
Abstract
Abstract
Purpose:
The aim of this study was to investigate the dynamic alteration of a low-density lipoprotein receptor (LDLr) and its binding and uptake of LDL after exposure to transforming growth factor-β2 (TGF-β2) in Tenon's capsule fibroblasts from glaucoma patients.
Methods:
Tenon's capsule fibroblasts obtained from patients who had undergone selective glaucoma surgery were cultured and stimulated with different concentrations (0.1–10 ng/mL) of TGF-β2 for 1, 3, 5, and 7 days. Expression of LDLr mRNA was detected with relative quantification real-time reverse transcription–polymerase chain reaction, and expression of the corresponding protein was demonstrated with western blot. The extent of binding and uptake of 1,1′-dioctadecyl-3,3,3′,3′-tetramethyl-indocarbocyanine perchlorate (DiI)-labeled LDL by GTFs was also checked.
Results:
After exposure to TGF-β2, LDLr was upregulated at the mRNA and protein levels in a concentration-dependent and/or time-dependent manner with a peak at a concentration of 5 ng/mL TGF-β2. Binding and uptake of LDL were time dependent, reaching saturation at the first 6 h.
Conclusions:
This study shows that LDLr has a dose– and/or time–effect relationship with TGF-β2 in Tenon's capsule fibroblasts from glaucoma patients. The results suggest that LDLr may play an important role in wound healing and scar formation in the activated Tenon's capsule fibroblasts from patients with glaucoma.
Introduction
Recently, studies showed that constant nanoparticle drug delivery to targeted tissues and cells may offer a greater therapeutic effect than traditional dosing methods. 8 Our previous studies suggested that the LDLr was overexpressed in human Tenon's capsule fibroblasts from patients without a diagnosis of glaucoma (HTFs) activated by TGF-β2 9 and the photosensitizer accumulates mainly inside the activated HTFs via the overexpressed LDLr to cause phototoxicity. 10 Another investigation confirmed the results of photodynamic therapy in antiscarring after glaucoma filtering surgery in vivo. 11 An evaluation of the LDL-targeting paradigm with respect to glaucoma is therefore warranted to determine whether activated fibroblasts may be targeted by LDL. We assumed that addition of TGF-β2 might simulate the condition of conjunctival scarring after surgery for glaucoma. Our study investigated the alteration of LDLr with Tenon's capsule fibroblasts from patients with a diagnosis of glaucoma (GTFs) with and without the addition of growth factors.
Methods
Primary tissue procurement and culture
Biopsies of human Tenon's capsules were obtained from patients who had undergone glaucoma surgery (n=9; 5 females, 4 males; GTFs) and 3 male patients (aged 20, 35, and 62 without any topical eye treatment; HTFs) who had undergone trauma or cataract surgery. Patients' age ranged from 16 to 60 years with a mean age of 35.56 years (35.56±16.41 years). The research procedures followed the tenets of the Declaration of Helsinki. Informed consent was received from the patients. A total of 9 participants had no other disease except glaucoma and no other ocular surgery/trauma history. Of the 3 HTFs donors, 2 had trauma and 1 accepted cataract surgery. To establish explanted cultures, the biopsies were dissected into 3×3 mm sections and placed in 35-mm2 culture dishes (Falcon, Becton-Dickinson) in Dulbecco's modified Eagle's medium (DMEM; Gibco) containing 10% (v/v) fetal calf serum (FBS). Cells were allowed to migrate from the explanted tissue and were then incubated at 37°C in a humidified 95% air/5% CO2 atmosphere and fed twice a week. Cells between the third and sixth passages were used in this study. Cultures were allowed to reach about 80% confluence. After 24 h of serum starvation in serum-free DMEM, the cells were incubated in serum-free DMEM with or without human recombinant TGF-β2 (R&D Systems) for 1, 3, 5, and 7 days. Final concentrations of TGF-β2 were 0.1, 0.5, 1.0, 5.0, and 10 ng/mL (the levels of the aqueous samples from patients with glaucoma were 2.74±1.23 ng/mL after glaucoma filtering surgery). 12
Characterization of Tenon's capsule fibroblasts of patients with glaucoma
Cultured GTFs and HTFs (2×105 cells/coverslip) were plated onto sterile coverslips, placed in 6-well plates containing DMEM supplement with 10% FBS and then incubated for 3 days. To carry out immunostaining, cells were fixed in 4% paraformaldehyde (pH=7.4) for 5 min and permeabilized in 0.1% Triton X-100 for 5 min at room temperature. Coverslips were then washed in phosphate-buffered saline solution (PBS, pH=7.6). Cells were then incubated in the primary antibodies, recognizing monoclonal mouse antivimentin (1:200 dilution; Sigma-Aldrich) and monoclonal rabbit antifibronectin (1:150 dilution; Sigma-Aldrich) overnight at 4°C. Then the coverslips were washed in PBS, incubated with secondary antibodies for 1 h at room temperature, and counterstained with 4′,6-diamino-2-phenylindole (DAPI; Invitrogen) for 5 min at room temperature. The coverslips were washed in PBS, dehydrated, and mounted on glass slides. The samples were viewed and photographed using a confocal microscope (510 UV–vis Meta, Zeiss).
