
Abstract
Abstract
Purpose:
Oxidative damages to the retinal pigment epithelium (RPE) have been suggested to play a key role in the pathogenesis of age-related macular degeneration. trans-Resveratrol (3,4′,5-trihydroxystilbene) is a nonflavonoid dietary polyphenol with various pharmacological effects, including antioxidant activity. The purpose of this study was to evaluate the potential protective effect of resveratrol against hydrogen peroxide induced oxidative stress in cultured human RPE cells.
Methods:
Human retinal D407 RPE cells were pretreated with resveratrol at 3 different concentrations (25, 50, and 100 μM) for 24 h and exposed for 1 h to 500 μM hydrogen peroxide. Cell viability, cytotoxicity, and the level of intracellular reactive oxygen species (ROS) were determined in basal and oxidative stress conditions. The concentration of reduced glutathione and the activities of catalase, superoxide dismutase, and glutathione peroxidase were also examined under both experimental conditions.
Results:
Resveratrol in culture media had no cytotoxic effect at a concentration of 25–100 μM but showed a protective effect against hydrogen peroxide-induced cytoxicity. Pretreatment with resveratrol induced a significant, dose-dependent increase of superoxide dismutase, glutathione peroxidase, and catalase activities. Moreover, resveratrol significantly enhanced the level of reduced glutathione under both basal and oxidative stress conditions. The significant inhibition of the intracellular ROS generation supports the hypothesis that resveratrol can also contribute to the antioxidant defense by directly scavenging the ROS in RPE cells.
Conclusions:
Our results indicate that treatment of RPE cells with resveratrol at micromolar concentrations confers a marked protection against oxidative stress. These data suggest that dietary supplementation of resveratrol may contribute to the prevention of RPE degeneration induced by oxidative stress.
Introduction
trans-Resveratrol (3,4′,5-trihydroxystilbene) is a polyphenol phytoalexin that can be found in more than 70 plants, including grapes, peanuts, and berries (mulberry). 4 Despite some concerns about the rate of intestinal uptake and on its bioavailability, an ever-growing body of in vivo and in vitro evidence points to protective effects of resveratrol in various diseases. Resveratrol exerts a wide range of biological effects, including cardiovascular protection, neuroprotection, anti-inflammatory, cancer chemoprevention, and antioxidant properties.4–6 In vitro studies have shown that resveratrol improves the antioxidant status in different types of cultured cells by increasing the antioxidant enzyme activities.7–9
Few studies deal with resveratrol effects on the prevention of eye diseases or on the protection of eye cells. However, moderate wine consumption has been associated with a decreased risk of development of AMD. 10 Resveratrol reduces hyperproliferation by inhibiting mitogen-activated protein kinase/extracellular signal-regulated kinase (ERK) and the ERK 1/2, and it reduces oxidative stress in cultured human RPE cells. 11 Moreover, resveratrol prevents ocular inflammation in endotoxin-induced uveitis by inhibiting oxidative damage and the nuclear factor-kappa B. Resveratrol also reduces oxidative damage on phagocytic functions, ameliorates UVA-mediated inhibition of human RPE cell phagocytosis, and prevents antibody-induced apoptotic death by the upregulation of Sirt1 and Ku70 activities and inhibition of caspase-3 activity in EIA.NR3 retinal cells.12–15
Most of the beneficial effects of resveratrol are related to its antioxidant potential. The goal of this study was to evaluate the effect of resveratrol on the antioxidant status of RPE cells in vitro by determining the activity of antioxidant enzymes, of the glutathione concentration, and of the intracellular reactive oxygen species (ROS) generation, in basal and induced oxidative stress conditions. Currently, there is no efficient treatment for AMD and dietary strategies that enhance the uptake of the antioxidants might be important for the prevention of this disease.
Methods
Chemicals
GIBCO® Dulbecco's modified Eagle's medium (DMEM) was purchased from Invitrogen. Fetal bovine serum, penicillin, streptomycin, amphotericin B, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), 2′,7′-dichlorodihydrofluorescein diacetate (DCF-DA), and resveratrol were purchased from Sigma. Sodium pyruvate, hydrogen peroxide, and all the other chemicals used were of analytical grade and supplied from Merck.
