
Abstract
Skeletal muscles, attached to bones via tendons, enable physical movement in animals. Muscle atrophy is associated with a decline in muscle mass and function and can detrimentally impact quality of life, leading to muscle weakness, altered fiber properties, and, ultimately, sarcopenia. Phlomis umbrosa Turcz. (Korean name: Han-Sok-Dan) has been used as a herbal medicine in Southeast Asia since ancient times. Although several studies have explored the properties of Han-Sok-Dan, further research is warranted to establish its therapeutic benefits for muscle function. This study investigated the effects of Phlomis umbrosa Turcz. water extract (PUW) in mice subjected to dexamethasone-induced muscle atrophy. Mice exhibiting dexamethasone-induced muscle atrophy experienced weight loss, reduced muscle mass, and functional decline. However, PUW administration effectively reversed these effects by maintaining muscle mass and strength, decreasing protein degradation-related marker expression, and enhancing signaling for protein synthesis. Notably, phosphorylations of mammalian target of rapamycin (mTOR), ribosomal protein S6 kinase beta-1, and eukaryotic translation initiation factor 4E-binding protein 1 were significantly enhanced in the PUW group, indicating the activation of anabolic signaling. Overall, PUW alleviates muscle atrophy induced by dexamethasone by modulating the balance between protein degradation and synthesis through regulation of the ubiquitin–proteasome system and phosphoinositide 3-kinase/protein kinase B/mTOR pathway. These findings reveal the potential of PUW as a natural therapeutic agent for preventing or managing muscle wasting.
INTRODUCTION
Skeletal muscles are essential in energy metabolism, movement, and overall physical function. 1 Muscles facilitate locomotion by attaching to bones via tendons and are actively involved in regulating metabolic processes such as glucose, lipid, and amino acid metabolism. Muscle atrophy, characterized by a gradual decline in muscle mass and function, markedly affects quality of life by causing muscle weakness, altered fiber composition, and diminished mobility. The mechanisms underlying muscle atrophy are complex; however, its contributing factors include oxidative stress, dysregulation of protein synthesis and degradation, autophagy, and activation of the ubiquitin–proteasome system (UPS), accelerating the breakdown of muscle proteins. 2 Chronic muscle atrophy can exacerbate various health conditions and, if left unaddressed, may lead to sarcopenia, a muscle-wasting condition commonly observed in aging populations. Although resistance training, nutritional supplementation, and pharmacological approaches have been explored as potential strategies to prevent muscle loss, no definitive cure or highly effective treatment is currently available. Therefore, developing novel strategies to prevent and alleviate muscle atrophy is essential for maintaining muscle health and promoting overall well-being.1,2
Myostatin (MSTN), which functions as an inhibitor of muscle growth, is primarily expressed in skeletal muscle and serves an indispensable role in muscle protein metabolism. Elevated MSTN expression has been observed in various muscle disorders, including muscular dystrophy, thereby accelerating protein degradation, inhibiting myocyte differentiation, and suppressing protein synthesis. 3 MSTN activates the Forkhead box O (FoxO)–ubiquitin ligase proteolytic pathway, promoting muscle atrophy through the enhanced expression of E3 ubiquitin ligases, such as F-box only protein 32 (atrogin-1) and muscle-specific RING-finger protein 1 (MuRF1). These ligases degrade essential sarcomeric proteins, including actin, myosin, and troponin, via the UPS, resulting in muscle fiber breakdown. 4 Conversely, muscle growth and regeneration are regulated by the phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt)/mammalian target of rapamycin (mTOR) signaling pathway, which is essential for cellular differentiation, proliferation, and growth. Activation of Akt promotes mTOR expression, enhancing muscle fiber growth and suppressing muscle loss by inhibiting FoxO-mediated catabolic signaling. Downstream targets of mTOR, such as S6K1 and 4E-BP1, further facilitate protein biosynthesis and muscle hypertrophy by regulating translation initiation. In addition, mTOR supports cellular anabolic activities, including the synthesis of proteins, lipids, and nucleic acids, while simultaneously inhibiting catabolic signaling pathways. 5 Therefore, maintaining an equilibrium between protein synthesis and degradation is necessary for muscle homeostasis.
Dexamethasone, a synthetic glucocorticoid extensively used for its potent anti-inflammatory and immunosuppressive effects, has been reported to induce muscle atrophy when administered excessively or over prolonged periods.6,7 Phlomis umbrosa Turcz. (Korean name: Han-Sok-Dan) is a traditional herbal medicine used in Southeast Asia for its antioxidant, anti-inflammatory, and anticancer properties, attributed to its active ingredients, such as phenols and flavonoids.8−11 Although previous studies have explored its effects on bone formation and metabolic disorders, limited research has investigated its potential therapeutic role in muscle atrophy. Considering the increasing global concern over muscle-wasting conditions, including muscular atrophy, the present study determined the impact of Phlomis umbrosa Turcz. water extract (PUW) on dexamethasone-induced muscle atrophy. In particular, we examined its influence on key protein synthesis and degradation pathways, including the PI3K, mTOR, Akt, 4E-BP1, and S6K1 pathways, to assess its role as a natural remedy for muscle preservation.