MTS assay
GTFs and HTFs were plated in 96-well plates (Falcon) at a density of 4.0×104 cells/well and stimulated with different concentrations of TGF-β2. The 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) assay was performed by adding 20 μL of CellTiter 96 Aqueous One Solution Reagent (Promega Corporation) per well with 100 μL serum-free DMEM and then incubated at 37°C in humidified 95% air/5% CO2 for 4 h. The optical density of the fluid in each well was read at 490 nm in a multiwell spectrophotometer (Titertek Multiscan, Flow Lab). Wells containing basic medium served as controls.
Quantitative real-time reverse transcription–polymerase chain reaction
The expression of the LDLr mRNA was examined with quantitative real-time reverse transcription–polymerase chain reaction (RT-PCR). In brief, GTFs and HTFs were plated in 60-mm2 dishes (Falcon) at a density of 2.5×104 cells/dish and stimulated as earlier. Total RNA was extracted from fibroblasts using the Trizol RNA extraction reagent (Invitrogen Corp.) according to the manufacturer's instructions. Purified RNA was reverse-transcribed using a SYBR PrimeScriptTM RT-PCR Kit (TaKaRa Corp.). Real-time quantification of LDLr mRNA was performed on an ABI PRISM 7000 Sequence Detection System using SYBR Green I as the reporter dye (TaKaRa Corp.). The comparative Ct method was employed, whereby the relative quantity of the target gene mRNA, normalized to GAPDH and relative to the calibrator, was expressed as fold change=2−ΔΔCt. The following human LDLr and GAPDH oligonucleotide primers were used in the real-time quantitative PCR: LDLr-F: 5′-CCG TCA GGC TAAA AGG TCAG -3′; LDLr-R: 5′-TTC GCC GCT GTG ACA CTTG-3′; GAPDH-F: 5′-GCA CCG TCA AGG CTG AGAAC-3′; GAPDH-R: 5′-TGG TGA AGA CGC CAG TGG A-3′. Duplicate PCRs were tested using the following amplification protocol: 95°C for 10 s followed by 38 cycles at 95°C for 5 s and at 60°C for 31 s.
Western blot
Whole cellular proteins were isolated from primary cultured GTFs and HTFs. Briefly, total cell lysates were obtained using lysis buffer containing 1.6% Triton X-100, 5 M urea, 0.1 mM leupeptin, and 1.5 mM phenylmethylsulfonyl fluoride. After centrifugation at 12,000 g for 30 min, the supernatant was removed and stored at −80°C. Protein concentrations were measured using a BCA assay kit according to the manufacturer's instructions (Shenergy Biocolor BioScience and Technology Company). Equal amounts of protein (20 μg) were subjected to 8% sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to PVDF membranes, which were blocked in 5% skim milk/PBS + 0.05% Tween-20. The membranes were probed with primary antibodies against human LDLr (1:1,000 dilution; Biovision) and β-actin (1:1,000 dilution; Cell Signaling technology) following the ECL-western blot protocol (Amersham Life Sciences), and the secondary antibodies were HRP-conjugated goat anti-rabbit (1:1,000 dilution; Cell Signaling Technology). The enhanced chemiluminescence (ECL) detection method was used, and blots were exposed to film for 30 s. Signals were quantified using the Image Quant ImageAnalysis Software (Amersham Biosciences).
Isolation of LDL from human plasma
Purified LDL was isolated from the plasma of healthy volunteers with sequential ultracentrifugation (d=1.019–1.063 g/mL) and labeled with the fluorescent probe, 1,1′-dioctadecyl-3,3,3′,3′-tetramethyl-indocarbocyanine perchlorate (DiI), in the presence of 3 mM ethylenediaminetetraacetic acid (EDTA). The resultant product was exhaustively dialyzed against PBS (pH 7.4), sterilized with a 0.22-μm filter, then aseptically packaged in a solution containing PBS at pH 7.4 and 0.02 mM EDTA, and stored under sterile conditions at 4°C.
LDL uptake assay
GTFs and HTFs were plated in 30-mm2 dishes (Falcon) at a density of 2.0×104 cells/dish and stimulated with a concentration of TGF-β2 (the best final concentration and incubation time according to the MTS assay results) in DMEM supplemented with 0.05% FBS. Labeled LDL (DiI) was examined via confocal microscopy using a Zeiss 510 UV–vis Meta microscope at 3, 6, and 9 h. DiI-LDL (10 μg/mL protein) was added to cells at the start of the experiment, and the cells were incubated at 37°C in a 5% CO2 incubator. The group cultured with basic medium served as controls. Cells were viewed at 40×magnification. Confocal microscopy was also used to measure the DiI fluorescence intensity in the 2 groups of GTFs and HTFs.
Data analysis
Analysis was performed using computer software (SPSS for Windows; SPSS 13.0) at individual time points using a one-way analysis of variance followed by least significant difference (LSD) analysis. The correlations were evaluated with linear correlation analysis. The values were expressed as mean±standard deviation (SD). All data were significantly different at P<0.05.
