
Abstract
Although probiotic products are widely used to promote gut health, their long-term efficacy is often constrained by poor colonization and transient microbial modulation. To address these limitations, we developed and evaluated a probiotic beverage comprising Lactobacillus-based strains (Lactobacillus rhamnosus LGD-0817, L. rhamnosus LDL-1708, Lactobacillus casei LPM-0188, Lactiplantibacillus plantarum LEP-1208, Lactobacillus delbrueckii subsp. bulgaricus RHM-1068, Lactobacillus paracasei LPA-2088, L. paracasei BMC-2020, Limosilactobacillus fermentum LAF-1387, and L. plantarum MCA-3066) formulated to optimize inter-strain synergy, enhance colonization efficiency, and improve long-term functionality. 16S rRNA sequencing conducted one week after treatment cessation revealed a marked enrichment of Lactobacillales and Lactobacillaceae—taxonomic groups to which administered probiotic strains belong—indicating durable colonization by the introduced strains. To further assess the functional relevance of this microbial shift, a dextran sulfate sodium-induced colitis model was employed. Probiotic intervention significantly improved disease outcomes, including increased colon length, reduced histopathological inflammation scores, enhanced goblet cell numbers, and lowered levels of pro-inflammatory cytokines (IL-6, Tumor Necrosis Factor-alpha (TNF-α), and Interleukin (IL)-1β). Collectively, these results underscore the efficacy of this formulation in achieving sustained modulation of the gut microbiota and alleviating intestinal inflammation, and support its potential for broader applications in gut health management.
INTRODUCTION
The gut microbiota has become a central focus of academic research and public interest due to its critical role in maintaining overall human health. 1 –3 Studies have shown that the gut microbiota facilitates digestion, absorption, and nutrient metabolism and plays a key role in immune regulation, anti-inflammatory responses, and antioxidation processes. 4,5 Beneficial gut microbes help maintain ecological balance by suppressing harmful bacteria and producing health-promoting compounds. Disruption of this balance is linked to various health issues, including digestive disorders, impaired immunity, and cardiovascular diseases. 6,7
Probiotics, particularly those from the Lactobacillus genus, have been extensively studied for their broad spectrum of beneficial effects on gut health. These effects include restoring microbial balance, enhancing immune function, aiding digestion, producing bioactive metabolites, exhibiting anti-inflammatory and antimicrobial activities, strengthening gut barrier integrity, and regulating intestinal motility. 8 –10 Given the growing recognition of the gut microbiota’s role in overall health, probiotics have become a key intervention for modulating gut microbiota and promoting intestinal well-being. 11
Probiotic beverages have become a widely recognized approach for promoting gut health, offering a convenient and effective method to deliver beneficial microbes. These beverages often feature multistrain formulations that combine selected probiotic strains to harness synergistic effects. 12,13 Unlike single-strain probiotics, multistrain formulations provide a broader spectrum of activity, enhancing their ability to modulate gut microbiota, restore microbial balance, and support intestinal health. This combination strategy has made probiotic beverages a promising tool for addressing gut health challenges. Despite these advances, current probiotic beverages still face significant limitations. A major challenge is the inefficient colonization of probiotics in the gut. Many probiotic strains fail to establish a stable presence due to competition with existing gut microbiota, exposure to gastric acid and bile salts, and individual variations in host factors. 14 Additionally, the long-term efficacy of these beverages is often insufficient, as the delivered probiotics may fail to maintain their beneficial effects without continuous consumption.
To address these shortcomings, we developed an optimized probiotic formulation comprising Lactobacillus rhamnosus LGD-0817, L. rhamnosus LDL-1708, Lactobacillus casei LPM-0188, Lactiplantibacillus plantarum LEP-1208, Lactobacillus delbrueckii subsp. bulgaricus RHM-1068, Lactobacillus paracasei LPA-2088, L. paracasei BMC-2020, Limosilactobacillus fermentum LAF-1387, and L. plantarum MCA-3066. Its effects on the gut microbiota were evaluated by 16S rRNA sequencing one week after treatment cessation, and its anti-inflammatory activity was examined in a dextran sulfate sodium (DSS)-induced colitis model. The results demonstrate that this formulation enables sustained modulation of the gut microbiota and alleviates intestinal inflammation, highlighting its potential for gut health management.