MATERIALS AND METHODS
Animals and treatments
Eight-week-old male mice were obtained from Samtako Bio Korea (Osan, Gyeonggi-do, Korea) and housed in an animal chamber under controlled environmental conditions (temperature: 23 ± 2°C and humidity: 55 ± 10%) with a 12:12 h light−dark cycle. All animals were fed a 5L79 diet (Orient Bio, Inc., Gyeonggi-do, Korea) and provided water ad libitum. Following a 1-week acclimatization period, the mice were measured for weight and grip strength and subsequently grouped into control (CON), dexamethasone-only (DEX), and dexamethasone plus PUW groups. Dexamethasone (1 mg/kg/body weight) was administered intraperitoneally for 24 days, while PUW (800 mg/kg body weight) was administered orally once daily over a 4-week period. The dosage was selected based on previous in vivo studies using P. umbrosa Turcz. extract, which demonstrated biological activity without adverse effects.12,13 After euthanization, the tissue from the gastrocnemius and quadriceps muscles was separated and subjected to subsequent assays. This study was carried out according to the Institutional Animal Care and Use Committee (IACUC) guidelines and was approved by Chonnam National University (protocol number: CNU IACUC-YB-2022-87).
Exercise
Ladder climbing was used as a resistance exercise in this study. Prior to grouping, the mice underwent a period of familiarization with the exercise. During this pretraining phase, a weight was attached to the base of the tail and was gradually increased from 0% to 100% of the mouse’s body weight. Following grouping, all mice continued the ladder climbing exercise thrice per week for 4 weeks. The ladder, constructed from acrylic, measured 100 cm in height and 7 cm in width, with 1-cm grid steps, and was set at a 70° incline.
Grip strength
Grip strength of the mice was measured twice a week using a BIO-GS3 grip strength meter (Bioseb, Vitrolles, France). Mice were allowed to grasp the grip with their forelimbs and hindlimbs, after which they were pulled down from the grid. This test was performed thrice, and the average peak force divided by each mouse’s body weight was taken for analysis.
Total RNA extraction and reverse transcription–polymerase chain reaction
Total RNA was isolated from homogenized gastrocnemius muscle using easy-BLUE™ reagent (iNtRON Biotechnology, Seongnam, Korea). RNA concentration and quality were assessed using a NanoDrop spectrophotometer (Seolin Tech, Seoul, South Korea), with A260/280 ratios ranging between 1.9 and 2.1. cDNA was synthesized from total RNA using the iScript cDNA Synthesis Kit (Bio-Rad Laboratories, Hercules, CA, USA) according to the manufacturer’s instructions and stored at 4°C for subsequent analysis. Reverse transcription–polymerase chain reaction (RT-PCR) was conducted using SYBR® Green Supermix (Bio-Rad Laboratories). Each reaction mixture contained the cDNA template, SYBR® Green reaction mix, ultrapure water, and gene-specific forward and reverse primers (Table 1). PCR amplification was carried out under the following thermal cycling conditions: an initial denaturation at 95°C for 3 min, followed by 40 cycles of denaturation at 95°C for 10 s, and annealing/extension at 60°C for 30 s. Relative gene expression levels were quantified using the ΔΔCT method.
Primer Sequences

Total protein extraction and western blotting
Gastrocnemius muscles were homogenized in liquid nitrogen using 1× radioimmunoprecipitation assay lysis buffer and the Halt™ protease and phosphatase inhibitor cocktail (Thermo Fisher Scientific, MA, USA). Next, the protein lysates obtained were centrifuged, and the resulting supernatant was collected and used for analysis. Protein concentration was determined using the Bradford assay. These lysates were then boiled at 100°C for 5 min, followed by subsequent separation of the proteins via sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transfer onto a polyvinylidene difluoride membrane. Next, the membrane was incubated with primary antibodies and corresponding secondary antibodies. It was then washed with Tris-buffered saline containing Tween 20, and the protein bands were visualized using enhanced chemiluminescence and photographed using the ChemiDoc MP imaging system (Bio-Rad Laboratories). Subsequently, the value of each protein band was analyzed using Image Lab software (version 6.1; Bio-Rad Laboratories).
Hematoxylin and eosin staining
The harvested quadriceps muscles were immersed in 10% formalin for fixation and subsequently embedded in a paraffin block. Sections were cut from the paraffin block, mounted on glass slides, stained with hematoxylin and eosin (H&E), and observed at 200× magnification using a microscope (Leica, Wetzlar, Germany). Myofiber cross-sectional areas (CSAs) were measured using ImageJ software (NIH, Bethesda, MD, USA).
Statistical analysis
All data were analyzed using SPSS 26.0 software (version 26; SPSS, Inc., Chicago, IL, USA). Statistical significance was determined using one-way analysis of variance, followed by Duncan’s multiple-range test, and was set at P < .05. Results are expressed as the means ± standard errors.
RESULTS
Effect of PUW on body weight and muscle function
To confirm the effect of PUW on muscle atrophy, we evaluated muscle mass and function in an in vivo model of dexamethasone-induced muscle atrophy. Mice were pretreated with PUW and subjected to resistance exercise before the daily administration of dexamethasone for 24 days. The dexamethasone-treated groups (DEX and PUW) displayed a decrement in body weights from 21 to 24 days of dexamethasone treatment and were not affected by the diet (Fig. 1A), suggesting successful induction of muscle atrophy by dexamethasone in this study.