Results
Morphological and immunocytochemical characteristics of fibroblasts
GTFs and HTFs were spindle-shaped in culture as reported. Immunocytochemically (Fig. 1), cultures were 100% positive for vimentin and fibronectin. Vimentin, along with microtubules and actin microfilaments, make up the cytoskeleton. Vimentin exists as a dynamic structure in fibroblasts. Fibronectin may enhance cell adhesion and spreading. It affects the routes of cell migration both in vivo and in vitro. Vimentin and fibronectin are classic markers for primary human fibroblasts and related to many vital cell functions such as cell motility, chemotactic migration, and wound healing. The total cell counts are shown in Fig. 1 after staining by DAPI.
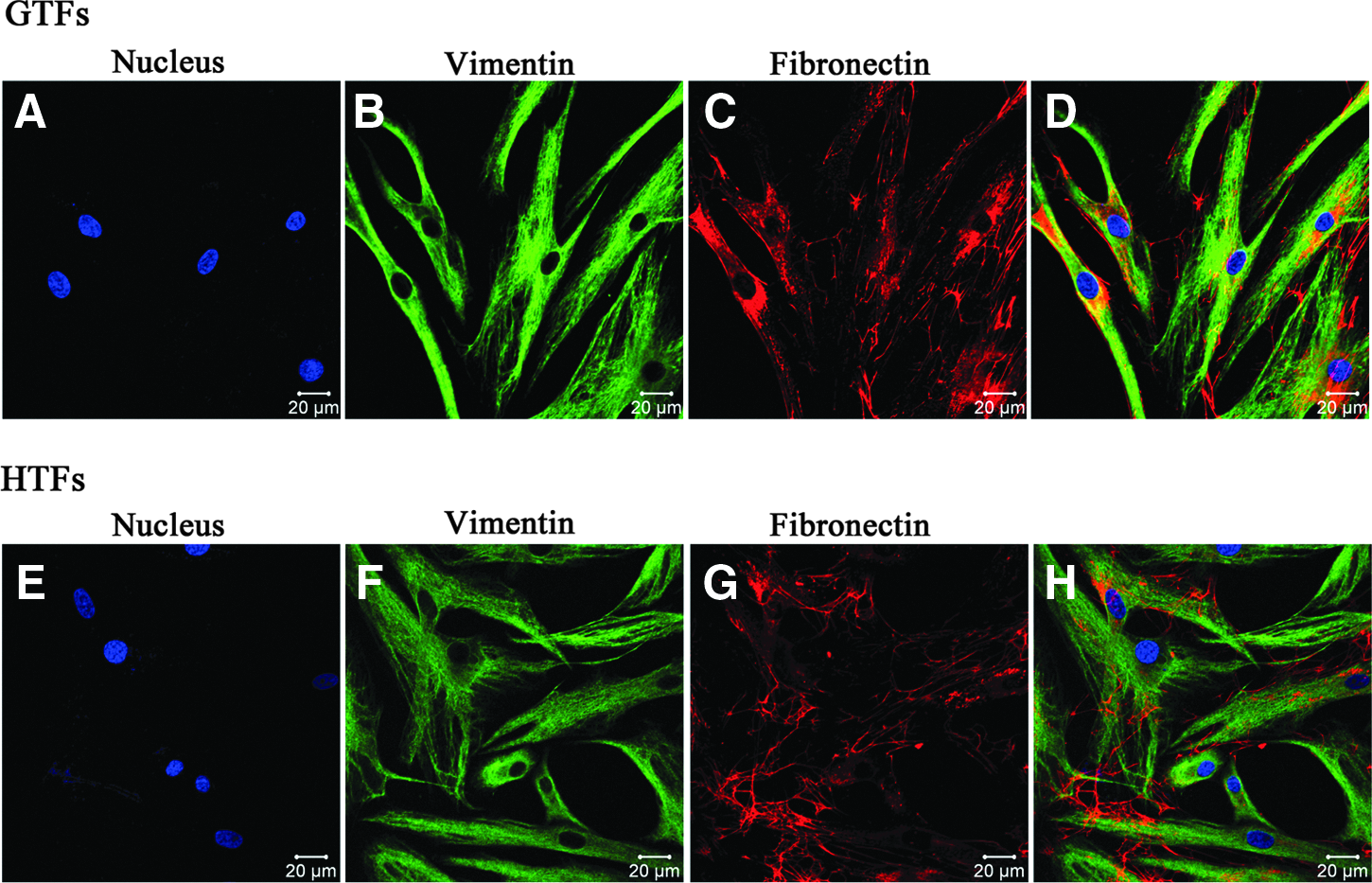
Identification of cultured primary GTFs and HTFs with vimentin (green) and fibronectin (red). Negative control using FITC-marked secondary antibody and Cy3-marked secondary antibody of GTFs
Effect of TGF-β2 on fibroblasts proliferation
The MTS assay was used to determine cell viability. The effect of TGF-β2 on the proliferation of cultured GTFs/HTFs in different concentrations is summarized in Fig. 2. The cell proliferation of GTFs/HTFs exposed to TGF-β2 occurred in a concentration-dependent and time-dependent manner in vitro. The greatest activity appeared at the concentration of 5.0 ng/mL, with the peak proliferation rate of 143% in GTFs. All concentrations significantly stimulated GTF/HTF proliferation compared with their own control after days 1, 3, 5, and 7 (P<0.05). For concentrations of 5 and 10 ng/mL, significant differences (P<0.05) were found between proliferation on days 3, 5, and 7 after treatment in GTFs. Significant differences were also found between day 1 and days 3, 5, and 7 at the concentration of 5 and 10 ng/mL in HTFs.