MATERIALS AND METHODS
Materials
The probiotic beverage used in this study was developed in collaboration with Zhongweiweiyuan Biotechnology Co., Ltd. (Anhui, China) and is commercially available under the name Yieryou®. The ingredients include water, isomaltulose, malt extract, Lactobacillus-based probiotics (108 CFU/mL, comprising L. rhamnosus LGD-0817, L. rhamnosus LDL-1708, L. casei LPM-0188, L. plantarum LEP-1208, L. delbrueckii subsp. bulgaricus RHM-1068, L. paracasei LPA-2088, L. paracasei BMC-2020, L. fermentum LAF-1387, and L. plantarum MCA-3066), sodium citrate, xanthan gum, vitamin C, potassium sorbate, and sucralose.
Reagents
DSS (catalog number HY-116282C) was purchased from MedChemExpress. Hematoxylin-eosin (HE) staining kit (catalog number G1120) and periodic acid-Schiff (PAS) staining kit (catalog number G1281) were purchased from Solarbio. Enzyme-Linked Immunosorbent Assay (ELISA) kits for IL-1β (catalog number BMS6002-2TEN), IL-6 (catalog number BMS603-2), and TNF-α (catalog number BMS607-3FIVE) were purchased from Thermo Fisher Scientific.
Animals
Five-week-old male C57BL/6J mice were purchased from Tsinghua University Experimental Animal Center and housed in an SPF-grade animal facility under controlled humidity (50 ± 5%), temperature (22 ±2°C), and a 12-h light-dark cycle. After one week of adaptive feeding, the mice were randomly divided into two groups (n = 6 each): the control group (CON) and the probiotic beverage group (YEY). The YEY group received 100 µL of the probiotic beverage via oral gavage daily for 30 days, while the CON group received an equal volume of water.
This study was approved by the Institutional Animal Care and Use Committee of Tsinghua University (Approval No: THU-LARC-2024-020) and was conducted in accordance with the ARRIVE guidelines. All animal procedures were carried out in accordance with relevant guidelines and regulations.
16S rRNA sequencing
One week after completing the probiotic gavage, total DNA was extracted from mouse fecal samples, and the hypervariable region (V3-V4) of the 16S rRNA gene was amplified via PCR (each group n = 6). The amplified products were purified, used to construct sequencing libraries, and sequenced on the Illumina MiSeq high-throughput platform. The raw sequences underwent quality control, assembly, and clustering, followed by denoising to generate amplicon sequence variants (ASVs). Taxonomic annotation was performed by comparing the ASVs with a reference database. Subsequent data analyses included alpha diversity and beta diversity.
DSS-induced colitis
Five-week-old male C57BL/6J mice were adapted for one week and then randomly divided into four groups (n = 6 each): CON group, DSS group, YEY group, and YEY+DSS group. All groups underwent 30 days of oral gavage. The YEY and YEY+DSS groups received 100 µL of the probiotic beverage daily, while the CON and DSS groups received equal volumes of water. After 30 days of gavage, the DSS and YEY+DSS groups were given 3% DSS solution as drinking water ad libitum for seven consecutive days to induce colitis.
HE staining and PAS staining
Colon tissue samples were fixed in 4% paraformaldehyde for 24 h and then dehydrated in 70% ethanol. A series of graded ethanol solutions (70%, 80%, 90%, 95%, and 100%) was used for further dehydration, followed by clearing with xylene. The dehydrated tissues were embedded in paraffin and sectioned into 5 µm-thick slices. The slices were stained according to the protocols of hematoxylin-eosin (HE) and PAS staining kits and observed under a microscope.
HE Staining: the slices were deparaffinized in xylene twice for 10 min each, rehydrated in graded ethanol solutions (100%, 95%, 85%, and 75%) for 3 min per gradient, and rinsed in distilled water for 2 min. They were then stained with hematoxylin for 10 min, washed in distilled water, differentiated in differentiation solution for 30 sec, and rinsed twice in tap water for 3 min each. Eosin staining was applied for 1 min, and the slices were quickly dehydrated. The dehydration process included sequential rinses in 75%, 85%, 95%, and 100% ethanol for 3 sec each, followed by 1 min in 100% ethanol. The slices were then cleared in xylene twice for 1 min each and mounted with neutral resin.