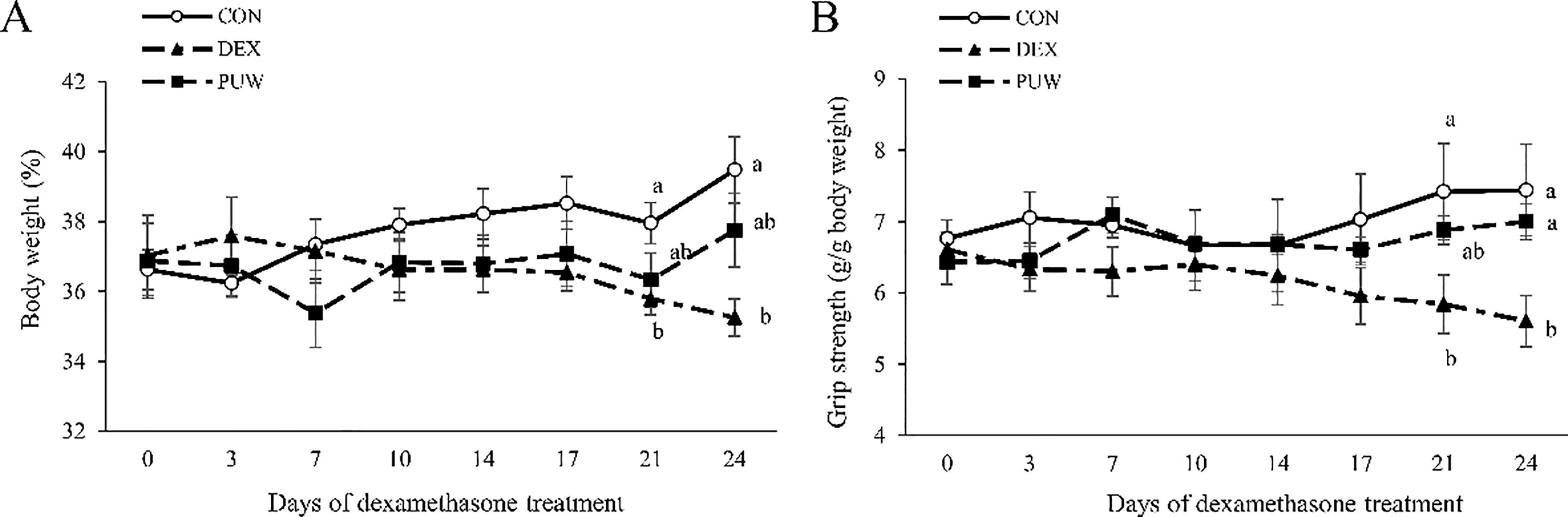
Effect of Phlomis umbrosa Turcz. water extract on body weight and muscle function in dexamethasone-induced muscle atrophy. Mouse
To assess changes in muscle function, grip strength was measured throughout the experiment. The DEX group exhibited a time-dependent decrease in grip strength, which was statistically significant from day 21 of dexamethasone treatment. In contrast, the PUW group initially demonstrated a similar decline but later showed a significant recovery in grip strength compared with the DEX group (Fig. 1B).
Recovery effect of PUW on muscle mass and myofiber damage
After euthanization, we measured the weights of the quadriceps and gastrocnemius muscles. Dexamethasone decreased the weight of both muscles; nevertheless, PUW significantly restored them (Fig. 2A). To further evaluate the protective role of PUW in mitigating muscle atrophy, myofiber CSA was analyzed using H&E staining. Dexamethasone administration markedly reduced myofiber size, whereas PUW treatment significantly restored CSA (Fig. 2B, C).
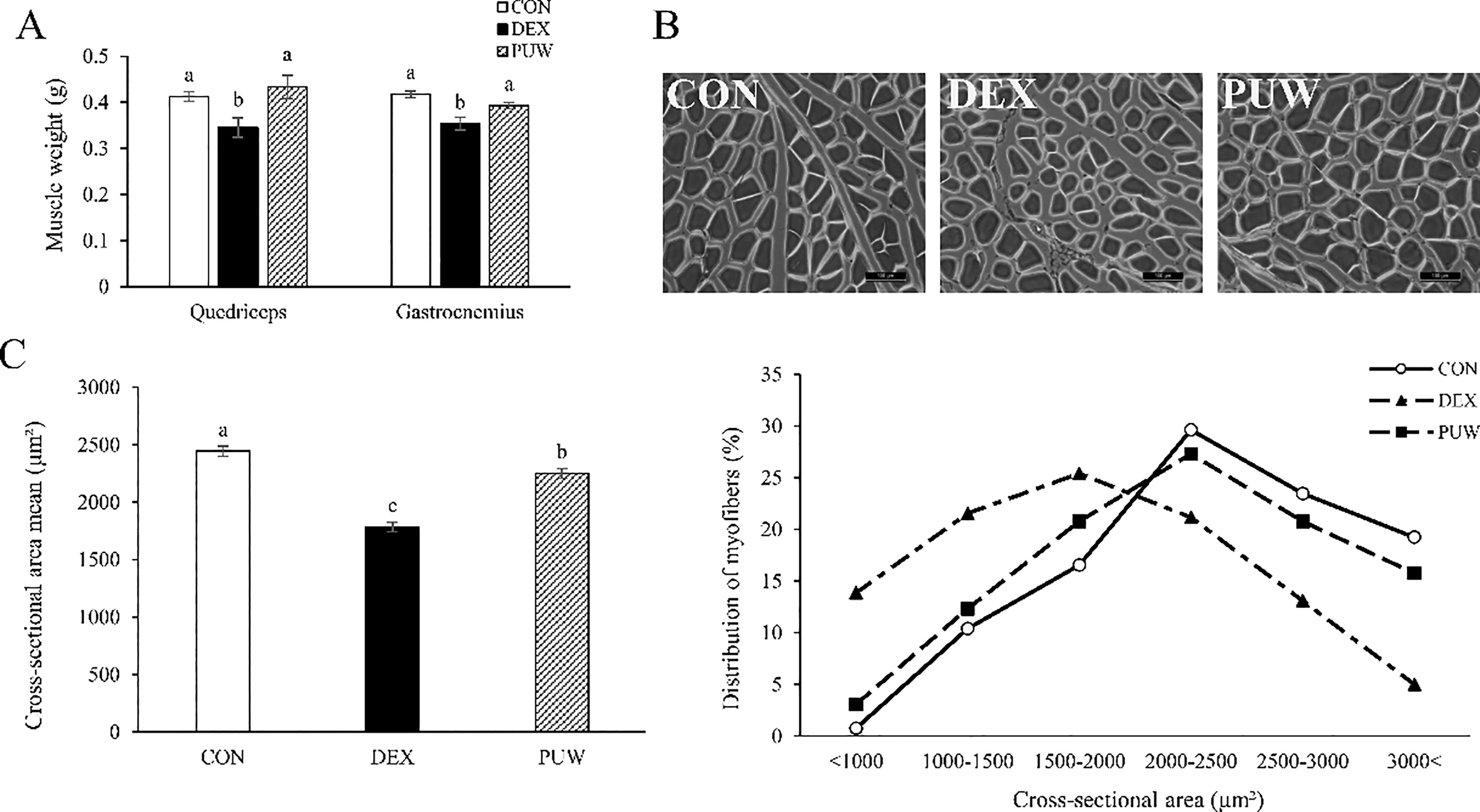
Effect of Phlomis umbrosa Turcz. water extract on muscle mass and myofibers in dexamethasone-induced muscle atrophy.
