
Abstract
Purpose:
Recent evidence suggests that oxidative injury plays a significant role in the pathogenesis of retinal degenerative diseases. Para-aminobenzoic acid (PABA) is a cyclic amino acid, which may act to decrease lipid peroxidation and oxidative injury. Our aim was to evaluate the efficacy of PABA in attenuating oxidative injury and rate of retinal degeneration in the rd10 mouse.
Methods:
PABA (50 mg/kg) was administered intraperitoneally six times per week in 28 rd10 mice from postnatal day 3. Twenty-four littermate control mice were similarly injected with saline. At 3, 4.5, and 6 weeks of age, electrophysiological (full field electroretinogram-ERG), quantitative histological, and immunohistochemical techniques were used to assess the course and extent of retinal degeneration. Degree of lipid peroxidation was determined by the measurement of thiobarbituric acid reactive species (TBARS) and retinal carbonyl content was quantified using the 2,4-dinitrophenylhydrazine method.
Results:
Dark adapted mixed rod-cone ERG responses at 3 weeks of age were higher in the PABA-treated group as compared to saline control (P < 0.05). By 4.5 weeks, this protective effect was largely abolished and by 6 weeks ERG was unrecordable in both groups. However, at both 3 and 4.5 weeks of age, light-adapted cone ERG amplitudes were better preserved in PABA-treated animals. At 4.5 weeks, thickness of the outer nuclear layer was 28.6% higher in the peripheral retina of PABA-treated mice as compared to controls (P < 0.05). Quantitative immunohistochemistry revealed 2.4-fold higher red/green cone opsin content in the retinas of PABA-treated mice (P < 0.005). At both 3 and 4.5 weeks, levels of TBARS and protein carbonyls were 49%–69% lower in PABA-treated retinas (P < 0.05–0.0005), suggesting less oxidative injury.
Conclusions:
PABA treatment may protect retinal function and attenuate the course of retinal degeneration in rd10 mice. Biochemical parameters indicate a lower degree of oxidative injury in PABA-treated retinas. PABA may potentially serve as an addition to antioxidative treatment for retinal and macular degenerations.
Introduction
R
In retinitis pigmentosa, progressive loss of photoreceptors occurs. In many cases, the mutated gene is rod-specific, leading to rod photoreceptor apoptosis and death. Cone degeneration is often secondary to rod loss. There is evidence that in many forms of retinal degeneration, photoreceptors eventually die via apoptosis,4 but the mechanisms that lead to cell death in each type of degeneration are not completely understood.5
Reactive oxygen species (ROS) were reported as possible mediators of in vitro photoreceptor apoptosis.6 , 7 Indeed, the retina is one of the most metabolically active tissues in the body, consuming oxygen more rapidly than other organs including the brain.8 Recently published data suggest that oxidative stress may play a critical role in the pathogenesis of neurodegenerative disorders9–12 including retinal degenerative diseases.13–18
Experiments in vivo also confirm that oxidative stress represents an important factor in pathogenesis of RP that leads to rod and cone death.19–21 In a pig model of RP, Shen and colleagues19 showed significant lipid peroxidation in cone photoreceptors. Recently, Ahuja-Jensen and colleagues22 demonstrated decreased levels of glutathione peroxidase and elevated malondialdehyde in the inner retina of rd1 mice with retinal degeneration caused by a mutation in the phospodiesterase (PDE) gene, suggesting the involvement of oxidative injury and lipid peroxidation reactions in the degenerative process. We have also demonstrated changes in iron homeostasis and oxidative injury in rd10 mouse retinas during the course of degeneration.23
It has recently been shown that treatment with a combination of antioxidants may decrease photoreceptor death in a number of mouse models of RP.24–26 In humans, Age-Related Eye Disease Study (the AREDS) trial suggested that antioxidants and zinc, alone and in combination, can slow progression of age-related macular degeneration.27 , 28 These findings support the importance of ROS-mediated injury in the development and progression of retinal degenerations, and demonstrate the therapeutic potential of antioxidants in treatment of RP and other retinal and macular dystrophies.