PAS Staining: the slices were deparaffinized in xylene twice for 10 min each, rehydrated in graded ethanol solutions (100%, 95%, 85%, and 75%) for 3 min per gradient, and rinsed in distilled water for 2 min. They were treated with periodic acid for 5 min to oxidize polysaccharides into aldehydes and then rinsed three times in distilled water for 1 min each. The slices were stained with Schiff reagent for 10 min to form red-purple complexes with the aldehydes, rinsed under running water for 5 min to remove unreacted Schiff reagent, and dehydrated through sequential rinses in 75%, 85%, 95%, and 100% ethanol for 3 sec each, followed by 1 min in 100% ethanol. Finally, the slices were cleared in xylene twice for 1 min each and mounted with neutral resin.
Colitis inflammation scoring
The degree of intestinal inflammation was evaluated based on HE staining results, using a five-point scale: no inflammation (0 points), slight infiltration of cells in the lamina propria (1 point), monocyte infiltration with crypt separation and mild mucosal hyperplasia (2 points), extensive inflammatory cell infiltration with mucosal structure damage, goblet cell loss, and significant mucosal hyperplasia (3 points), and severe inflammation with crypt abscesses or ulceration (4 points). Each section was assessed blindly, selecting five random fields of view per section. The average score of the fields was used as the final inflammation score for the section. Higher scores indicate more severe inflammation.
ELISA
Colon tissues (100 mg) were homogenized in 100 µL of Radioimmunoprecipitation Assay (RIPA) lysis buffer containing protease inhibitors. The homogenate was centrifuged at 13500 x g at 4°C, and the supernatant was collected as the total protein sample. ELISA kits were used to measure the protein levels of IL-1β, IL-6, and TNF-α in the supernatant. The results were normalized to the total weight of the colon tissue and expressed as pg/mg.
Statistical analysis
GraphPad Prism 7 (Graphpad software) was used to assess statistical significance. The data are presented as x̄ ± s. One-way analysis of variance was used for comparisons among multiple groups, followed by Tukey’s post hoc test. P < .05 was considered statistically significant.
RESULTS
Effects of the probiotic beverage on body weights
Following adaptive period, the baseline body weights of the mice were recorded. Subsequent weight measurements were taken on days 10, 20, and 30 of the gavage treatment. As shown in Table 1, there were no significant differences in body weight between the YEY group and the CON group at any stage of the gavage period. These results indicate that the probiotic beverage did not have a noticeable impact on the growth and development of the mice.
Effects of the Probiotic Beverage on Body Weights
CON, control group; YEY, probiotic beverage group.
Effects of the probiotic beverage on gut microbiota
One week after the cessation of the probiotic treatment, 16S rRNA sequencing was conducted to assess its impact on gut microbiota composition in mice. As shown in Figure 1, the YEY and CON groups shared 571 ASVs, with 18 unique to the YEY group and 60 unique to the CON group, suggesting the emergence of group-specific microbial signatures following intervention.

Venn diagram of ASV between groups. ASV, amplicon sequence variant.
Within-sample microbial diversity was evaluated using standard alpha diversity indices, including Shannon, Simpson, Chao1, Goods Coverage, and PD Whole Tree. As summarized in Table 2, none of these indices differed significantly.
Alpha Diversity Statistics Based on Amplicon Sequence Variant
While overall alpha diversity remained comparable, taxonomic profiling based on relative abundance revealed compositional differences between groups. As shown in Figure 2A, Firmicutes and Bacteroidota remained the dominant phyla in both cohorts; however, probiotic treatment in the YEY group led to a relative reduction in Firmicutes, along with modest but discernible increases in Campylobacterota, Actinobacteriota, and Deferribacterota. At the genus level (Fig. 2B), microbial composition in the CON group was largely dominated by unclassified taxa grouped under the “Others” category. In contrast, the YEY group exhibited a higher proportion of taxonomically defined genera. Most notably, Lactobacillus abundance increased from 0.038% in the CON group to 2.67% in the YEY group, representing an approximate 70-fold elevation. Additionally, Ligilactobacillus and Helicobacter appeared at higher relative levels in the YEY group, while modest increases were also observed for Mucispirillum and Dubosiella.

Relative abundance of gut microbiota.
To statistically validate these compositional shifts, linear discriminant analysis effect size was performed. As shown in Figure 3A and B, the YEY group exhibited significant enrichment of Lactobacillales (order) and Lactobacillaceae (family)—taxonomic groups to which all nine administered probiotic strains belong—indicating successful colonization by the introduced microbes. In contrast, the CON group showed enrichment of Lachnospiraceae bacterium A2, unidentified_Lachnospiraceae, Lachnospirales, and Lachnospiraceae, suggesting a shift away from these resident taxa following probiotic intervention.
