
Abstract
Abstract
Purpose:
PF-655, a synthetic 19-mer siRNA, targeting the RTP801 gene is currently in clinical trials for the treatment of wet age-related macular degeneration and diabetic macular edema. Preclinical studies have shown a dose-related suppression of RTP801 expression in rat disease models. Investigative studies were conducted with PF-655 to validate the Dutch-Belted rabbit as a biologically relevant species for gene silencing to support nonclinical ocular toxicity and continual dosing studies.
Methods:
Cross-species comparison and DNA sequencing was done to determine the level of homology between PF-655 and rabbit RTP801. Human (HEK 293) and rabbit (SIRC cornea) cell lines were stimulated with CoCl2 to mimic hypoxic stress (an inducer of RTP801 expression) and treated with PF-655. Taqman-polymerase chain reaction and immunoblot analysis were performed to gauge RTP801 expression in cell culture and rabbit retinas.
Results:
Sequence analysis showed a 1-base mismatch in the PF-655 targeting site from genomic DNA of Dutch-Belted rabbit and the SIRC cell line, a cornea cell derived from the New Zealand White rabbit. HEK and SIRC CoCl2-stressed cells induced RTP801 expression 10–20-fold above control conditions. Treatment with 20 or 100 nM PF-655 showed a decrease in gene expression, 40%–50% relative to appropriate controls. RTP801 mRNA was detectable in primary rabbit retina tissues, with cycle threshold values showing a large linear range for the assay.
Conclusion:
These results support our investigation into cross-species validation of gene suppression by a therapeutic siRNA designed to a human gene. The SIRC cell line was utilized as a surrogate to test the degree of RTP801 gene silencing induced by PF-655 in vitro. With a 1-base mismatch, the level of silencing in a rabbit ocular cell line was comparable to that of a human cell line. Sequence analysis and expression data confirmed the relevance of the RTP801 target gene in rabbits and the utility of this species as a relevant animal model. Additionally, our work outlines a tractable method that validates relevant larger non-rodent species for ophthalmic drug testing.
Introduction
The RTP801 gene (also termed REDD1 and DDIT4) was identified independently by 3 research groups studying genes involved in a variety of cellular stress responses such as hypoxia, ER stress, reactive oxygen species, and DNA damage.11–13 More recent studies have shown that RTP801 plays a role in the TSC1/TSC2-mediated inhibition of mammalian target of rapamycin (mTOR) and is involved in regulating cellular responses in a variety of cell types such as neurons and skeletal muscle.14–16 A critical role for RTP801 in the pathogenesis of ocular disease was revealed in a study utilizing RTP801 knockout mice. 17 In the paper, the authors used a model of high oxygen exposure in newborn mouse pups and subsequent recovery in a normal air environment to induce abnormal vascularization and retinopathy or retinopathy of prematurity. RTP801 deficient mice had decreased levels of retinal neovascularization and attenuated some of the pathogenesis in their ocular disease model. This article along with others, which indicate that RTP801 plays a critical role in the pathological process of acute and chronic stress-induced disease and potentiates VEGF expression during hypoxic stress, make this gene an attractive target for siRNA therapeutic intervention within the eye.18,19
Since siRNAs target genes for mRNA degradation based on DNA sequence identity, cross-species gene comparison becomes important when selecting animals for drug testing. In addition, for continual ocular dosing and long-term ocular toxicity studies, larger animal species such as dog, pig, or rabbit may be required to satisfy regulatory requirements. 20 PF-655, a siRNA targeting RTP801, is in development for the treatment of DME and wet-AMD. In this study, we assessed whether PF-655, designed to target the human RTP801 gene, can be used to knockdown expression of the homologous rabbit gene in order to validate this species for nonclinical ocular safety and continual dosing studies. The data indicate that expression of RTP801 can be induced under acute hypoxic stress in rabbit and human cells. Even with a 1 base mis-match in the siRNA hybridization targeting site, PF-655 is able to silence the RTP801 gene in rabbit species down to levels similar to what was experimentally observed in human cells. Our results point to an induction pathway that is conserved between rabbit and human species for the regulated expression of RTP801 and validate the rabbit species as an appropriate non-rodent model for further preclinical investigation with PF-655. Additionally, our work outlines a tractable method that validates larger non-rodent species for ophthalmic testing of siRNA drug candidates.