Para-aminobenzoic acid (PABA) is a nonprotein amino acid that is widely distributed in nature. Chemically, PABA contains a benzene ring substituted with an amino group and a carboxylic acid. It is sometimes referred to as vitamin Bx, but it is neither a vitamin nor an essential nutrient for humans. In the past, PABA has been widely used as an ultraviolet filter in sunscreen formulations. Nowadays, PABA derivates are continuing to attract attention of investigators due to their wide spectrum of biological effects and low toxicity. PABA induces endogenous interferon production and manifests antiviral properties,29 , 30 reduces the mutagenicity of various chemical mutagens,31 , 32 shows a radioprotective effect,33 reduces cis-diamminedichloroplatinum(II) nephrotoxicity,34 , 35 has anticoagulant activity,36 and, in combination treatment with antibiotics, has synergistic inhibitory activity against Pseudomonas aeruginosa and Staphylococcus aureus.37
It has been shown by Akberova and colleagues38 that PABA decreases the levels of hydroperoxide and malonic dialdehyde, and normalizes catalase activity in the cornea and lens of the eye following hypoxia. In a second study, these investigators demonstrated that PABA attenuates lipid peroxidation in guinea pig retinas exposed to hypoxia,39 suggesting that it has antioxidant properties. Stroeva and colleagues showed that PABA reaches the retina following subcutaneous injection in the rat, and stimulates membrane disk formation in photoreceptor outer segments of Campbell rats with retinal degeneration.40 , 41
The purpose of the present study was to evaluate the efficacy of PABA in reducing the rate of retinal degeneration in the rd10 mouse model of RP.
Methods
Animals and anesthesia
rd10 mice (Jackson Labs, Maine, USA), manifest a rapidly progressive retinal degeneration within the first few months of life caused by a mutation in the rod phosphodiesterase gene.42 , 43 From 2 to 3 weeks of age and on, histological examination reveals progressive loss of photoreceptors in the outer nuclear layer with rapidly deteriorating electroretinographic (ERG) responses, and the degenerative process is largely complete by 6–8 weeks of age.42 , 44
Animals were maintained on a 12:12 h light:dark cycle without limitation of food and water. All procedures were conducted in compliance with the ARVO Statement for the Use of Animals in Ophthalmology and Vision Research and were approved by the Animal Ethics Committee of the Hebrew University-Hadassah Medical School. For ERG recordings, animals were anesthetized with intramuscular injections of ketamine HCl (50 mg/kg) in combination with the relaxing agent xylazine (5.0 mg/kg). Local anesthetic drops were also administered.
Experimental groups and protocol
rd10 mice included in the treatment group (n = 28) received intraperitoneal injections of PABA in saline (50 mg/kg; Sigma-Aldrich Inc., St. Louis, MO, USA) six times per week from postnatal day 3 (P3) until 6 weeks of age. Twenty-four littermate mice served as controls and received intraperitoneal injections of similar volumes of sterile saline solution according to the same protocol.
Electroretinography (ERG)
At 3, 4.5, and 6 weeks of age, full field ERGs were recorded in anesthetized animals following overnight dark adaptation using a Ganzfeld dome and a computerized system (Espion, Diagnosys Llc, Littleton, MA). Pupils were dilated with 1% tropicamide and 2.5% phenylephrine (Fisher Pharmaceuticals, Tel-Aviv, Israel). Local anesthetic drops (benoxinate HCl, 0.4%; Fisher Pharmaceuticals) were administered prior to insertion of gold-wire active electrodes placed on the central cornea. A reference electrode was placed on the tongue and a needle ground electrode was placed intramuscularly in the hip area. Dark-adapted rod and mixed cone-rod responses to a series of white flashes of increasing intensities (0.05–48cd·s/m2) and light-adapted 1 Hz and 16 Hz cone responses were recorded. All ERG responses were filtered at 0.3–500 Hz, and signal averaging was applied.
Retinal histology
At 3 and 4.5 weeks of age, eyes were enucleated and fixed in Davidson solution. After dehydration, samples were embedded in paraplast and serially cut into 4 µm sections through the center of the optic nerve. For descriptive histology and quantitative analysis, sections were stained with hematoxylin and eosin or with the proper fluorescent markers as detailed below. All observations and photography were performed using a fluorescent microscope (Olympus BX41, Japan) equipped with a DP70 digital camera. Image processing and quantification were performed using Adobe Photoshop 7.0 and ImageJ software (http://rsb.info.nih.gov/ij/download.html).