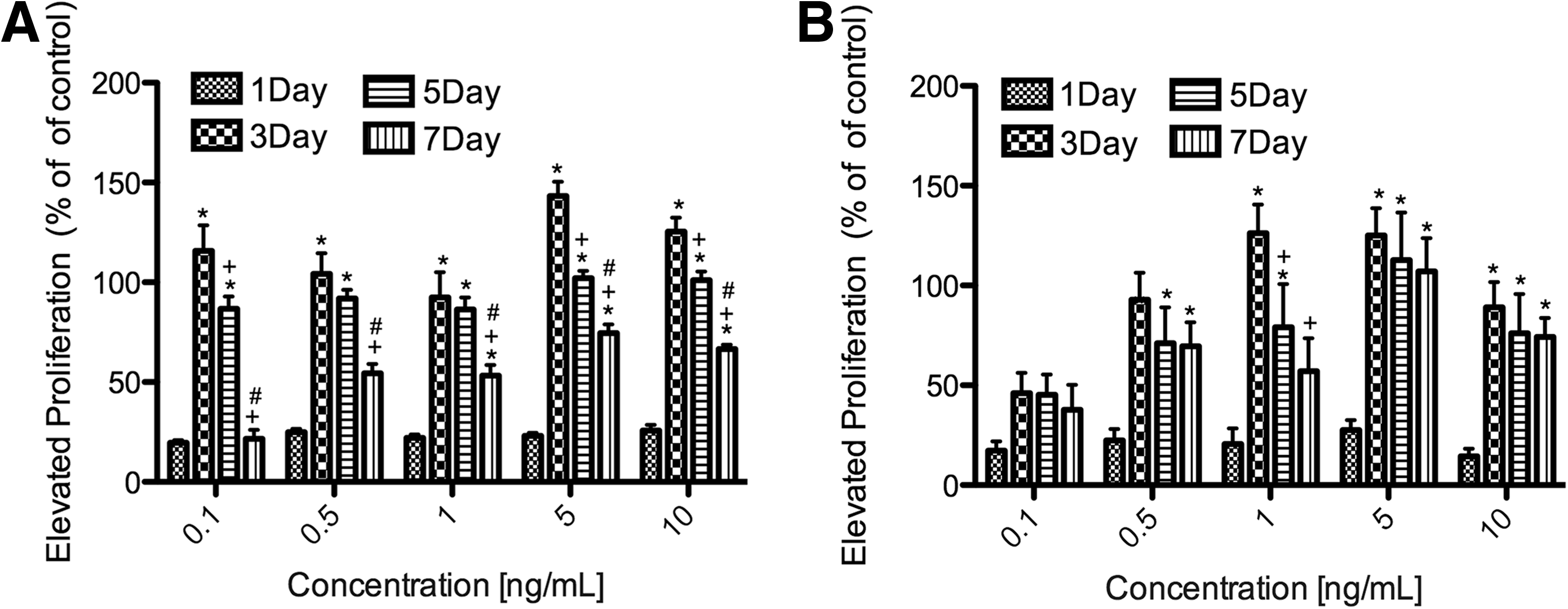
GTF
Expression of LDLr after exposure to TGF-β2 in fibroblasts
To determine whether LDLr expression was elevated after exposure to TGF-β2 in GTFs/HTFs, we used quantitative real-time RT-PCR to detect LDLr mRNA and western blot analysis to examine the LDLr protein. The results showed concentration-dependent and time-dependent upregulation of LDLr mRNA (Fig. 3) and protein (Fig. 4) expression at 5 concentrations of TGF-β2. Similar to the corresponding rates of cell proliferation, the mRNA and the protein levels of LDLr reached their peaks at 5 ng/mL TGF-β2 after 3 days of culture in GTFs and then decreased. At the peak on day 3, the mRNA level of GTFs had increased 18-fold, whereas the mRNA level of HTFs had increased 5-fold. The protein levels of LDLr reached 26-fold above in GTFs, whereas the protein levels of LDLr reached 8-fold above in HTFs.