Methods
Cell culture and siRNA transfection
The cell lines 293T and SIRC were obtained from ATCC (Manassas, VA), and cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum, L-glutamine, and penicillin/streptomycin antibiotics. Cells were grown on standard tissue-culture grade plastic dishes and 6-well plates in 5% CO2 humidified chambers. 16–18 h before RNAi transfection, the cells were washed with 1× phosphate-buffered saline, trypsinized, and re-plated onto 6-well culture dishes at 40%–50% confluence. Transfection was performed using Lipofectamine 2000 reagent (Invitrogen) according to the manufacturer's instructions. CoCl2 was obtained from Sigma Chemical and resuspended in RNase-free H2O. HEK 293 and SIRC cells were treated with CoCl2 for 3–9 h at a final concentration of 100 μM. The phosphoinositide-3 kinase (PI3K) inhibitor, LY294002, was obtained from Cell Signaling Technologies. Stimulation with CoCl2 or treatment with LY294002 was done 48 h post-transfection.
Sequence information for the siRNA oligos is as follows;
PF-655-5′ GUGCCAACCUGAUGCAGCU (licensed and patent owned by Quark Pharmaceuticals).
Nonspecific siRNA control–5′ UAAGGCUAUGAAGAGAUAC (sequence has no significant homology to any known mammalian genes).
Cellular fluorescence analysis
A fluorescein isothiocyanate (FITC)-labeled siRNA oligo duplex corresponding to PF-655 sequence was synthesized by IDT DNA. Microscopy was performed using an inverted fluorescence microscope (Olympus). Flow cytometry was performed using the Accuri C6 cytometer (BD Biosciences).
Cell viability assay
The CellTiter Glo assay (Promega) was used to assess cell viability following the manufacturer's protocol, and the signal was captured on a 96-well plate reader (Tecan).
Immunoblot analysis
Protein lysates were prepared using radio-immunoprecipitation assay (RIPA) buffer supplemented with protease inhibitors (Pierce). Sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transfer was performed using 10–20 μg of whole cell lysate per sample on a 4%–12% Bis-Tris gel using the Invitrogen XCell II Mini-Lock system. All steps were done according to the manufacturer's instructions. Antibodies for RTP801 (DDIT4) and β-actin were obtained from ProteinTech Group and Santa Cruz Biotechnology, respectively. IRDye conjugated secondary antibodies were obtained from Rockland Immunochemicals. The LiCor Odyssey platform was used to detect western signals and capture images.
Taqman-polymerase chain reaction
Total RNA was purified using the RNeasy mini kit (Qiagen). First-strand cDNA synthesis was performed using a cDNA reverse transcription kit (Applied Biosystems). Real-time quantitative polymerase chain reaction (q-PCR) was performed in duplicate using the Roche LightCycler 480 platform, reagents, and the following probe/primer sequences (5′ to 3′):
Human RTP801-FAM-CCGACAGTGCCCTCCAAGACAG-TAMRA; CACTGGCTTCCGAGTCATC; TCCACCCCAAAAGTTCAGTC
Human β-actin: FAM-ATCAAGATCATTGCTCCTCCTGAGCGC-TAMRA; CCTGGCACCCAGCACAAT; GCCGATCCACACGGAGTACT
Rabbit RTP801: FAM-CACGAGGGTCAGCTGGAAGGT-TAMRA; GTGGAGCAGGGCAAGAG; AAGGTTGAGTTGGCCGAG
Rabbit β-actin: FAM-CACGGAGCGCGGCTACAGCTTC-TAMRA; CCTGACCGACTACCTCATGAAGA; TTGATGTCCCGCACGATCT
Primers for examining the expression of interferon (IFN) response genes were purchased from Invivogen and used according to the manufacturer's instructions.
Relative gene expression was calculated using the delta-delta Ct (2−ΔΔCt) method with β-actin serving as the reference gene for RTP801 and GAPDH as the reference for the IFN response genes.