Muscle fibers were further classified based on size distribution. The DEX group exhibited a higher proportion of small-sized myofibers than the CON group, whereas the PUW group showed a fiber size distribution comparable with the CON group (P < .05, Fig. 2C). These findings confirm that PUW effectively alleviates dexamethasone-induced muscle atrophy by restoring muscle mass and function.
Effect of PUW on muscle protein metabolism-related gene expressions
To evaluate the impact of PUW on muscle protein metabolism at the transcriptional level, mRNA expressions of key genes associated with muscle degradation and synthesis factors were analyzed in the gastrocnemius muscle using RT-PCR. The DEX group exhibited a significant upregulation of MSTN, Atrogin-1, and MuRF1 expression than the CON group. Nonetheless, PUW administration significantly downregulated the expressions of these genes (P < .05, Fig. 3).
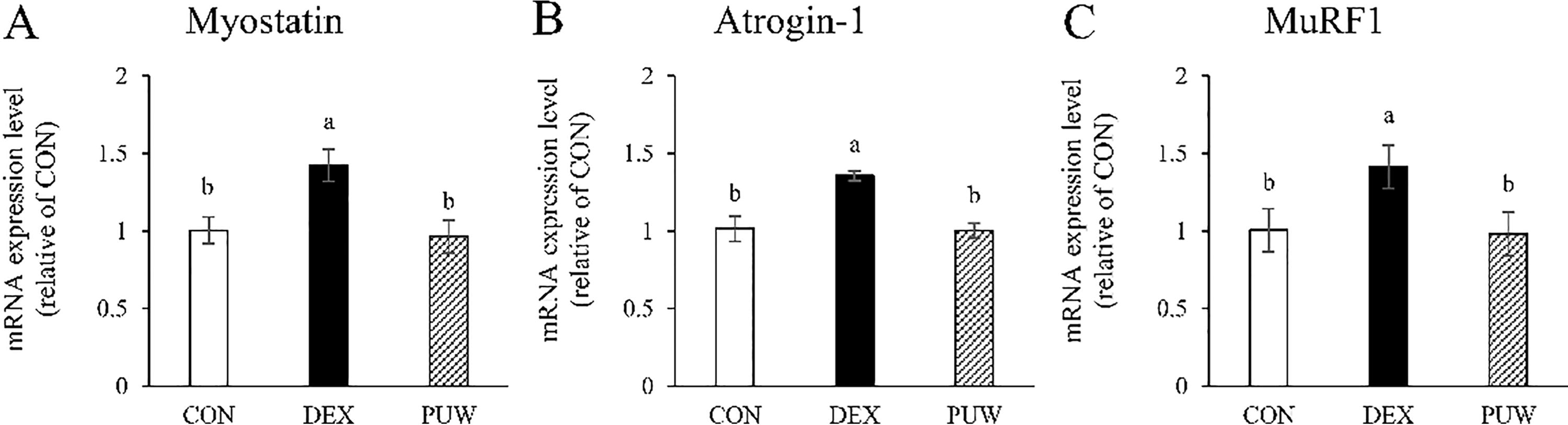
Gene expression of protein degradation factors. The mRNA expression levels of the protein degradation factors
Conversely, the DEX group showed significant downregulation in the mRNA expression of PI3K, Akt, mTOR, and S6K1, key regulators of muscle protein synthesis; however, the levels were restored following PUW treatment (P < .05, Fig. 4). These findings suggest that PUW attenuates dexamethasone-induced muscle atrophy by inhibiting the degradation of muscle proteins and promoting those pathways involved in protein synthesis.