Structural analysis of outer nuclear layer
In order to standardize measurements of photoreceptor degeneration between animals, in all cases histologic sections that include the optic nerve were taken. This allows to control for two possible biases, related to differential rate of degeneration across the retina of rd10 mice43 and cone opsin distribution in the mouse retina.45 Since the rate of degeneration differs between central and peripheral retina, each hemi retina was divided into three parts of equal length: central, mid-peripheral and peripheral retina. The thickness of the outer nuclear layer and the number of photoreceptor nuclei rows were measured three times in the center of each part, and averaged with the measurements in the corresponding retinal third in the opposite hemiretina of the same section.
Immunohistochemistry
Deparaffinized sections were incubated in a decloaking chamber (Biocare Medical, USA) with 10 mM citrate buffer (pH 6.0) at 110°C, blocked with PBS solution containing 1% bovine serum albumin, 0.1% triton-°100, and 3% normal donkey serum, and subsequently incubated for 1 h with one of the following primary antibodies: antir-hodopsin (rabbit polyclonal, 1:100; Santa Cruz Biotechnology, Inc.), anti-blue-sensitive opsin (goat polyclonal, 1:75; Santa Cruz Biotechnology, Inc., Santa Cruz, CA), or anti-red/green opsin (rabbit polyclonal, 1:100; Chemicon International, Inc., Billerica, MA). After washing, the appropriate secondary anti-body was applied for 1 h: CY™2-conjugated donkey anti-rabbit IgG (1:200), or rhodamin Red™-X-conjugated donkey anti-goat IgG (1:200; both from Jackson ImmunoResearch Laboratories, Inc., West Grove, PA). Nuclei were counterstained with 4,6-diamidino-2-phenylindole (DAPI)–containing mounting medium (Vector Laboratories, Burlingame, CA). To determine the specificity of the antigen–antibody reaction, corresponding negative controls with the secondary antibody alone were performed.
Quantitative immunohistochemistry
All measurements were performed on sections passing through the optic nerve with similar cone gradient. Content of rhodopsin, red/green and blue opsins was quantified by measuring the fluorescence-positive area in digital images. Only areas in which fluorescence exceeded a certain threshold were defined as “positive” and included in the measurement. In each eye at least 4–5 sections were measured, results were averaged and means were used for further statistical analysis.
Measurements of total protein, protein carbonylation, and TBARS levels
At 3 and 4.5 weeks of age eyes were enucleated, the retinas were gently dissected out, and placed in lysis buffer (1% deionized Triton X-100 and 0.1% sodium azide in 50 mM Tris-HCl pH 7.5 with 0.25 µM of phenylmethylsulfonyl fluoride). For further biochemical analysis retinas were homogenized in a motor-driven polytetrafluoroethylene-glass homogenizer and sonicated for 30 s. Samples were then incubated on ice for 30 min (vortexed every 5–6min) and centrifuged at 3,000 rpm (1,000g) for 15 min. The supernatant was collected and used for analysis.
Total protein content was estimated using the bicinchoninic acid (BCA) assay Kit (Rockford, IL, USA) in 96-well plates (TC Microwell 96F SI W/LID Nunclon D, Roskilde, Denmark). The results were read using a micro plate reader (MR 5000 Dynatech Laboratories, Chantilly, VA, USA) equipped with a 570 nm filter.
Degree of lipid peroxidation was estimated by measurement of thiobarbituric acid reactive species (TBARS) in six PABA-treated and four saline-treated pools of retinal homogenates (each containing 2–3 retinas).46 Retinal carbonyl content was estimated using the 2,4-Dinitrophenylhydrazine method as described by Levine and colleagues.47
Statistical analysis
Statistical analysis was performed using the unpaired two-tailed t-test to compare ERG responses and histologic data between the groups, and the ANOVA test was applied for examining the biochemical data. Differences were considered statistically significant when P ≤ 0.05. Results are presented as mean ± SEM.
Results
In rd10 mice treated by intraperitoneal injections of PABA, retinal functions, and particularly cone-driven responses, were better preserved compared to saline-injected littermates (Fig. 1). Dark-adapted b-wave amplitudes at 3 weeks were 1.3-fold higher in the PABA-treated group as compared to saline controls (P < 0.05; Fig. 1C). By 4.5 weeks this protective effect of PABA on dark-adapted retinal function was largely abolished (Fig. 1D). Dark-adapted a-wave amplitudes did not differ between the two groups at 3 weeks of age (Fig. 1A) and were very difficult to quantify at 4.5 weeks of age because of their very low amplitudes under most stimulating intensities (Fig. 1B). Differences between the two groups in cone-derived function under light-adapted conditions were more consistent. Both at 3 and 4.5 weeks, ERG responses showed higher cone-derived amplitudes in PABA-treated animals, attaining statistical significance in some cases and showing a trend for higher amplitudes in others (Fig. 1E–1F). At 6 weeks of age ERG responses were nondetectable in most animals of both groups.