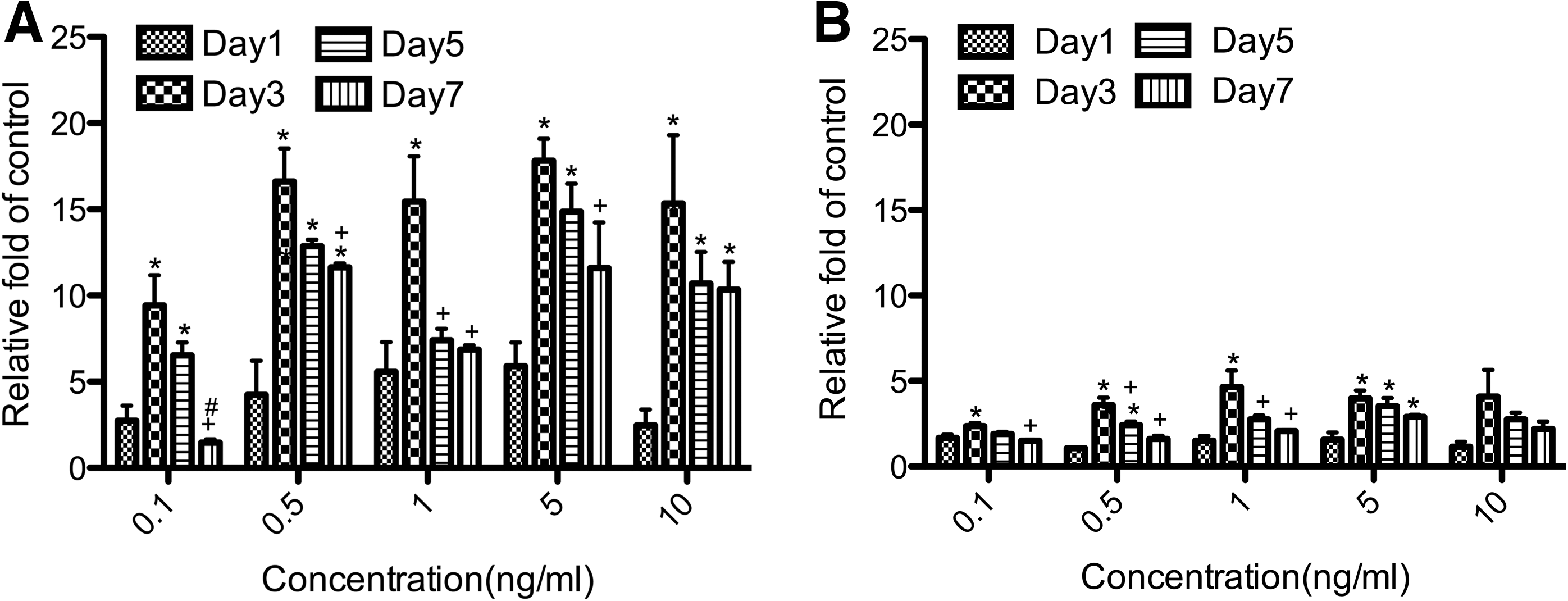
Increased level of LDLr mRNA in the activated GTFs
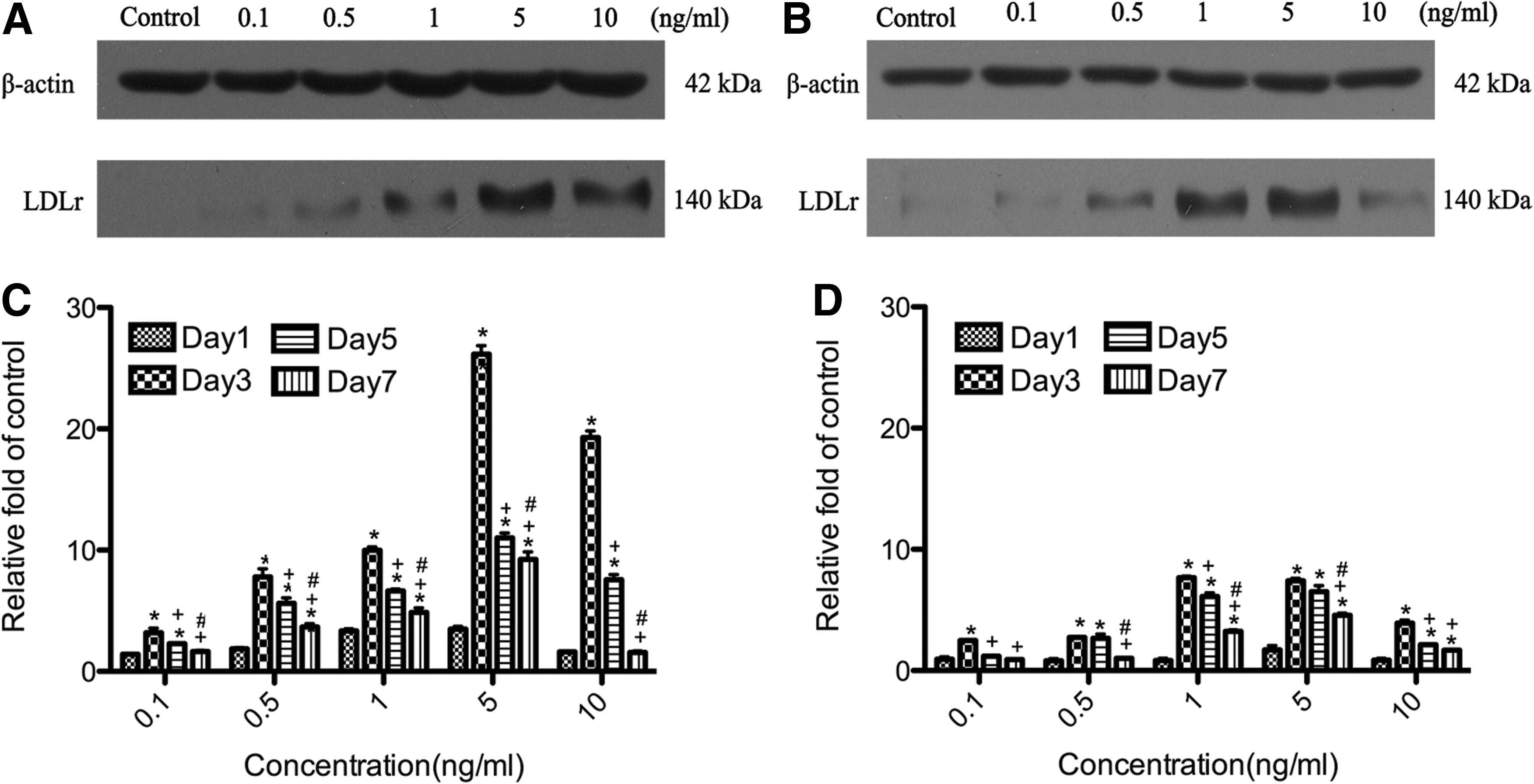
Dynamic alteration of LDLr protein expression in the activated GTFs and HTFs after exposure to TGF-β2 with the western blot method using monoclonal antibodies against LDLr and β-actin on the traditional 1D sodium dodecyl sulfate–polyacrylamide gel electrophoresis in total cell proteins. β-Actin expression was used as an internal control. Representative western blot bands for the relative expression of LDLr protein in all groups of GTFs
Correlation between expression of LDLr and proliferation of fibroblasts
Linear correlation analysis showed a significant positive correlation between the LDLr mRNA level and proliferation level (r=0.567; P<0.01) and between the LDLr protein level and the proliferation level of GTFs (r=0.620; P<0.01), respectively (Fig. 5A). There was also a significant positive correlation between the LDLr mRNA level and the proliferation level (r=0.425; P<0.01) and between the LDLr protein level and proliferation level of HTFs (r=0.602; P<0.01), respectively (Fig. 5B).
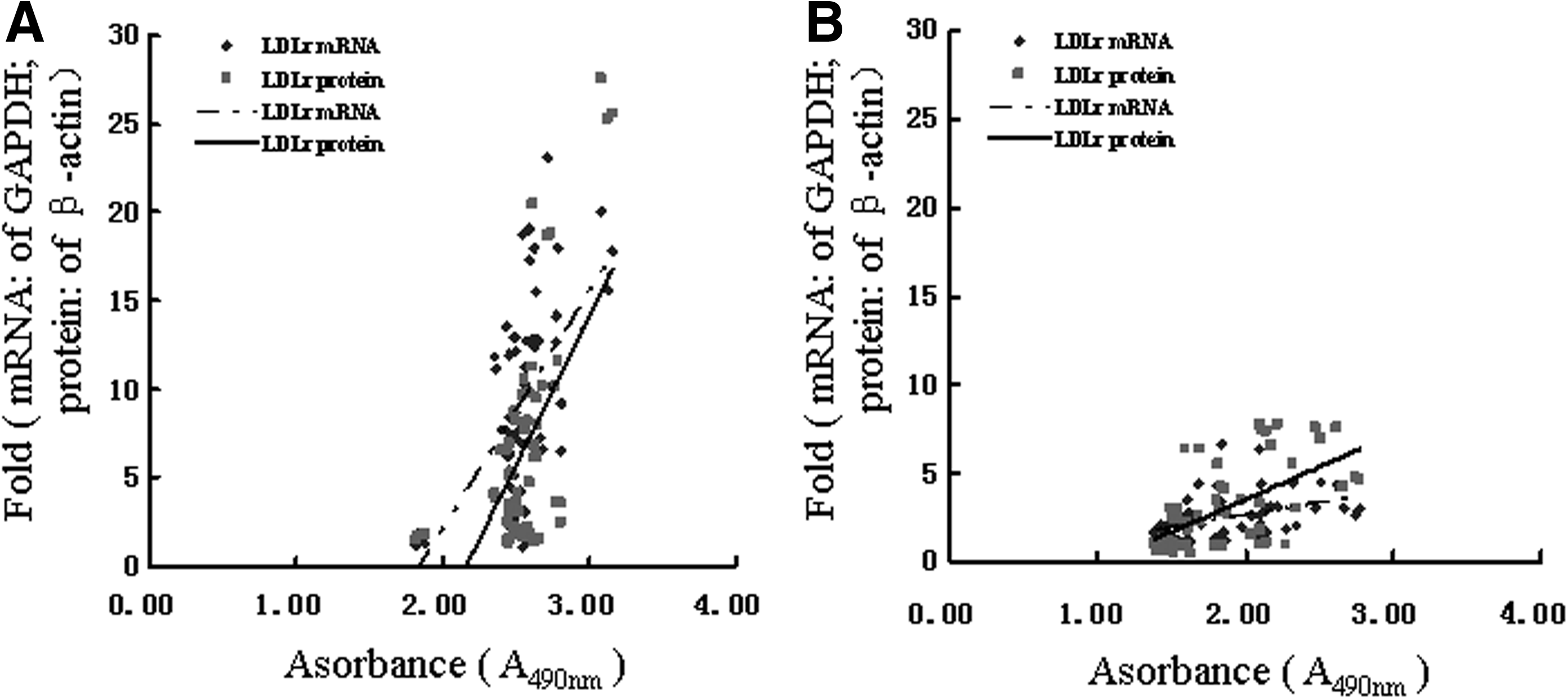
Correlation between the expression of LDLr (mRNA and protein level) and the proliferation in GTFs
Uptake of DiI-LDL
Uptake of the DiI-labeled LDL particles (10 μg/mL protein) by living GTFs/HTFs was assessed by incubating cells at 37°C for 3, 6, and 9 h before confocal microscopy examinations were carried out. A representative confocal microscopic image series of the activated GTFs and the control is shown in Fig. 6. DiI-LDL uptake increased in a time-dependent manner and appeared to saturate at 6 h. In addition, at every time point, especially at 6 h, TGF-β2 resulted in higher uptake of DiI-LDL particles by GTFs than that by the control. This difference is due to either the amount or activity of LDLr in GTFs. At 9 h, the fluorescence of the control