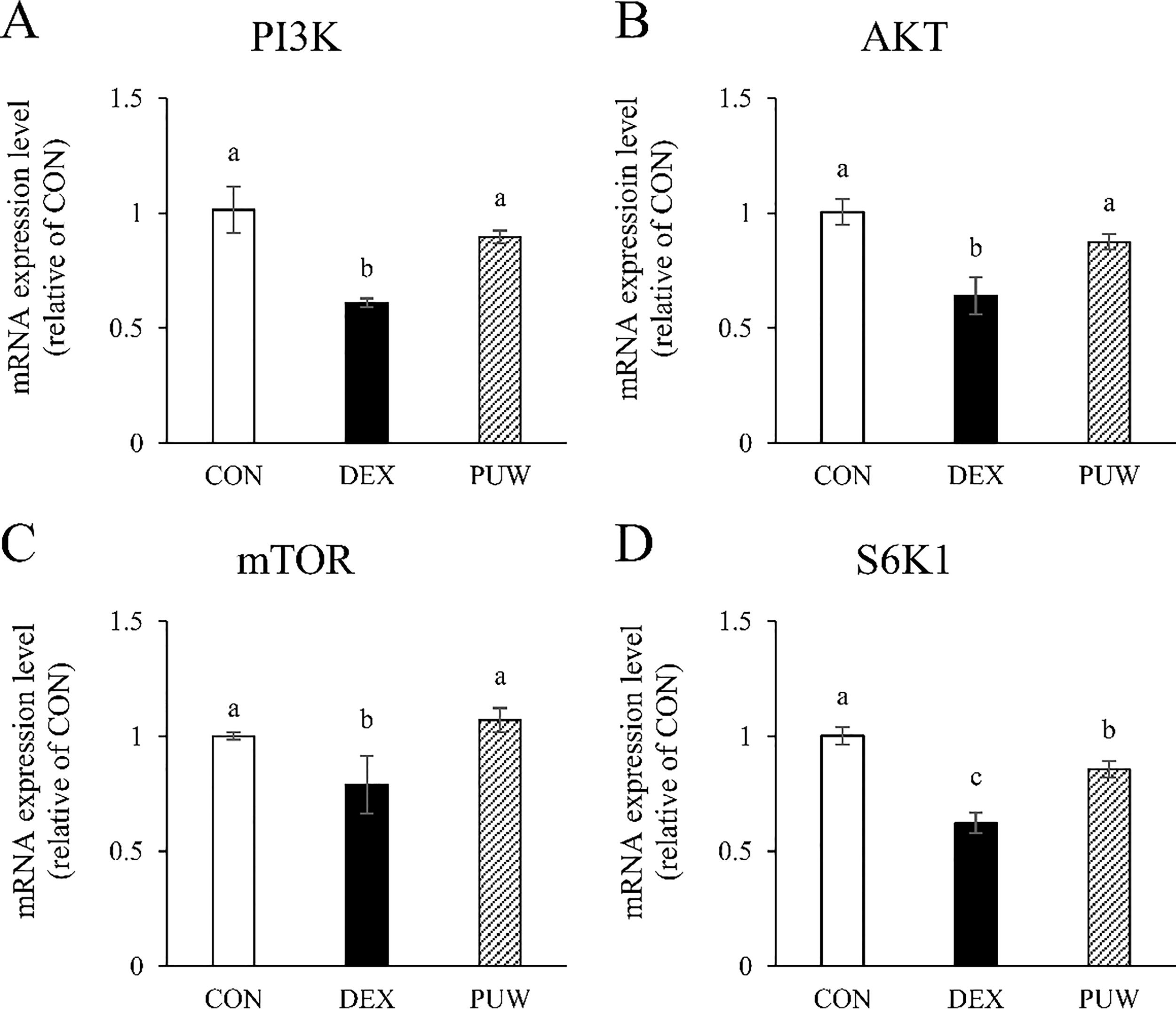
Gene expression of muscle synthesis factors. The mRNA expression levels of the protein synthesis factors
Effect of PUW on muscle protein metabolism-related protein expression
To further examine the influence of PUW on muscle protein metabolism, we performed Western blot analysis on gastrocnemius muscle tissue. The DEX group showed a significant increase in phosphorylated FoxO compared with the CON group, which was in agreement with its role in the UPS (Fig. 5A, B). FoxO, a downstream target of MSTN, is phosphorylated and activated by MSTN, promoting muscle protein degradation. PUW treatment significantly suppressed phosphorylation of FoxO, indicating its potential to inhibit muscle proteolysis.