DNA sequencing
DNA was isolated from SIRC cells and whole-blood drawn from Dutch-Belted rabbit using the mammalian Genomic DNA Miniprep kit (Sigma-Aldrich). Primers were designed to flank the PF-655 targeting site in exon 3 of the RTP801 gene in rabbit species. PCR was carried out using 200 ng of genomic DNA from each rabbit species as template plus standard buffer reagents and Taq polymerase (Invitrogen) and oligonucleotides 5′ TACCTGGACGGGGTGTCCCTGCC and 5′ CGCAGGCCGCACGGCTCGCTGTA. The resulting genomic DNA fragment of ∼250 base pairs was purified by agarose gel excision (Qiagen) and sent to a contract lab for Sanger sequencing (GeneWiz).
Results
A partial mRNA sequence of the RTP801 gene from the rabbit species, Oryctolagus cuniculus, was obtained from the Ensembl Genome website (www.ensembl.org) and aligned to the homologous human gene (NCBI Accession NM_019058) (data not shown). The mRNA target site of PF-655 siRNA was included in the partial sequence alignment and differed from the human sequence by 1 base at the number 7 position within the 19-nucleotide siRNA region (Fig. 1A). The human sequence and PF-655 both have an adenine at this position, while the rabbit has a cytosine, giving the rabbit sequence a 94.7% identity compared with humans.

A single nucleotide mis-match exists in the PF-655 small interfering (siRNA) target site of the RTP801 gene in the Dutch-Belted and New Zealand White rabbit species.
To investigate whether PF-655 could silence the rabbit RTP801 gene, we chose to use the SIRC cell line. The cell line is derived from cornea tissue of the New Zealand white rabbit and amendable to transfection studies in cell culture. Since ocular drug studies in rabbit species are usually performed in the pigmented eyes of Dutch-Belted rabbits, we next performed DNA sequencing on the RTP801 gene using genomic DNA from SIRC cells and blood from Dutch-Belted.21,22 As shown by the DNA sequence chromatograms, the Dutch-Belted rabbit has the same PF-655 siRNA target sequence (Fig. 1B, upper panel DNA sequence chromatogram) compared with the SIRC cell line (Fig. 1B, lower panel chromatogram).
RTP801 was originally cloned as a stress responsive gene, and its expression is dependent on PI3K activity under hypoxic conditions.15,18 To determine whether we could see induction of RTP801 in our human and rabbit cell culture model system, we used CoCl2 treatment to mimic hypoxia. 23 HEK 293 and SIRC cells were treated with CoCl2 in culture for 1 or 3 h with or without the presence of LY294002, a PI3K inhibitor. Immunoblot analysis showed low to undetectable levels of RTP801 in untreated cells, while the protein was induced in a time-dependent manner in both human and rabbit cells (Fig. 2A, B). Treatment with LY294002 abolished the CoCl2-induction of RTP801, confirming that upregulation of this gene requires PI3K activity and this pathway dependence is conserved in the rabbit.

Protein expression of RTP801 can be induced with hypoxia-like stimulation in human and rabbit cell lines and is dependent on PI3K activity.
To determine the proportion of cells that will take up the siRNA for gene silencing, PF-655 was synthesized with an FITC label at the 5′-end of the sense strand. HEK 293 and SIRC cells were transfected with the labeled PF-655 reagent, and the proportion of cells that were positive for FTIC was determined by fluorescence microscopy and flow cytometry. The 2 complementary methods showed that a very high proportion of the cell line from both species took up PF-655 (Fig. 3A, B). Flow cytometry indicated that >90% of the HEK and SIRC cells were positive for FITC signal, indicating that both cell lines responded well to PF-655 transfection and potential differences in gene silencing effects between the 2 cell types could not be attributable to differences in transfection efficiency.