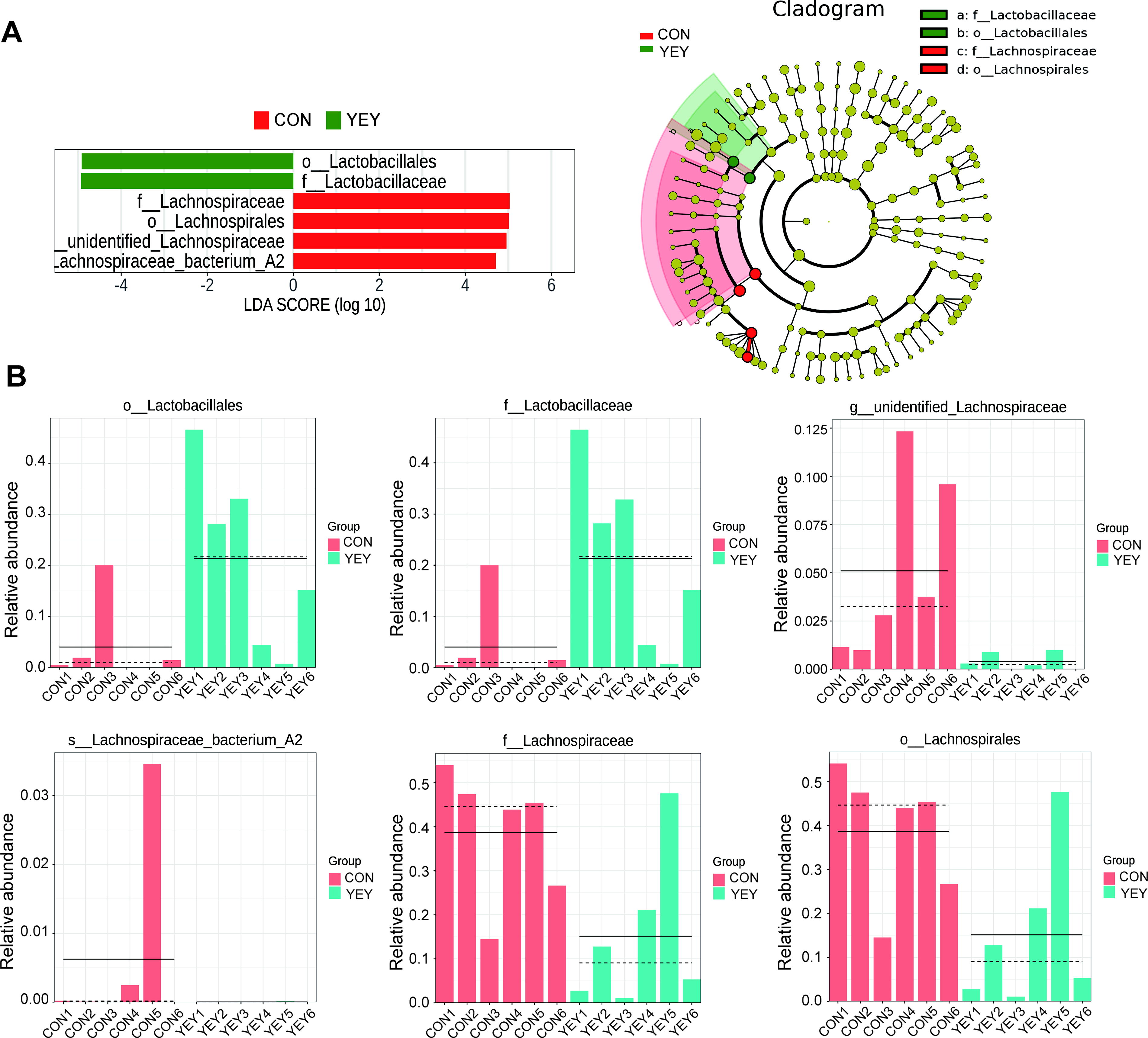
LEfSe analysis.