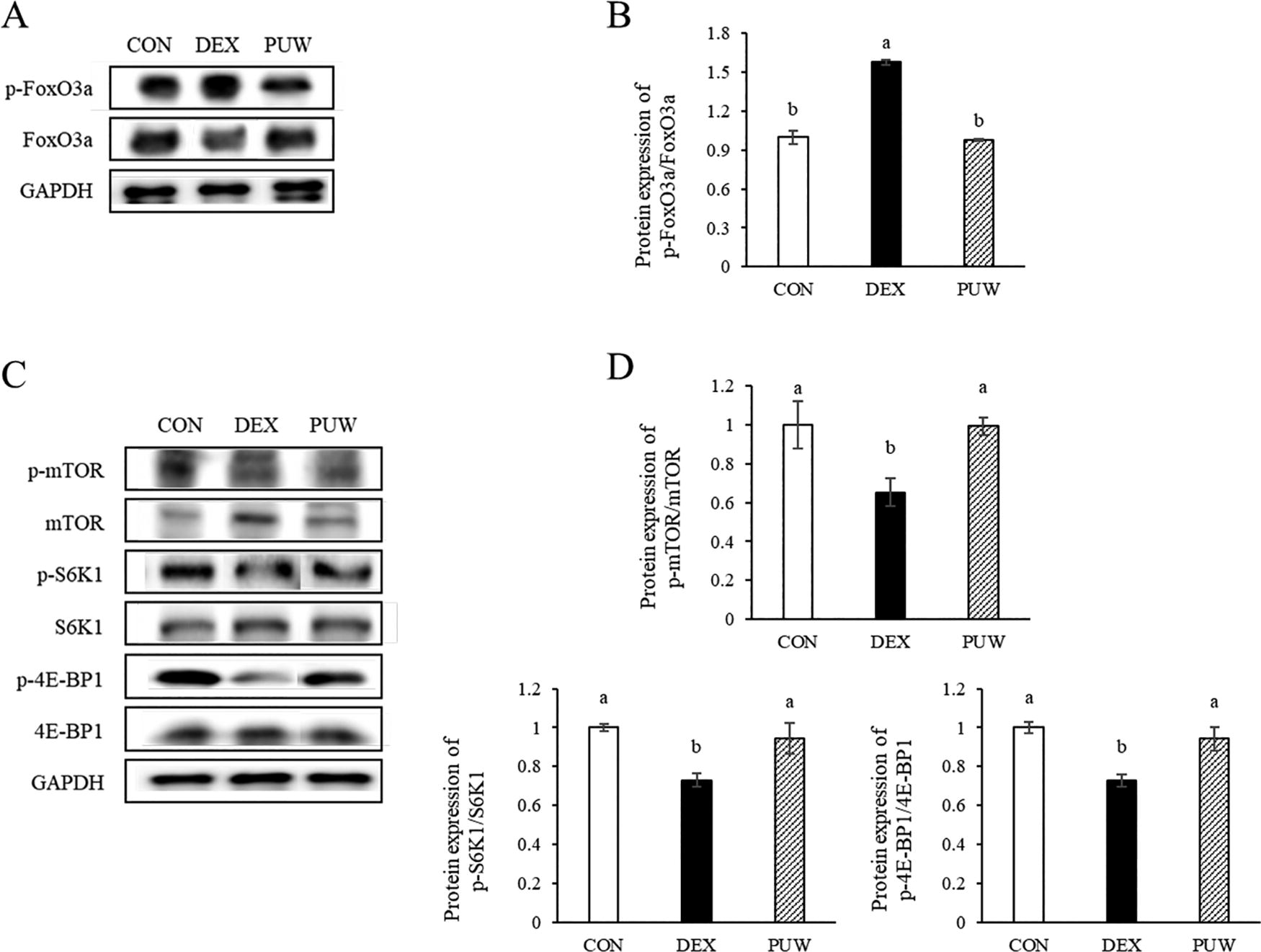
Protein expression levels of muscle degradation and synthesis factors and the effect of Phlomis umbrosa Turcz. water extract.
In addition, phosphorylation levels of mTOR, S6K1, and 4E-BP1, key regulators of muscle protein synthesis, were significantly reduced in the DEX group compared with those in the CON group (P < .05, Fig. 5C, D). PUW administration effectively restored the phosphorylation levels of these essential anabolic marker proteins, suggesting its role in promoting muscle protein synthesis (P < .05). Since mTOR regulates the balance between anabolic and catabolic processes by activating S6K1 and 4E-BP1, these findings indicate that PUW potentially enhances muscle regeneration by restoring protein synthesis pathways.
DISCUSSION
Muscle atrophy is defined as a pathological condition characterized by an imbalance between protein synthesis and degradation, leading to a reduction in muscle mass and impaired muscle function. 14 Glucocorticoids such as dexamethasone exacerbate muscle wasting by promoting catabolic pathways while suppressing anabolic signaling pathways.15,16 In this study, we evaluated the effects of PUW against dexamethasone-induced muscle atrophy in mice. Our findings demonstrate that PUW effectively alleviates muscle loss by restoring muscle protein homeostasis. In particular, PUW suppressed protein degradation pathways and enhanced anabolic signaling, ultimately preserving muscle mass and function.
