levels, it significantly improved the GSH/GSSG ratio (dietary × LPS/Gal effect; P < .05), increasing it by 1.38- and 1.56-fold compared with those in the L diet- and H diet-fed mice (Fig. 8E). These findings suggest that LM hydrolysate enhances hepatic antioxidant defenses, alleviates MASLD, and mitigates secondary damage caused by LPS/Gal under MASLD conditions.

Effects of an H diet, LM hydrolysate, and LPS/Gal challenge on liver oxidative stress markers in C57BL/6J mice. Five-week-old male C57BL/6J mice were fed either an L diet or an H diet for 8 weeks. Concurrently, mice on the H diet received LM hydrolysate by gavage and were then treated with DPBS or LPS (50 μg/kg)/Gal (400 mg/kg). Liver
DISCUSSION
This study evaluated the hepatoprotective effects of LM enzymatic hydrolysate against MASLD and ALI induced by LPS/Gal in an MASLD mouse model. An 8‐week intervention was selected because an H diet for <20 weeks leads to liver steatosis and increased lipid accumulation without severe inflammation or fibrosis, 47 allowing assessment of the effects of LM hydrolysate during the early phase of MASLD. MASLD was induced in model animals with a high‐fat, high‐fructose, and high‐cholesterol diet that produced fat degeneration, lobular inflammation, hepatocyte ballooning, fibrotic lesions, and alterations in the hepatic transcriptome. Concurrently, mice received LM hydrolysate via daily oral gavage, and ALI was subsequently induced by LPS/Gal. The findings demonstrated that LM hydrolysate prevented MASLD and alleviated liver dysfunction associated with chronic metabolic stress, while its capacity to mitigate LPS/Gal‐induced ALI remained minimal. This highlights a disparity in efficacy between chronic and severe acute injury.
Edible insects have been reported to possess anti-inflammatory, antioxidant, antihypertensive, and other bioactivities, showing potential in preventing and mitigating MetS and related diseases. 48 LM is particularly noteworthy for its high protein content (∼50–65%), abundant unsaturated fatty acids, and polyphenolic compounds, which contribute significantly to its nutritional and functional value. 49 Nutrient density in LM is largely attributed to its natural diet, which includes various grasses and crops during migration. 50 Moreover, the combination of these nutritional components supports various metabolic processes and contributes to overall health benefits, reinforcing the role of LM in the management of MetS and related disorders.
Protein hydrolysis enables new and novel food applications by altering the biological, nutritional, or functional properties of proteins, thereby improving antioxidant, antihypertensive, antimicrobial, and anticancer effects. 51 Enzymatic hydrolysis of food by-products also promotes effective waste utilization, reduces waste, and generates valuable bioactive compounds. 52 Edible insect protein hydrolysis enhances its bioactivity, widely demonstrating antihypertensive, antidiabetic, and antioxidant properties. 53 Furthermore, the chitinous exoskeleton of LM may limit nutrient bioavailability. 54 However, hydrolysis releases retinol, vitamins, and carotenoids and may produce peptides with anti-inflammatory and antioxidant activities. 11 Recent studies have also confirmed that LM hydrolysates can alleviate dyslipidemia and improve hepatic lipid metabolism in rats, which may be attributed to their high content of total phenols, carotenoids, and excellent DPPH radical scavenging activity. 31 However, further research will be required to confirm the specific bioactive compounds responsible for these observed effects.
The Western diet is widely used in diet-induced obesity (DIO) models to investigate the pathophysiological mechanisms underlying MASLD, which is essential for developing therapeutic strategies. 55 Modifications to DIO diets exacerbate MASH in murine models; a high-fat and high-fructose diet supplemented with cholesterol induces hepatic lobular inflammation and hepatocyte ballooning. 56 After 8 weeks of dietary intervention, H diet-fed mice exhibited significantly higher body weights than did L diet-fed mice, with differences becoming apparent after 4 weeks (Fig. 1A). Additionally, H diet significantly increased total WAT weights, regardless of LPS/Gal or LM hydrolysate intervention (Fig. 1H). H diet and LPS/Gal significantly elevated the liver weight, whereas LM hydrolysate mitigated this increase (Fig. 1D).
Dyslipidemia is a hallmark of MetS, contributing to increased risk of cardiovascular diseases, diabetes, and MASLD. 57 Elevated TG, TC, and LDL-C levels promote hepatic fat accumulation, triggering liver damage and inflammation associated with MASLD. 58 Serum lipid analysis revealed that LPS/Gal decreased TC levels, increased HDL-C levels, and improved the HDL-C/LDL-C ratio, regardless of diet or LM hydrolysate intervention (Fig. 2D, E, G). Elevated TG and LDL-C levels disrupt lipid homeostasis and play a pivotal role in the onset and progression of MASLD, with LM hydrolysate treatment reducing TG levels and lowering LDL-C compared with those in the L and H diet groups (Fig. 2C, F).