Cell culture and treatment
The human retinal pigment epithelial cell line D407 was obtained from Dr. Richard Hunt (University of South Carolina). The cells were maintained in DMEM supplemented with 10% fetal bovine serum, 1 mM sodium pyruvate, 100 U/mL penicillin, 100 μg/mL streptomycin, and 2.5 μg/mL amphotericin B, at 37°C, 5% CO2, and 95% relative humidity. The cells were seeded in 25 cm3 flasks at a concentration of 6×105, or in 96-well plates (104 cells/well). After reaching 80% confluence, the growth medium was removed and replaced by a medium containing resveratrol. Resveratrol was dissolved in ethanol and diluted to the appropriate concentration (25, 50, and 100 μM) in the cell culture medium. The concentration of ethanol did not exceed 0.01% concentration in the culture medium. Control cells received the same concentration of ethanol in the culture medium. To ensure reproducibility, assays were carried out between passage no. 25 and passage no. 50.
Exposure of cells to H2O2
Twenty-four hours after resveratrol treatment, the culture medium was removed, and after being washed with PBS, the cells were exposed to 500 μM H2O2 for 1 h in phenol-red-free DMEM. Thereafter, the cells were washed twice with cold PBS, and then specifically lysed for each enzyme determination.
Viability assay
The MTT assay uses the property of viable cells to reduce 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT reagent) into a blue formazan. The method reflects the activity of mitochondrial succinate-dehydrogenase. 16 D407 cells were plated (10,000 cells per well) in 96-well plates and treated with resveratrol and/or H2O2 as described above. The cells were washed with PBS and 200 μL solution of MTT 0.5 mg/mL in HBSS buffer was added to each well. After 2 h of incubation, the MTT reagent was removed and the formazan particles were solubilized with 200 μL DMSO. The absorbance was read at 550 nm and at 630 nm (for background) with a microplate plate reader HT BioTek Synergy (BioTek Instruments). Cell viability was expressed as a percentage of control (cells incubated in normal medium only).
Evaluation of cell damage by LDH leakage assay
D407 cells were seeded (10,000 cells/well) in 96-well plates for 24 h, and treated with resveratrol and/or H2O2 as described above. The amount of LDH released into the cell culture medium was evaluated using the colorimetric LDH assay kit (Sigma). The method is based on the ability of LDH to convert NAD+ to the reduced form, which in turn converts tetrazolium dye to a colored formazan.
Measurement of intracellular ROS
The method uses the fluorescent probe DCF-DA, which is cell membrane permeable. DCF-DA is hydrolyzed by cellular esterases to DCF, which then reacts with intracellular ROS to form the fluorescent DCF, which is measured with a fluorescence plate reader. Cells were cultured (10,000 cells per well) in 96-well black plates and incubated with DCF-DA 20 μM in HBSS. Fluorescence was monitored for 4 h at 37°C at excitation wavelength 485/10 nm and emission wavelength 528/20 nm. 17
Antioxidant enzymes and reduced glutathione
After completing the treatment, cell extracts were prepared according to the manufacturer's instruction. For each parameter the protein content was determined by the bicinchoninic acid assay (Sigma). For all antioxidant enzymes, the activity is expressed in mg protein.
Superoxide dismutase assay
Intracellular superoxide dismutase (SOD) activity was assayed with a commercially available kit (Cayman Chemical Company). The method is based on the conversion of a tetrazolium salt into a formazan by the superoxide radical generated in the xanthine/xanthinoxidase system. A standard curve obtained with bovine erythrocyte SOD was used to determine the enzyme activity. The absorbance was monitored at 460 nm using a microplate reader.
Glutathione peroxidase assay
Glutathione peroxidase (GPx) activity was determined using a commercial kit (Cayman Chemical Company). The GPx Assay kit measures GPx activity indirectly by a coupled reaction with glutathione reductase (GR). Oxidized glutathione (GSSG) produced upon reduction of hydroperoxide by GPx is recycled to its reduced state by GR and NADPH. The absorption decrease rate at 340 nm is directly proportional to the GPx activity.