Dexamethasone, a type of glucocorticoid, is a synthetic adrenocortical hormone with anti-inflammatory effects, which is commonly used in the treatment of various conditions, including allergies, skin diseases, asthma, and chronic obstructive pulmonary disease. It is an effective and safe drug, used in assorted forms and formulations. 17 However, systemic use may elicit side effects such as osteoporosis, muscle pain, high MSTN expression, and edema. In addition, experimental muscle atrophy is frequently induced by dexamethasone treatment due to its ability to trigger myoclonus following prolonged or high-dose administration.18–20 Dexamethasone-induced muscle atrophy is primarily attributed to the activation of UPS, which promotes muscle protein degradation. A critical regulator of this process is MSTN, a key inhibitor of muscle growth. MSTN activates FoxO transcription factors, which, in turn, stimulate the expression of E3 ubiquitin ligases, such as atrogin-1 and MuRF1, leading to increased muscle protein breakdown. Consistent with previous studies, our results reveal that dexamethasone markedly increases MSTN, atrogin-1, and MuRF1 expression at the mRNA and protein levels. However, PUW administration effectively downregulated these catabolic markers, suggesting that PUW mitigated muscle atrophy by inhibiting MSTN-mediated activation of the UPS.
P. umbrosa Turcz. extract has been shown to treat osteoporosis, degenerative arthritis, and bone formation effectively. 21 Several studies have established a strong interrelationship between skeletal muscles and bones. 22 Growth hormones, including insulin-like growth factor 1, are instrumental in the differentiation of osteoblasts and myoblasts, and certain pathways, such as the Wnt signaling pathway, affect both the skeleton and muscles.23−26 In addition, research has shown that bone and muscle mass peak at a similar time in humans, and muscle loss leads to skeletal loss. 27 Furthermore, myokine, a protein secreted by muscles, directly affects the skeleton. 28 Based on these studies, we hypothesized that PUW potentially influences muscle function.
In contrast to activating catabolic pathways, maintaining muscle mass requires the stimulation of anabolic pathways, particularly the PI3K/Akt/mTOR signaling cascade, which is crucial for promoting muscle protein synthesis. PI3K activation leads to Akt phosphorylation, thereby suppressing FoxO-mediated protein degradation while simultaneously stimulating downstream targets such as mTOR, S6K1, and 4E-BP1. As a critical regulator of the translation process and muscle hypertrophy, mTOR activation facilitates protein synthesis while inhibiting catabolic signaling pathways (Fig. 6).4,29,30,31–34 Our study demonstrated that dexamethasone significantly suppressed the expression of PI3K, Akt, mTOR, S6K1, and 4E-BP1, leading to impaired protein synthesis. However, PUW treatment restored the expression of these anabolic markers, indicating that PUW enhanced muscle protein synthesis by reactivating the PI3K/Akt/mTOR signaling pathway.
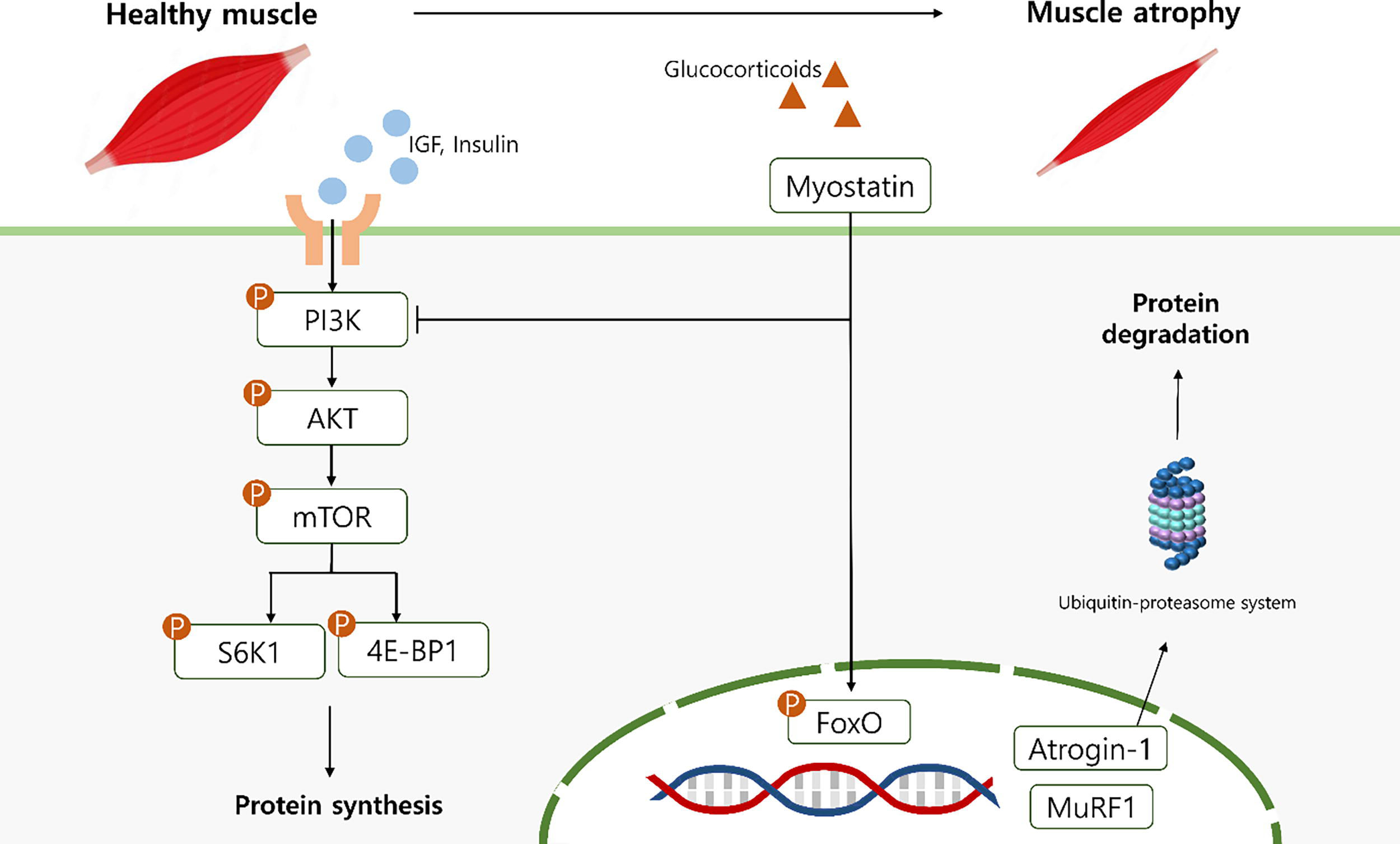
Schematic illustration of the effects of PUW on dexamethasone-induced muscle atrophy. PUW alleviates dexamethasone-induced muscle atrophy by suppressing MSTN/FoxO-mediated protein degradation and restoring PI3K/Akt/mTOR signaling, thereby enhancing protein synthesis and reducing catabolic activity.