Amelioration of DSS-induced colitis by the probiotic beverage
In light of the enrichment of Lactobacillales and Lactobacillaceae in the YEY group—taxa that include several strains previously reported to possess immunoregulatory and anti-inflammatory properties—we next sought to determine whether the probiotic beverage could alleviate DSS-induced colitis. To this end, mice were administered the probiotic beverage for 30 days, followed by a 7-day induction of colitis using 3% DSS. As shown in Figure 4A and B, the colon length of DSS-treated mice was significantly reduced by approximately 20.2% compared with the CON group (P < .01). However, the colon length in the YEY+DSS group was significantly increased by approximately 10.6% compared with the DSS group (P < .05), indicating that the probiotic beverage alleviates DSS-induced colonic shortening.

Colon length in mice.
As shown in Figure 5A, the HE staining results indicate severe structural disruption in the colonic mucosal layer of the DSS group. The glandular arrangement was irregular, with some glands missing or degenerating, and the mucosal layer exhibited significant atrophy or detachment. Extensive inflammatory cell infiltration was observed, particularly in the submucosal and muscular layers, accompanied by ulcer formation and tissue necrosis in certain areas. In contrast, the colonic tissue structure in the YEY+DSS group showed notable improvement. Statistical analysis revealed that the histological inflammation score of the YEY+DSS group was approximately 27.4% lower than that of the DSS group (P < .05) (Fig. 5B).

HE staining results of mice colon.
Goblet cells are a critical cell type in the intestinal epithelium, primarily responsible for secreting mucus that forms a protective mucus layer lining the intestinal wall. A reduction in goblet cells leads to decreased mucus secretion, causing the mucus layer to thin or disappear. This damage weakens the intestinal barrier’s defense function, increasing the exposure of epithelial cells to pathogens and antigens, thereby exacerbating intestinal inflammation. As shown in Figure 6A and B, the number of goblet cells (stained magenta) in the DSS group was significantly reduced by approximately 66.4% compared with the CON group (P < .01). By comparison, the YEY+DSS group showed a significant increase in goblet cell numbers by approximately 55.9% compared with the DSS group (P < .05).
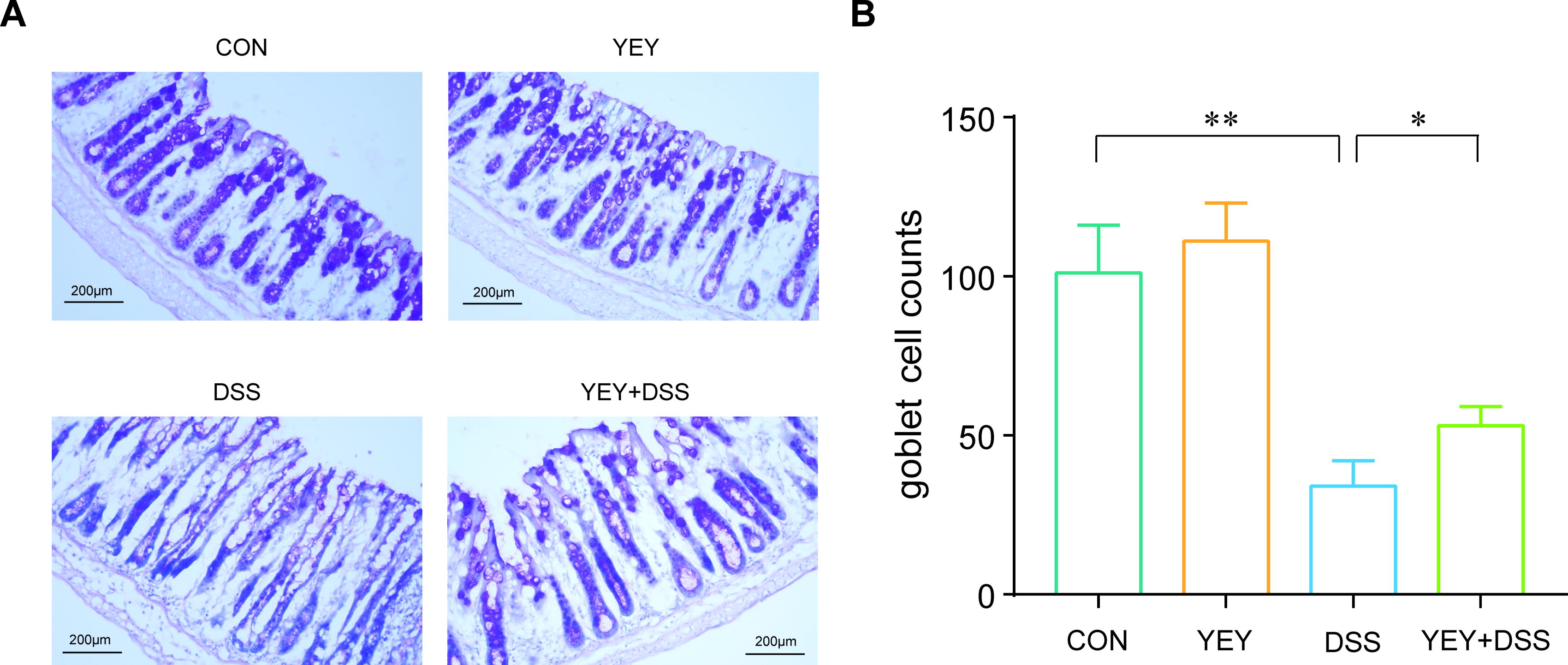
PAS staining results of mice colon.