Patients with MASLD frequently develop dyslipidemia. A meta-analysis of 8,515,431 patients with MASLD from 22 countries reported dyslipidemia prevalence rates of 69% and 72% in patients with MASLD and MASH, respectively. 59 The liver regulates fatty acid metabolism via fatty acid absorption and release, anabolic de novo synthesis, and catabolic β-oxidation. 60 LM hydrolysate reduced hepatic lipid accumulation (Fig. 5C), prompting an investigation of related molecular mechanisms. H diet reduced Srebp1 and Fasn gene expression (Fig. 6E, G), likely due to a negative feedback response to hepatic lipid accumulation, whereas LPS/Gal elevated Acc and Fasn expression and reduced Cebp/α expression (Fig. 6F, G, H).
LPS/Gal significantly elevated the gene expression of Cd36 and Acox1, which are key markers of catabolic lipid utilization via β-oxidation (Fig. 6I, J). LM hydrolysate effectively mitigated the LPS/Gal- and H diet-induced increases in Acc gene expression (Fig. 6F), improved anabolic lipid metabolism, reduced hepatic lipid accumulation, and downregulated genes associated with fatty acid synthesis. The high protein content of LM hydrolysate likely provides essential amino acids that promote lipid metabolism, whereas its bioactive polyphenols may regulate hepatic steatosis through anti-inflammatory pathways. Excess hepatic fat accumulation in MASLD is known to induce oxidative and ER stress, triggering inflammation and apoptosis. 61
The combination of H diet and LPS/Gal elevated hepatic TNF-α and IL-6 protein levels, whereas LM hydrolysate significantly reduced the levels of cytokines as well as their respective genes (Fig. 5B; Fig. 6A, C). Additionally, LM hydrolysate increased the expression of Hmox1 and HO-1 proteins, thereby enhancing the hepatic antioxidant capacity (Fig. 6D; Fig. 7A). LPS/Gal elevated MDA, a marker of oxidative stress, but reduced the levels of the antioxidant markers SOD, GSH, and GSSG (Fig. 8A–D). Importantly, LM hydrolysate restored SOD levels and improved the GSH/GSSG ratio, thereby reducing oxidative stress and alleviating MASLD (Fig. 8B, E).
Despite these benefits, LM hydrolysate showed limited efficacy in preventing ALI, likely due to the severity of the LPS/Gal model, which is characterized by acute inflammation and extensive hepatocyte necrosis. 62 The excessive inflammatory response in this model may surpass the protective capacity of LM hydrolysate, which seems more effective in managing chronic liver conditions. Future studies should focus on identifying the specific bioactive components of LM hydrolysate, evaluating its efficacy in less severe ALI models, and exploring its potential in combination with other antioxidants or anti-inflammatory agents to enhance its therapeutic benefits.
CONCLUSION
This study highlights the hepatoprotective potential of LM hydrolysate in mitigating diet-induced MASLD and the associated liver pathology. Our findings demonstrated that LM hydrolysate significantly alleviated MASLD by alleviating dyslipidemia, reducing hepatic fat accumulation, and attenuating liver inflammation and oxidative stress. Specifically, LM hydrolysate administration lowered serum TG and LDL-C levels, decreased hepatic TNF-α and IL-6 concentrations, and increased liver HO-1 expression, indicating a protective role against liver dysfunction. Mechanistically, LM hydrolysate enhanced antioxidant capacity and modulated lipid metabolism while suppressing inflammatory cytokine production. Although LM hydrolysate did not effectively prevent LPS/Gal-induced ALI, it significantly mitigated MASLD and reduced the susceptibility to ALI under MASLD conditions. These results underscore the potential of LM hydrolysate as a functional food or dietary supplement for MASLD management. Future research should focus on identifying and characterizing the specific bioactive compounds in LM hydrolysate and evaluating their efficacy in clinical trials to fully harness their therapeutic potential.
Footnotes
AUTHORS’ CONTRIBUTIONS
J.Z.: Conceptualization, data curation, formal analysis, investigation, methodology, software, validation, visualization, and writing—original draft. S.J.: Conceptualization, data curation, investigation, methodology, software, validation, visualization, supervision, and project administration. J.-H.H.: Conceptualization, formal analysis, methodology, visualization, writing—original draft, resources, writing—review and editing, and supervision. Y.J.: Conceptualization, data curation, resources, supervision, project administration, and funding acquisition. All authors have read and agreed to the published version of the article.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
No funding was received for this article.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.