Catalase assay
Sample preparation and assay procedure followed the kit instructions (Cayman Chemical). The method is based on the reaction of catalase with methanol in the presence of an optimal concentration of H2O2. The formaldehyde produced is measured colorimetrically with 4-amino-3-hydrazino-5-mercapto-1,2,4-triazole (Purpald) as chromogen. A standard curve made with bovine liver catalase was used to determine the enzyme activity. The absorbance was measured at 540 nm with a microplate reader.
Glutathione assay
The reduced glutathione (GSH) assay was performed using an optimized enzymatic recycling method with GR (Cayman Chemical Company). The method is based on the reaction of GSH with 5,5′-dithio-bis-2-(nitrobenzoic acid) (DTNB) in the presence of GR, glucose-6-phosphate dehydrogenase, and NADP+ and produces a yellow-colored 5-thio-2-nitrobenzoic acid (TNB). The standard curve was made with a GSSG standard, having the equivalent GSH concentration between 0 and 16 μM. Results are expressed as μmol GSH/mg protein in cell pellets. The absorbance was read after 30 min at 405 nm, using a microplate reader.
Statistical analysis
This analysis was carried out using the Dunnett multiple comparison test of Graph Pad Prism version 5.00. Data represent the means±SD of at least 3 independent experiments (*significant P<0.05, **very significant P<0.01, ***extremely significant P<0.001).
Results
Effect of resveratrol, H2O2, and their combined treatment on the viability and membrane integrity of human retinal pigment epithelial cells
Treatment with 500 μM H2O2 for 1 h decreased the cell viability by 57% (P<0.001). Resveratrol treatment alone did not cause cytotoxic effects or a significant extent of cell death. However, resveratrol pretreatment showed a statistically significant protective effect against H2O2-induced cytotoxicity (P<0.05) by increasing the cell viability by 24% at 25 μM and by 23% at 50 and 100 μM (Fig. 1a). Treatment with resveratrol alone did not result in a significant modification of LDH activity, the absorbance values being lower than the controls but not statistically significant. For cells treated with H2O2, LDH leakage increased by 11%, which is statistically significant compared with control (P<0.001). Pretreatment with resveratrol 25 and 50 μM, but not with 100 μM, resulted in restoring LDH leakage close to the corresponding control cells (P<0.05) (Fig. 1b).

Effect of resveratrol/and or hydrogen peroxide treatment on retinal pigment epithelium (RPE) cells viability
Effect of resveratrol treatment on the ROS generation under basal and oxidative stress conditions
Hydrogen peroxide treatment of RPE cells caused a significant increase of ROS generation compared with control (P<0.001). Resveratrol treatment alone did not significantly modify the ROS level. However, pretreatment with resveratrol followed by addition of H2O2 inhibited significantly the increase of the ROS level (P<0.001). The inhibition of ROS generation in resveratrol pretreated cells is directly correlated with the resveratrol concentration (Fig. 2).