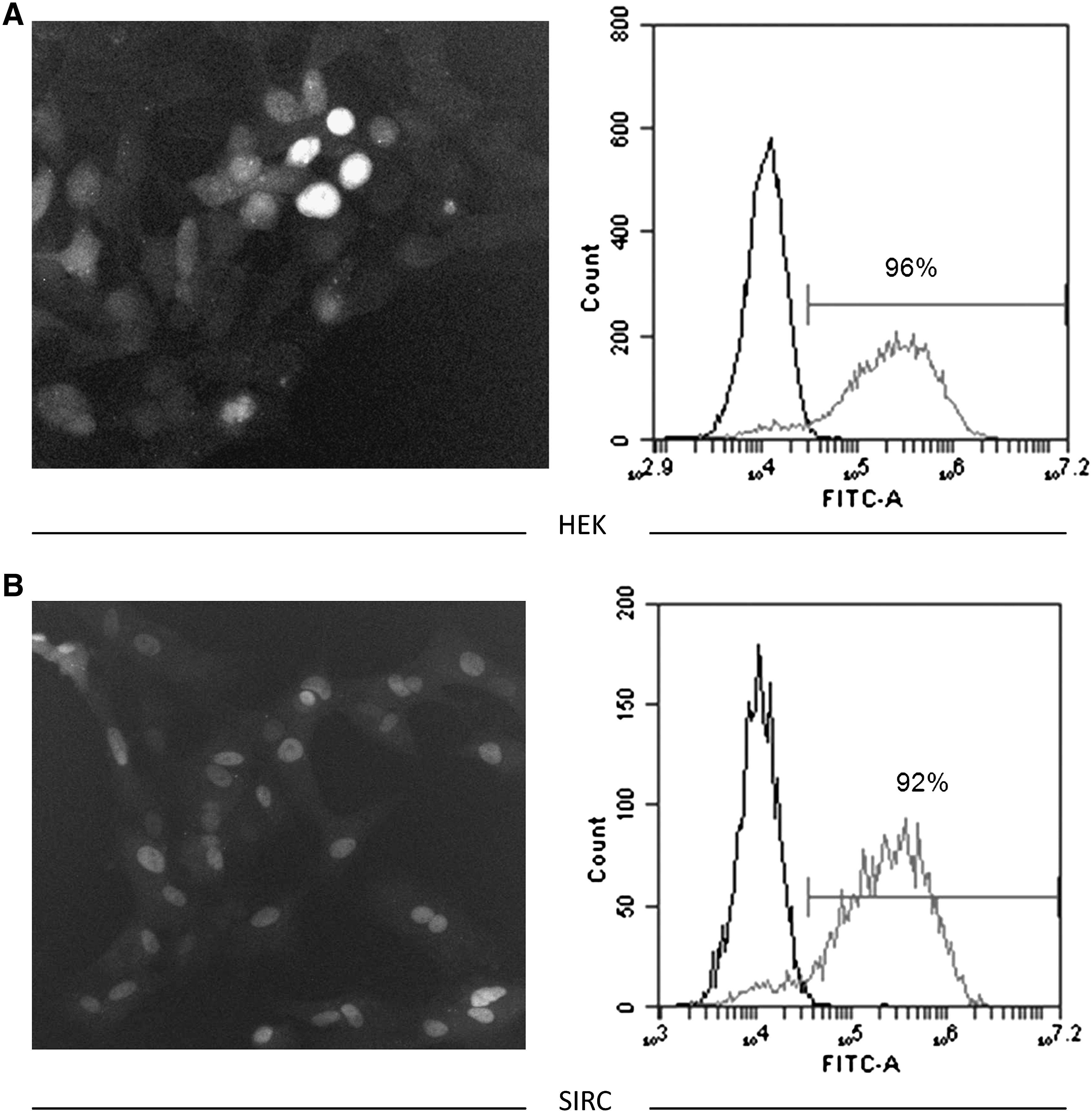
Human and rabbit cell lines exhibit similar levels of PF-655 uptake in vitro.
Next, we used PF-655 in a series of experiments to investigate the RTP801 gene silencing in both human and rabbit cells. HEK 293 cells were left untreated or transfected with either a nonspecific control siRNA duplex (NS) or PF-655 at 10 or 50 nM. Two days post transfection, the cells were acutely stimulated with CoCl2, and expression analysis of RTP801 was carried out at the protein and mRNA levels (Fig. 4A, B). As seen from the previous results of Fig. 2, CoCl2 stimulation induced protein expression of RTP801 and PF-655 was able to knockdown its expression at 50 nM, while the NS siRNA control had no significant effect. At the mRNA level, CoCl2 induced RTP801 expression 15–20-fold compared with the control, and PF-655 was able to decrease this expression to ∼25%–50% in a dose-dependent manner. Similar results on protein and mRNA expression were also observed with the SIRC cell line after PF-655 transfection (Fig. 4C, D). Compared with control and NS siRNA treated cells, PF-655 was able to decrease protein and mRNA expression to levels comparable with that observed in the HEK 293 cells. As an internal positive control for hypoxia induction with CoCl2 treatment, lysates from HEK cells were probed with an antibody for HIF-1α, a transcription factor that is upregulated under hypoxic stress. As seen in the immunoblot analysis (Fig. 4A, lower panel), HIF-1α is induced with CoCl2 treatment. Interestingly, it appears that PF-655 knockdown of the RTP801 gene further increased HIF-1α expression, indicating an important feedback loop with this transcription factor along with SP1 and their regulation of the RTP801 gene promoter. 18 The HIF-1α antibody used did not cross-react with the rabbit (data not shown).