As shown in Figure 7A–C, the levels of three pro-inflammatory cytokines—IL-6, TNF-α, and IL-1β—were significantly elevated in the DSS group compared with the CON group, with increases of approximately 107.3% (P < .01), 77.8% (P < .01), and 140.1% (P < .01), respectively. In contrast, the YEY+DSS group showed a significant reduction in these cytokine levels compared with the DSS group, with decreases of approximately 28.3% (P < .01), 25.3% (P < .01), and 19.8% (P < .01), respectively.
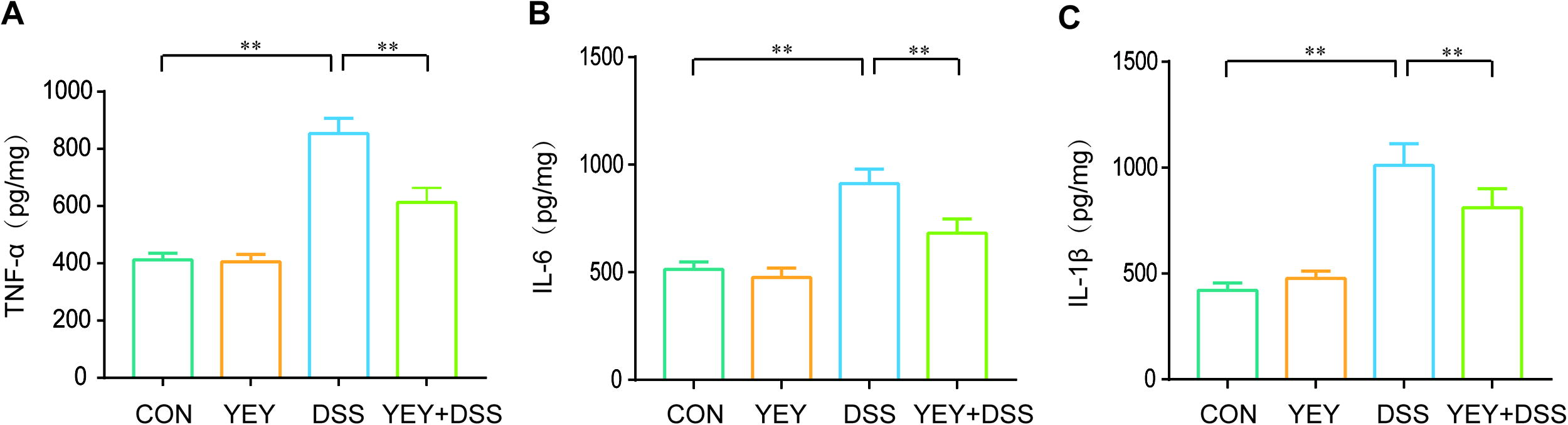
Inflammatory cytokine levels in mice colon.
DISCUSSION
Probiotics have emerged as a promising strategy to beneficially alter gut microbial ecosystems and improve host outcomes. However, their long-term efficacy is often constrained by poor colonization and transient microbial changes. In this study, we evaluated an optimized, multistrain Lactobacillus-based probiotic formulation—comprising L. rhamnosus LGD-0817, L. rhamnosus LDL-1708, L. casei LPM-0188, L. plantarum LEP-1208, L. delbrueckii subsp. bulgaricus RHM-1068, L. paracasei LPA-2088, L. paracasei BMC-2020, L. fermentum LAF-1387, and L. plantarum MCA-3066—that was formulated to optimize inter-strain synergy, enhance colonization efficiency, and improve long-term functionality.