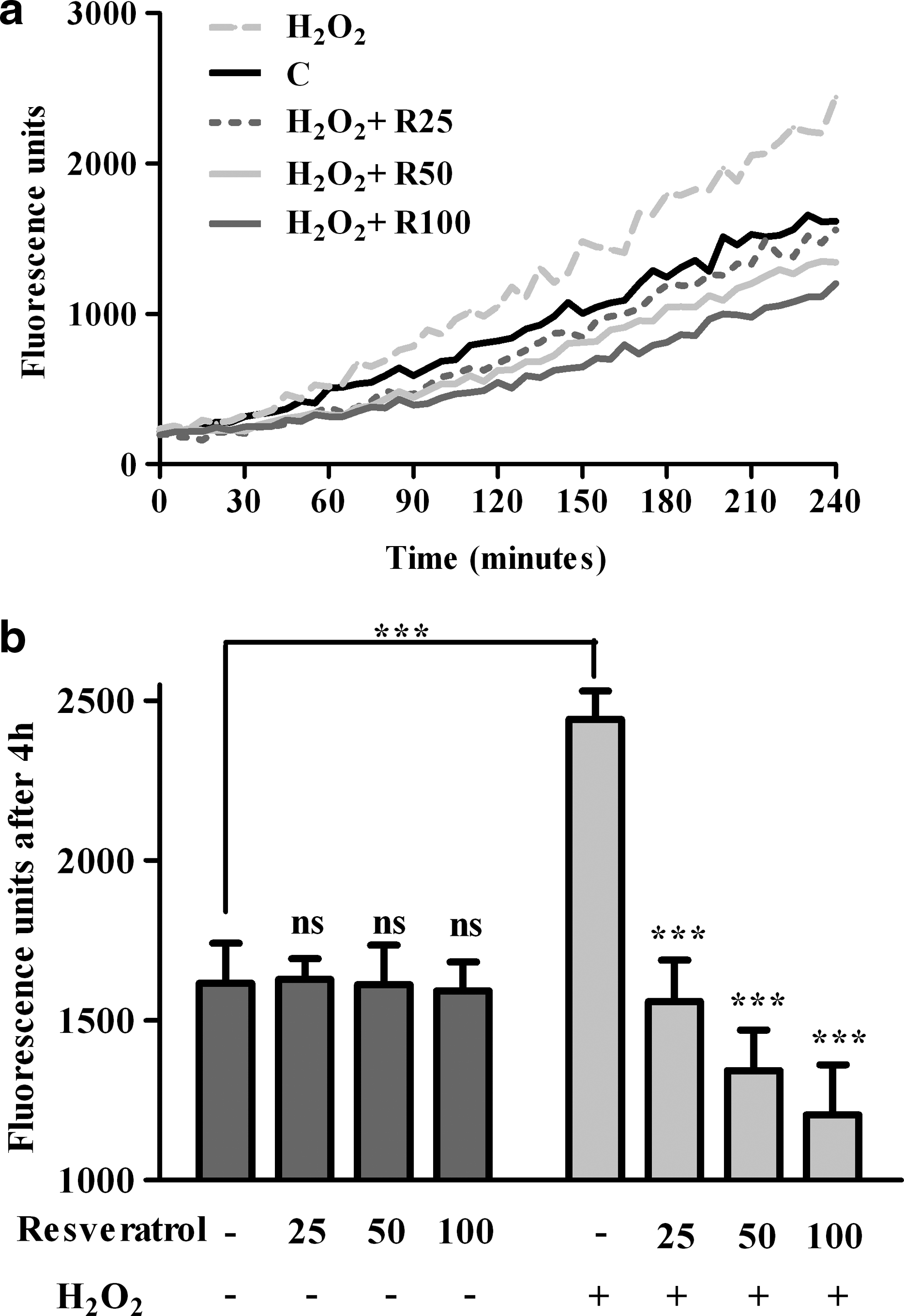
Fluorescence detection of intracellular reactive oxygen species in RPE cells recorded in time course
Antioxidant enzymes activities
SOD, peroxidase, and catalase are key enzymes of the antioxidant cell defense
SOD activity decreased by 67% (P<0.0001) after the addition of H2O2 compared with control cells. Resveratrol treatment alone at the concentration of 100 μM resulted in a significant (P<0.01) increase of the enzyme activity. In the case of cells exposed to oxidative stress and pretreated with resveratrol, the SOD activity increased significantly (P<0.01). The enzyme activity was only 10% lower for 25 μM resveratrol+H2O2–treated cells than in control cells (Fig. 3a).