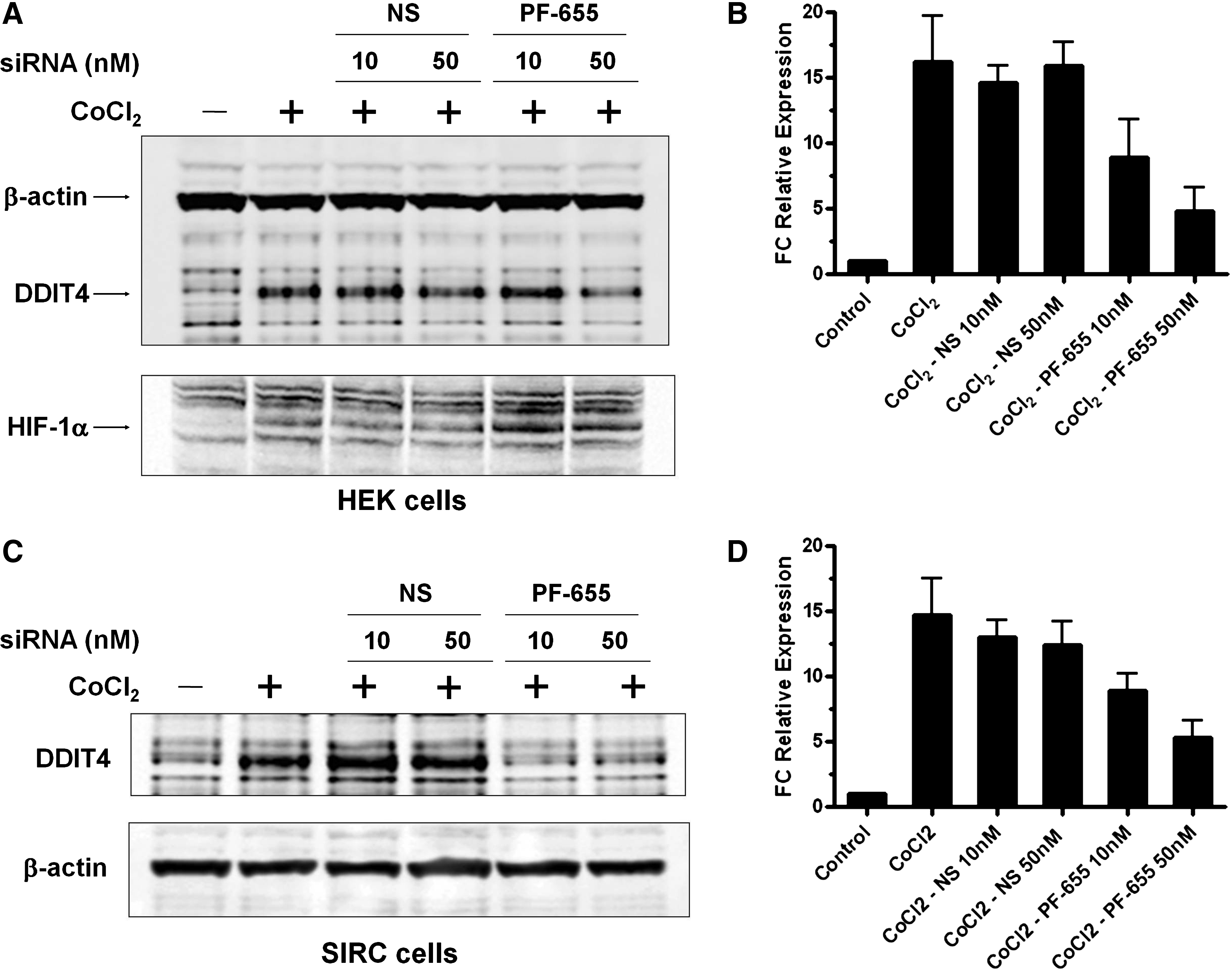
Induced expression of RTP801 in human and rabbit cell lines can be silenced by PF-655 at both the mRNA and protein levels.