Retinal function as measured by full field electroetinography (ERG) in para-aminobenzoic acid (PABA)- and saline-treated rd10 mice at the age of 3 weeks (left) and 4.5 weeks (right). (
The effect of PABA on the course of retinal degeneration was also assessed using morphometric histological parameters (Fig. 2). The thickness of the outer nuclear layer (ONL), which contains the nuclei of the photoreceptors, and the number of nuclei rows in the ONL were determined. As in rd10 mice the extent of degeneration is not uniform in different regions of the retina,44 all measurements were performed in a consistent and controlled fashion in different regions of retinal sections passing through the optic nerve (detailed in Methods). Already at 3 weeks of age thickness the of ONL and number of rows of nuclei were markedly reduced in both experimental groups as compared to normal C57Bl6 mice (which normally have ∼12 rows of nuclei in the ONL at this age). The ONL continued to degenerate during the next 1.5 weeks (Fig. 2B and 2D as compared to Fig. 2A and 2C; note 2-fold difference in y-axis). At 3 weeks, there is a trend for a thicker ONL and more rows of nuclei in the central and mid-peripheral retina in PABA-treated mice as compared to saline-treated controls (Fig. 2A and 2C). At 4.5 weeks, both ONL thickness and number of nuclei rows in the ONL were significantly larger in the peripheral retina of PABA-treated mice in comparison with saline-treated controls (P < 0.05, Fig. 2B, 2D, and 2H compared to 2F).

Outer nuclear layer (ONL) thickness and number of photoreceptor nuclei rows in para-aminobenzoic acid (PABA)- versus saline-treated rd10 mice retinas. (
To further characterize the effect of PABA on photoreceptor survival, rhodopsin, blue opsin and red/green opsin contents were quantified in retinal sections using immunofluorescent techniques (Fig. 3). Rhodopsin content and blue opsin content did not differ between the experimental groups (Fig. 3A and 3B, left and central columns; note the large decline in rhodopsin content in rod photoreceptors between 3 and 4.5 weeks of age, reflecting the severe and rapid nature of the degenerative process in this model). At both 3 and 4.5 weeks of age, red/green cone opsin content was significantly higher in the retinas of rd10 mice treated with PABA as compared to saline controls (P < 0.05, Fig. 3A and 3B, right columns; Fig. 3C and 3D show cells containing red/green opsin at 4.5 weeks of age in a saline-treated (C) vs. PABA-treated (D) eye).
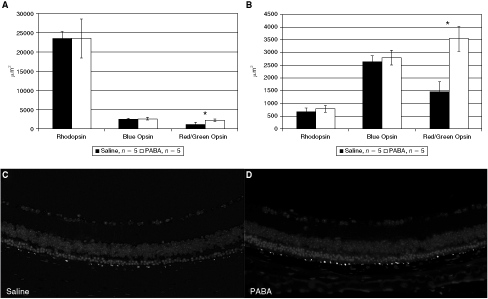
Retinal rhodopsin, blue opsin and red/green opsin content in para-aminobenzoic acid (PABA)- versus saline-treated rd10 mice. (
To examine whether treatment with PABA affected extent of oxidative injury in the retina, we measured degree of lipid peroxidation [as determined by the thiobarbituric acid reactive species (TBARS) production assay] and the degree of oxidative injury to proteins as reflected by levels of protein carbonyls (Fig. 4). Both measures show a highly significant protective effect of PABA treatment. TBARS production was lower by 49% at 3 weeks (P < 0.05) and by 69% at 4.5 weeks (P < 0.0005) in PABA-treated animals versus saline-treated controls. Protein carbonyl levels were similarly reduced by 68% at 3 weeks (P < 0.005) and 58% at 4.5 weeks (P < 0.05). These results suggest reduced oxidative injury in PABA-treated retinas.