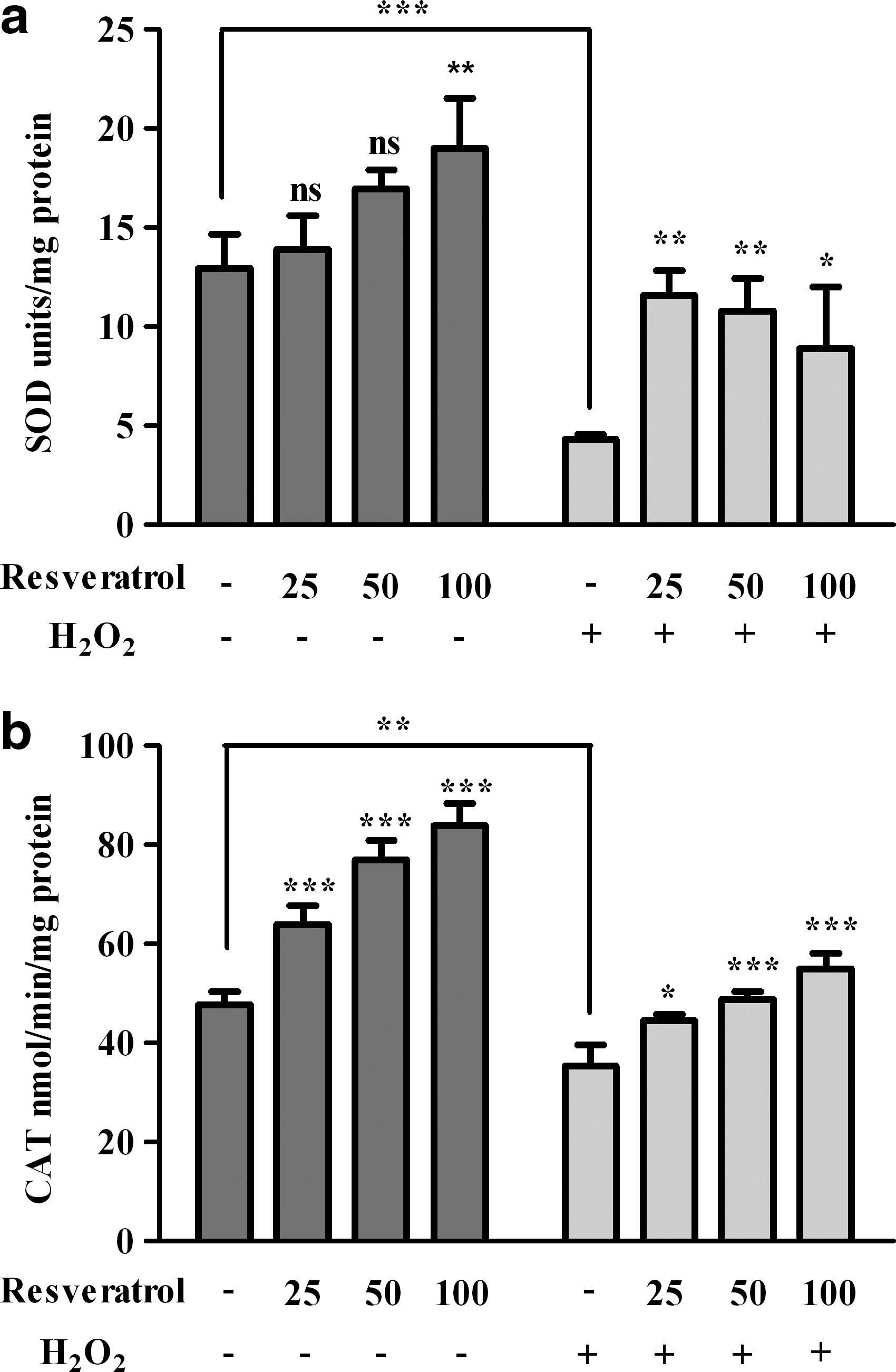
Effects of resveratrol/and or hydrogen peroxide treatment on superoxide dismutase (SOD)
Catalase activity was significantly reduced after H2O2 treatment (P<0.01). The enzyme activity was induced in both resveratrol and resveratrol+H2O2–treated cells significantly and dose dependently (P<0.001) (Fig. 3b).
As shown in Fig. 4, GPx activity was also decreased after H2O2 treatment (P<0.05). The addition of resveratrol to culture medium resulted in a significant increase of GPx activity, mostly for the 100 μM concentration (P<0.001). Hydrogen peroxide, after a 24 h pretreatment with resveratrol, caused an increase of GPx activity, mostly significant for the higher concentrations of resveratrol (50 and 100 μM, P<0.001).