Since an important downstream target of RTP801 is mTOR, which, in turn, regulates multiple components of the translational machinery and protein synthesis, we next investigated whether gene silencing of RTP801 with PF-655 had any adverse effects on cell viability. HEK and SIRC cells were left untreated or transfected with NS control siRNA or PF-655 at 25 or 100 nM. Then cell viability was assayed by examining total cellular ATP content at 24, 48, and 72 h post treatment (Fig. 5A, B). In HEK cells at 48 h, gene silencing at the lower 25 nM concentration of PF-655 caused cell viability to drop slightly to around 80%, compared with the 100% viability observed in the NS control 25 nM treated cells. At the higher siRNA concentration of 100 nM, both the NS and PF-655 treated cells exhibited decreased cell viability readings at 48 and 72 h (70%–85% cell viability relative to control cells), indicating the adverse effect on cell viability was not due specifically to RTP801 silencing (Fig. 5A). In the SIRC cell line, the only significant effect of PF-655 on cell viability was observed at 72 h in both the 25 and 100 nM treated cells (Fig. 5B). The NS control siRNA treated SIRC cells did not show any decrease in cell viability across the 2 siRNA concentrations and 3 time points.
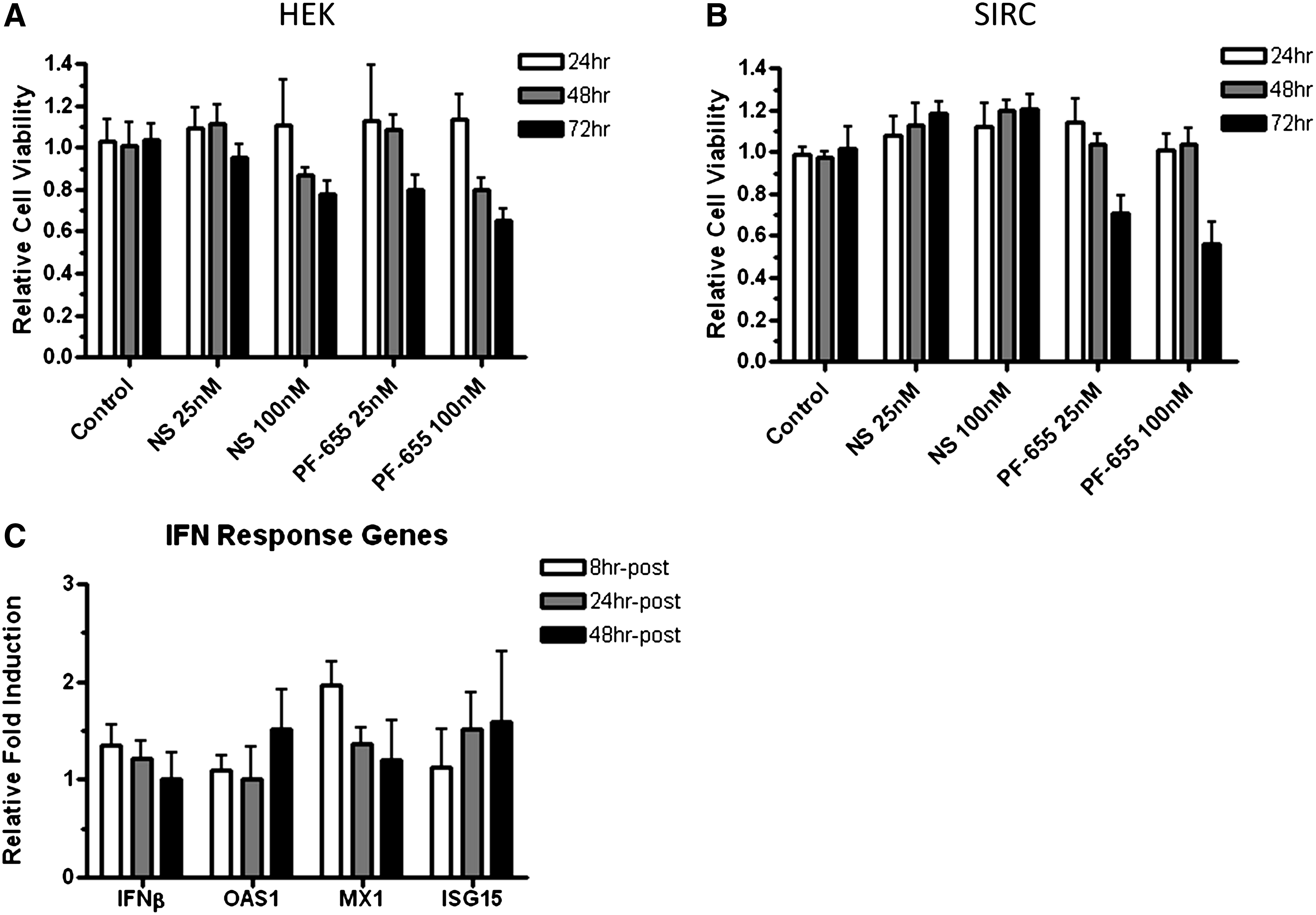
PF-655 exhibits effects on cell viability and induction of interferon (IFN) response genes.
We then examined the expression of genes involved in IFN responses during RTP801 silencing. HEK 293 cells were left untreated or transfected with 100 nM of PF-655. Induction of 4 IFN response genes (IFNβ, OAS1, MX1, and ISG15) were examined by q-PCR at 8, 24, and 48 h post treatment. Compared with untreated cells, increases in the expression of IFN response genes were observed, but the magnitude of changes were all less than 2.5-fold (Fig. 5C). The highest changes were about a 2-fold increase in MX1 expression at 8 h post-transfection and another 2-fold increase in ISG15 expression at 48 h post-transfection. The majority of the gene expression changes resulting from PF-655 treatment were 1.5-fold or less relative to control cells.
We next asked how long RTP801 silencing can last in vitro. HEK and SIRC cells were left untreated or transfected with PF-655 at 100 nM and q-PCR for the gene was assessed at 24, 48, 72, and 96 h post-treatment. Compared with the control, at 24 h, RTP801 gene levels were down to 75% and 63% in HEK and SIRC cells, respectively (Fig. 6). RTP801 silencing peaked at 48 h in both cell types, with levels down to ∼45% and 38% in HEK and SIRC, respectively. At 72 h post-treatment with PF-655, RTP801 gene expression reached levels to what were observed at 24 h in both cell types, and by 96 h, the silencing effect had largely diminished with mRNA levels reaching those observed in control cells.
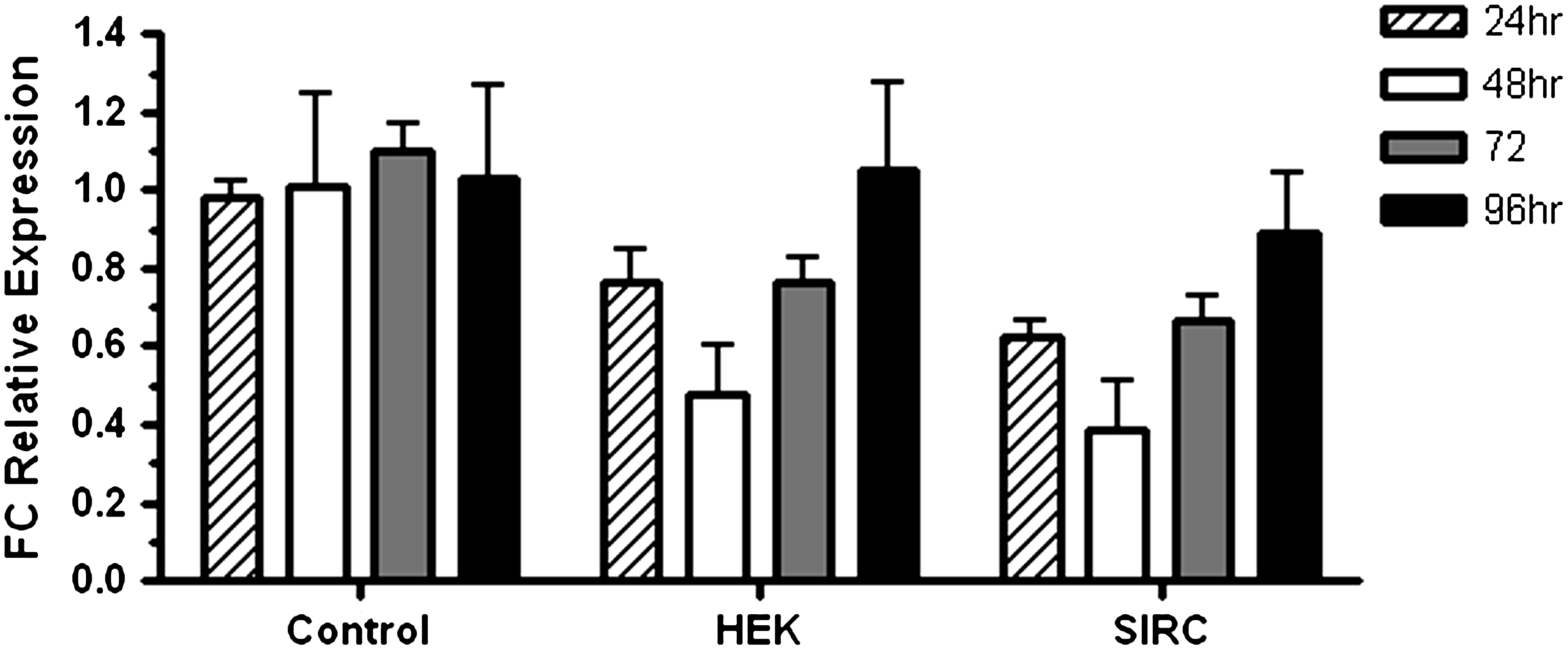
Time course of RTP801 silencing induced by PF-655 in human and rabbit cell lines. HEK 293 and SIRC cells were either mock transfected (control) or transfected with PF-655 at 100 nM. Real-time PCR was done for the RTP801 gene at the indicated time points. Data are normalized to β-actin and are indicated as FC relative to control cells. Standard deviation is shown by error bars, and the mean was calculated from 2 independent experiments.
Since the overall goal of our investigative studies with PF-655 was to help validate the Dutch-Belted rabbit species as an appropriate larger animal model for ocular toxicity and continual dosing studies with this therapeutic siRNA, we wanted to explore the expression level of RTP801 in primary retina tissues. In addition, we wanted to establish a confidence range for expression