decreased, whereas the treated cells remained at a high level. Uptake of the DiI-labeled LDL particles (10 μg/mL protein) by living HTFs showed a similar way with a slightly lower peak than in the GTFs. The experiment was repeated with a separately produced batch of LDL and yielded identical results.
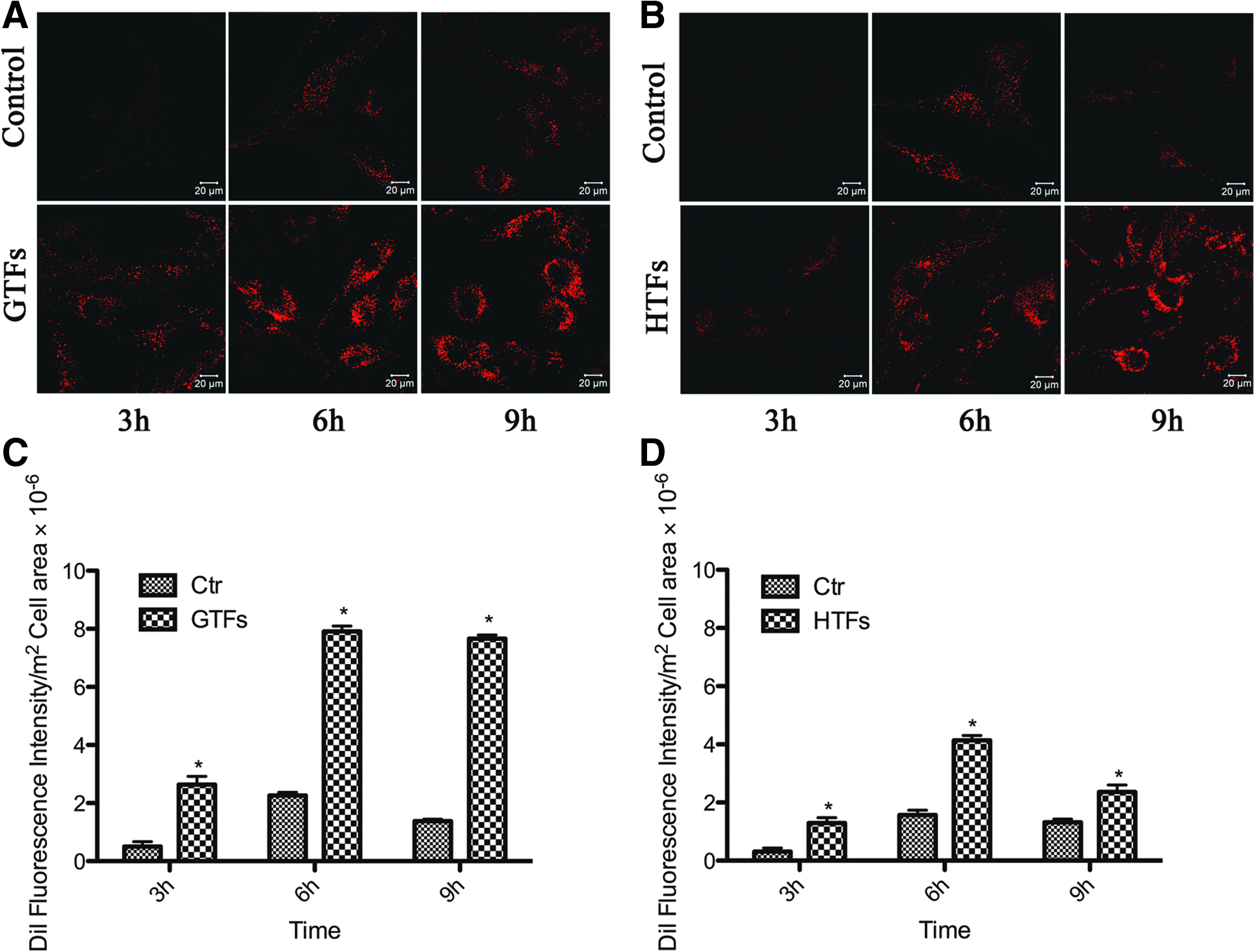
Representative experiments illustrating that DiI–LDL uptake by living GTFs and HTFs was time-dependent. Confocal microscopy showed uptake of DiI-labeled LDL (10 μg/mL protein) by activated GTFs
Discussion
It is well established today that the activation of Tenon's capsule fibroblasts is the central regulatory element of the scarring reaction.13–15 Activation of Tenon's capsule fibroblasts includes mitogenic stimulation, phenotypic transition to myofibroblasts, and stimulation of ECM gene expression and secretion.16–18 A large body of evidence shows that growth factors play a major role in regulating these events. It has been demonstrated that TGF-β2 can function as a growth stimulator only for fibroblastic cells. This effect is indirect and is mediated via the induction of the sis-proto-oncogene that codes for the B chain of PDGF. 19 The present study confirms that both GTFs and HTFs are efficiently stimulated by TGF-β2 in a concentration-dependent and/or time-dependent manner. TGF-β2 stimulates DNA synthesis in quiescent substratum-attached cultures of mouse AKR-2B cells without other added growth factors, but with delayed kinetics, compared with stimulation with other growth factors. 20 In our study the peak proliferation rate appeared at 3 days postexposure to TGF-β2, which is in agreement with a previous investigation. 21 Cell growth stimulated by TGF-β2 brings about alterations in the functions of the cell membrane. Several studies reported that the uptake of glucose and amino acids into the cells is enhanced by TGF-β2.22,23 However, the relationships of these membrane changes to growth stimulation are not known.