Effects of resveratrol/and or hydrogen peroxide treatment on glutathione peroxidase (GPx) activity
Effect of resveratrol on reduced glutathione content
Treatment of the cells with 500 μM H2O2 alone induced a significant decrease of the GSH level (P<0.001). The intracellular GSH level did not increase significantly at low concentrations of resveratrol (25 and 50 μM) but was significantly induced for cells pretreated with 100 μM (P<0.01). The positive effect of resveratrol was more evident in cells exposed to H2O2, where the GSH levels increased 2.3-fold for 50 μM resveratrol and 2.4-fold for 100 μM resveratrol (Fig. 4).
Discussion
ROS play a major role in the mediation of cellular damage. Like other cells, RPE uses endogenous enzymatic and nonenzymatic antioxidants as a mechanism of ROS detoxification. Several antioxidant enzymes are involved in the primary or secondary antioxidant defense of RPE cells: catalase, GPx, glutathione S-transferase, CuZnSOD and MnSOD, heat shock protein 27 and 90, and heme oxygenase.18–21 RPE is also particularly rich in nonenzymatic antioxidants such as tocopherol, ascorbic acid, lutein, and zeaxanthin. 2 It was found that in RPE cells, the upregulation of antioxidants and an increased nuclear DNA protection play key roles in the adaptation to oxidative stress. 21 Supplementation with antioxidants represents a strategy to counteract the deleterious effects of ROS.
In the present study we used micromolar concentrations of resveratrol, according to previous studies regarding the antioxidant potential of resveratrol in various cultured cells.7–9,11,14,22 The results of our study indicate that resveratrol protects D407 RPE cells against hydrogen peroxide-induced cytotoxicity as evident from the viability (MTT), cytotoxicity (LDH leakage), and intracellular ROS generation assays.
Hydrogen peroxide is a ubiquitous oxidant found in ocular tissues in vivo; it is produced by the RPE during the phagocytosis of photoreceptors' outer segment.23,24 In this study we induced an acute, sublethal oxidative stress applying 500 μM H2O2 to the culture medium, in nonadapted RPE cells. At this concentration we observed a severe decrease of cell viability, a change in cell morphology (rounding) but not a severe change of cell density. Shamsi et al. reported that treatment of RPE cells with 100 μM hydrogen peroxide for 5 h determined significant changes in cell morphology and a cytopathic effect, whereas for 500 μM hydrogen peroxide they observed a 32% decrease of RPE cells' viability. 25 We showed that resveratrol pretreatment did not have cytotoxic effects in the absence of H2O2, but that it partially reversed the cytotoxic effects of H2O2 in cells exposed to oxidative stress (Fig. 1). Very recently, Sheu et al. demonstrated that resveratrol pretreatment of RPE cells alleviates the inhibition of phagocytosis in acrolein and combined acrolein/hydrogen peroxide-induced toxicity. 26 Other studies showed that resveratrol pretreatment conferred a marked protection against xanthine oxidase/xanthine, 4-hydroxy-2-nonenal, and doxorubicin-induced cytotoxicity in H9C2 cardiomyocytes cells, and against tert-butyl hydroperoxide-induced cytotoxicity in primary rat hepatocytes.7,9
Resveratrol was proved to have an intrinsic antioxidant capacity due to the redox properties of its hydroxyl groups and the delocalization of electrons. 27 The hydroxyl group in the 4′ position and the trans-configuration of resveratrol are structural determinants for the antioxidant activity of resveratrol. 28 Resveratrol was found to be an effective scavenger of hydroxyl, superoxide, and metal-induced radicals and to protect against lipid peroxidation and DNA damage caused by ROS in a cell model. 29 The antioxidant potential of resveratrol was demonstrated in several cultured cell models by inhibiting intracellular ROS generation.7,8,11 The present study demonstrated that resveratrol is a very potent inhibitor of intracellular ROS in RPE cells under oxidative stress conditions (Fig. 2).