Following 30 days of oral gavage and a subsequent 1-week washout period, 16S rRNA sequencing revealed a targeted modulation of gut microbial composition. A significant enrichment of Lactobacillales and Lactobacillaceae was observed in the probiotic-treated group, suggesting durable colonization by the introduced strains or stimulation of closely related endogenous taxa. Although there were no statistically significant differences in alpha diversity metrics, the observed compositional shifts suggest that the probiotic’s efficacy may depend more on the selective enrichment of specific taxa than on overall microbial richness. Notably, a relative depletion of Lachnospiraceae—a family containing taxa previously reported in inflamed intestinal environments 15 —was also detected, suggesting a compositional transition away from pro-inflammatory microbial signatures. These microbiota-level alterations were accompanied by physiological improvements in a DSS-induced colitis model. Compared with CON, mice receiving the probiotic exhibited increased colon length, reduced histopathological scores, recovery of goblet cell numbers, and lowered concentrations of pro-inflammatory cytokines (TNF-α, IL-6, and IL-1β). These findings suggest that the formulation not only remodeled gut microbial composition but also conferred protective effects on mucosal architecture and inflammatory responses under pathological conditions.
These beneficial effects are likely driven by the synergistic and complementary functionalities of the probiotic strains, which together contribute to durable colonization and intestinal protection. For instance, L. rhamnosus exhibits strong adhesion to intestinal epithelial cells, boosting colonization efficiency. 16 –18 L. casei produces antimicrobial peptides that foster competitive exclusion of pathogens and support a favorable niche for other strains. 19 –21 L. plantarum generate short-chain fatty acids, crucial for maintaining intestinal pH and nourishing epithelial cells, while L. fermentum enhances oxidative stress resistance through the production of antioxidants. 22 –26 L. delbrueckii subsp. bulgaricus contributes to fermentative processes, generating metabolites with anti-inflammatory properties, and L. paracasei is known for their immunoregulatory effects, stimulating anti-inflammatory cytokine production and enhancing mucosal immunity. 27 –30
Despite these encouraging findings, several limitations should be acknowledged. First, the investigation was conducted in a murine model, which, while widely used for preclinical evaluation, may not fully account for the complexity of human gut physiology and microbiota-host interactions. Differences in diet, immune regulation, and microbial composition across species could influence the colonization dynamics and therapeutic efficacy of the administered probiotics. Second, although the probiotic formulation consisted of nine well-characterized Lactobacillus strains, their individual contributions to the observed effects remain undetermined. The collective benefits are likely the result of synergistic interactions, but further studies employing strain-specific tracking methods are needed to dissect the functional hierarchy. Third, the study primarily focused on taxonomic alterations based on 16S rRNA gene sequencing, which provides limited resolution for functional inference. Future investigations integrating metagenomics, metabolomics, and host transcriptomics would enable deeper mechanistic insight into microbial function and host-microbe interactions. Finally, a one-week washout period may not be sufficient to establish a long-lasting change in the gut microbiota. Extended follow-up studies are needed to evaluate the durability of microbial shifts and their relevance to long-term gut health.
CONCLUSION
This study highlights the efficacy of this optimally designed probiotic formulation in achieving sustained gut microbiota modulation and alleviation of intestinal inflammation, and supports its potential for long-term gut health management.
Footnotes
ACKNOWLEDGMENT
The authors thank Zhongweiweiyuan Biotechnology Co., Ltd. for providing the probiotic beverage used in this study.
AUTHORS’ CONTRIBUTIONS
Conceptualization: H.Z. and W.H. Methodology: H.Z., Y.L., and W.H. Software: H.Z. Validation: W.H. Formal analysis: H.Z. and W.H. Investigation: H.Z., Y.L., and W.H. Resources: Q.G. and B.Z. Data curation: H.Z. and W.H. Writing—Original draft preparation: H.Z. and W.H. Writing—review and editing: H.Z. and W.H. Visualization: H.Z. and W.H. Supervision: B.Z. and W.H. Project administration: B.Z. and W.H. Funding acquisition: Q.G., B.Z., and W.H.
AUTHOR DISCLOSURE STATEMENT
The authors declare a collaborative relationship with Zhongweiweiyuan Biotechnology Co., Ltd. in the development of the probiotic beverage used in this study. The company provided financial support but had no role in the study design, data collection, data analysis, interpretation of results, or preparation of the article. The authors declare no competing financial interests.
FUNDING INFORMATION
This study was financially supported by Zhongweiweiyuan Biotechnology Co., Ltd.