Our results show that there is a significant positive correlation between expression of LDLr and proliferation rate whether in GTFs or HTFs. The expression of LDLr and cell proliferation in activated GTFs was markedly elevated just like the manner of activated HTFs after exposure to TGF-β2. It is reasonable to postulate that LDLr expression in cultured cells relies on the growth rate. This is further supported by a previous study. 24 The LDLr is the prototype of a large family of structurally homologous cell surface receptors, which fold in the ER and function as endocytic and signaling receptors in a wide variety of cellular processes. Tumor tissues display elevated receptor-mediated uptake of LDL compared with normal tissues. 25 What interested us most was that our results showed that LDLr protein level increased relatively higher than the mRNA level. Presumably, there are multiple ways for the regulation of LDLr expression in these activated fibroblasts. This is further supported by previous in vitro studies of cultured cells.26–28
There are many possible sources of TGF-β2 at the wound site in vivo and local. The concentration may be much higher in the microenvironment of fibroblasts than in wound fluid because of paracrine secretion by platelets, inflammatory cells, and even the fibroblasts themselves. In fact, during conjunctival wound healing, there is a variety of mediators, not only growth factors but also activated complement products, proinflammatory cytokines, and vasoactive factors, to stimulate the fibroblasts that reinforce each other to activate fibroblasts, resulting in a much higher proliferation of fibroblasts. 29 Therefore, higher postoperative overexpression of LDLr in the activated fibroblasts is expected.