Skeletal muscle is comprised of multiple fiber types, primarily type 1 and type 2 fibers, which differ in contraction speed and metabolic characteristics. Type 1 fibers, recognized for their slow contraction and high mitochondrial content, are fatigue-resistant and are mainly engaged during aerobic activities. In contrast, type 2 fibers contract rapidly and fatigue quickly, contributing significantly to early gains in strength during resistance training. Notably, type 2 fibers are preferentially affected in sarcopenia, exhibiting greater susceptibility to atrophy.35,36 To specifically examine the effects of PUW on type 2 fibers, ladder climbing—a form of resistance exercise—was implemented in this study. This model provided a more physiologically relevant approach to assessing muscle responses to catabolic stress and intervention.
In addition, grip strength was assessed as a functional marker of overall muscle strength. As demonstrated in previous studies, grip strength correlated strongly with the strength of other muscle groups and could even predict lower-limb strength and physical performance.37–40 Consistent with these findings, PUW-treated mice maintained grip strength despite dexamethasone-induced muscle atrophy, suggesting systemic protective effects. Moreover, PUW administration suppressed the expression of UPS-related catabolic genes and proteins but restored protein synthesis signaling pathways. These molecular and functional improvements support the potential of PUW to alleviate muscle atrophy by targeting both anabolic and catabolic processes, especially in fast-twitch (type 2) muscle fibers, which are more susceptible to degradation.
Although the present study did not identify the specific active compounds responsible for the effects of PUW, previous studies have reported that the presence of several bioactive components, such as phenolic acids, iridoid glycosides, and terpenoids, in this plant, which are known to exhibit antioxidant and anti-inflammatory properties. Compounds such as luteolin and apigenin, identified in related Phlomis species, presumably contribute to muscle-protective mechanisms by modulating oxidative stress and inflammatory signaling pathways.41 Considering the observed improvements in antioxidant enzyme activity, suppression of signals associated with catabolic pathways, and activation of protein synthesis pathways in our study, such compounds are conceivably involved in the efficacy of PUW. Future studies can focus on the phytochemical profiling and bioactivity-guided fractionation of PUW to identify and isolate its active constituents. This can help clarify the specific molecular targets and underlying mechanisms of the protective effects of PUW on skeletal muscle.
CONCLUSION
In our study, muscle atrophy was induced in mice via dexamethasone injection, resulting in reduced body weight and muscle mass compared with the CON group. Grip strength and myofiber size were also significantly decreased in the DEX group; however, PUW treatment effectively restored these parameters. At the molecular level, the DEX group exhibited upregulation of MSTN, Atrogin-1, and MuRF1 mRNA expression, and downregulation of PI3K, Akt, mTOR, and S6K1. Notably, these alterations were significantly attenuated by PUW administration, indicating its potential therapeutic effect. Furthermore, the increased level of p-FoxO observed in the DEX group was suppressed by PUW. PUW treatment also restored the phosphorylation levels of mTOR, S6K1, and 4E-BP1, initially reduced by dexamethasone. In conclusion, our findings suggest that PUW, a natural compound, may effectively counteract dexamethasone-induced muscle atrophy by regulating key pathways involved in protein degradation and synthesis, highlighting its potential application in the prevention or management of sarcopenia.
AUTHORS’ CONTRIBUTIONS
Y.L.: Investigation, data curation, formal analysis, methodology, visualization, writing—original draft, and writing—review and editing; J.P. and W.J.: Conceptualization, methodology, formal analysis, visualization, writing—review and editing, and supervision. All authors have read and approved the final version of this article.
Footnotes
AUTHOR DISCLOSURE STATEMENT
The authors declare no conflicts of interest.
FUNDING INFORMATION
This work was supported by the National Research Foundation of Korea (NRF) grant, funded by the Korean government (MSIT) (NRF-2021R1C1C2008063).