SOD isoenzymes are present in human eyes, consisting of the extracellular SOD (EC-SOD), the cytosolic copper- and zinc-containing SOD (Cu/Zn-SOD, or SOD1), and the mitochondrial manganese-containing SOD (Mn-SOD). 30 SOD specifically catalyzes the dismutation of the superoxide anion to form oxygen and hydrogen peroxide. SOD enzymes work in conjunction with H2O2-removing enzymes, such as catalase and GPx. It was shown that the activity of RPE antioxidant enzymes, mainly catalase, decreases with aging in humans, but the expression of SOD, catalase, and heat shock proteins was increased in RPE from human donors suffering AMD.20,31 Our study showed that pretreatment of RPE culture cells with resveratrol increases the activity of the antioxidant enzymes SOD, GPx, and catalase. A significant increase of SOD activity was recorded for cells pretreated with resveratrol, especially when cells were challenged with H2O2 (Fig. 3a). Catalase is an essential enzyme for hydrogen peroxide detoxification in RPE because phagocytosis of rod outer segments leads to a high production of H2O2. It was shown that a short-term exposure (1 h) of nonadapted RPE cells to a higher concentration of H2O2 (3 mM) determined a very severe decrease in CAT (7-fold), GPx (2.1-fold), and CuZnSOD (37-fold) activities. 21 Under the experimental conditions used in our experiments, the activity of all antioxidant enzymes decreased in hydrogen peroxide-treated cells. Instead, we found a significant dose-dependent increase of catalase activity in RPE cells pretreated with resveratrol under both basal and oxidative stress conditions (Fig. 3b).
GSH has a crucial role in maintaining cellular redox status. GSH cooperates with GPx in the detoxification of H2O2. In addition, GSH participates in reactions with glutathione S-transferase to bind ROS, for example, attachment of ·NO to form S-nitrosoglutathione adducts. GR functions to regenerate the antioxidant capacity, converting glutathione disulfide (GSSG) into GSH. 24 In our study, the treatment of RPE cells with both resveratrol alone and resveratrol+H2O2 determined a significant increase of GSH concentration and of GPx activity (Fig. 4). The highest concentration of resveratrol (100 μM) restored the GSH content at the level of control cells. Similar results were obtained when cardiomyocytes, aortic smooth muscle cells, primary hepatocytes, and human erythrocytes were treated with resveratrol in the same range of concentration.7–9,32
Resveratrol is efficiently absorbed and metabolized very rapidly mainly into sulfo-conjugates and glucuronides, which are eliminated by urine. Consequently, the plasma concentration of free resveratrol is very low, even after high dosage oral administrations. However, due to its lipophilic nature, resveratrol may be bound to the cellular fraction in blood or in lipophilic tissues, which lead to the underestimation of its concentration. Despite the large number of studies, the distribution of resveratrol in tissues and cells, including the eye, remains to be determined. 33 Several studies on resveratrol pharmacokinetic and safety in humans showed that it is well tolerated and no marked toxicity was reported. 33 The highest dose was 5 g of resveratrol, as a single intake and it did not cause serious adverse effects. 34 Currently, several strategies are developed to improve the resveratrol bioavalability. 33
RPE is continuously subjected to the aggressive attack of ROS, resulting either from phagocytosis or from metabolic processes. It is considered that oxidative stress at RPE level is one of the major causes of AMD. This study demonstrates that resveratrol can contribute to the protection of RPE cells by enhancing antioxidant enzyme activities, by increasing the level of reduced glutathione and by inhibiting the intracellular ROS generation under oxidative stress conditions. More studies on the expression of antioxidant enzymes are needed to understand the mechanism of action of resveratrol at cellular levels in RPE cells. Taking into account the lack of toxicity, studies on human subjects are necessary to see if a resveratrol-rich diet can have a preventing effect on AMD.
Footnotes
Acknowledgment
This study was supported by PNCD II ID_854 Research Grant.
Author Disclosure Statement
No competing financial interests exist.