The present study also showed that uptake of the DiI-labeled LDL particles by living cells increased in a time-dependent manner in GTFs and HTFs with the addition of TGF-β2. At the early stage, the DiI-labeled LDL was located under the apical plasma membrane and then translocated and accumulated in the cytoplasm. We also found that there was a significant difference in the amounts of LDLr between the activated GTFs and the control. Similarly, the peak value in HTFs was higher than its control. As LDL, via a specific receptor-mediated uptake, was internalized into cells, the cells can absorb a large amount of LDL within a relatively short time. With a recycle time of up to 10 min and a diurnal time of 24 h, one can assume that each receptor mediates the transport of up to 144 LDL particles into LDLr-expressing cells per day. This provides an extremely efficient system for delivering a natural cargo of cholesterol.
In vitro experiments suggest that human conjunctival fibroblasts are sensitive to mitomycin C in an incremental fashion. 30 Hence, the elevated expression of LDLr could be of potential interest for targeted delivery of drugs, that is, as the signal amplification mechanism in the process of receptor recycling.
Conclusions
In summary, our results suggest that LDLr may play an important role in wound healing and scar formation in the activated Tenon's capsule fibroblasts from glaucoma patients. This finding may offer a new insight for more precise and rational design of conjunctive drug and/or gene delivery nanosystems. As for glaucoma patients, LDL antiscarring drug delivery vehicle system may be a novel way in reducing future bleb fibrosis and enhancing the success rate of surgery.
Footnotes
Acknowledgments
This study was supported by grants from the National Basic Research Program (973) of China (No. 2007CB512207) and the National Natural Science Foundation of China (No. 30672275, No. 30400486).
Author Disclosure Statement
No competing financial interests exist. The authors alone are responsible for the content and writing of the article.