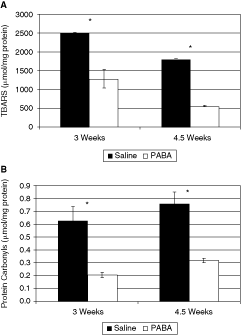
Measures of oxidative injury in para-aminobenzoic acid (PABA)- versus saline-treated rd10 mice retinas. (A) Thiobarbituric acid reactive species (TBARS) production, a measure of lipid peroxidation. (B) Protein carbonyls levels, an indicator of protein oxidative injury. Black bars, saline-treated control; white bars, PABA-treated group. All results are presented as mean ± SEM; six PABA-treated and four saline-treated pools of retinal homogenates (each containing 2–3 retinas) were included. ∗Statistically significant difference.
To rule out systemic toxicity of PABA treatment, animal weight and survival were monitored. No differences were observed between PABA-treated and saline-treated groups.
Discussion
In the present study, functional and structural parameters suggest that systemic administration of the amino acid PABA may confer partial protection against the rapid and aggressive retinal degenerative process in the rd10 mouse model of RP. In this model, a mutation in the β-subunit of the rod phosphodiesterase gene leads to severe retinal dysfunction and progressive, massive death of rod but also cone photoreceptors within the first 2 months of life. In PABA-treated animals, the protective effect on dark-adapted, mainly rod-derived retinal function was more apparent at 3 weeks and was largely abolished by 4.5 weeks, while structural differences were more apparent at 4.5 weeks. This is probably due to a short delay between progression of electrophysiological dysfunction and the appearance of the corresponding structural changes. The protective effect of PABA on cones, the “innocent bystanders” in this rod-driven degenerative process, was more consistent. Light-adapted cone function and red/green cone opsin content were better preserved in PABA-treated eyes than in saline-treated control littermates at both 3 and 4.5 weeks of age. Measures of oxidative injury to retinal lipids and proteins at these two time-points showed 50%–70% protection conferred by systemic PABA administration. The functional and structural rescue effect of PABA was abolished by 6 weeks of age, when in both groups ERG responses became unrecordable and only sparse photoreceptor nuclei were evident in histological sections.
The ability of PABA to reduce retinal lipid peroxidation was also described by Akberova and colleagues in a model of hypoxia.29 , 39 Interestingly, a trophic effect of PABA on photoreceptor outer segment disks in another model of hereditary retinal degeneration, the Campbell rat, was previously suggested by Stroeva and colleagus.40 PABA has an excellent safety profile. There have been rare reports of liver toxicity following ingestion of large doses,48 and possible interactions with sulfa drugs,49 but it is considered a dietary supplement and intake is not restricted. In the current study animal weight, survival and behavior did not differ between control and PABA-treated groups.
In genetically-determined retinal degenerations, besides death of the retinal cells which harbor the mutated gene, very often collateral injury to other photoreceptor populations, which do not express the defective protein, occurs. As such, both in humans and in animals with retinal degeneration caused by rod-specific mutations, cone photoreceptor death often ensues. Different mechanisms which may explain this collateral injury have been proposed, including the possibility that rods normally secrete factors necessary for cone survival,50 that dying rods secrete toxic agents, which affect the cones,51 and also the possibility that oxidative injury plays a role. Indeed, treatment with antioxidants has been shown to confer protection in AMD in humans (AREDS trial)27 , 28 and in animal models of retinal degeneration.25 , 26 In one study, systemic treatment of rd124 , 25 mice with lutein, zeaxanthin, alpha lipoic acid, and reduced l-glutathione decreased oxidative DNA damage in rod photoreceptors and increased photoreceptor survival. Importantly, in that study, administration of individual antioxidants had no significant rescue effect but the combination was highly effective, indicating an additive or synergistic effect. In rd1, rd10, and Q344ter mice, a combination of the antioxidants alpha-tocopherol, ascorbic acid, Mn(III)tetrakis (4-benzoic acid) porphyrin, and alpha-lipoic acid also enhanced cone survival.26 The effect of each agent alone was relatively small and often not statistically significant, but given together, substantial rescue was achieved. These studies highlight two major points: first, oxidative injury probably plays an important role in the pathogenesis of retinal degenerations caused by rod-specific mutations, and decreasing this injury promotes cone survival. Second, combining antioxidants provides an additive or perhaps even synergistic effect. In this context, PABA, with its antioxidative activity and excellent safety profile, may be a beneficial addition.
Footnotes
Acknowledgment
This study was supported by The Yedidut 1 Research Fund.