of this gene such that knockdown with PF-655 in vivo would not abolish detection with a q-PCR assay. Both left and right retinas were dissected from the Dutch-Belted rabbit and total RNA was prepared. Serial 4-fold dilutions were made, and reverse-transcriptase (RT) 24 reactions were done for each individual dilution. As a negative control, a no-enzyme (−RT) reaction with included with the undiluted RNA. q-PCR results for RTP801 show that that is no discernible difference in expression between left and right retina tissues (Fig. 7, left panel). In addition, there is a near perfect correlation between RNA dilution and cycle-threshold (Ct) values for the RTP801 gene, indicating a high efficiency for this assay. Results for the −RT sample showed a positive RTP801 Ct value of ∼33 (in both left and right retinas). This result indicates that Ct values near or above this mark have low confidence in detecting RTP801 expression in rabbit retina tissue, and a confident limit of detection in this assay is approximately down to 5%–10% of the expression level observed in the undiluted RNA sample. As an interesting comparison, a similar procedure was performed for β-actin in the Dutch-Belted retina. The results here indicate that the −RT control Ct value lies outside the positive detection range for this gene (Fig. 7, right panel). Most likely the difference seen between the 2 genes in this experiment is a result of the high mRNA copy number for β-actin compared with the lower levels observed for RTP801.

q-PCR analysis of RTP801 expression in rabbit retina tissues. Total RNA was isolated from right and left retinas of Dutch-belted rabbits. Four-fold serial dilutions of RNA were made, and reverse-transcriptase (RT) reactions were performed. A negative control (−RT enzyme) was included to determine the level of background signal in the q-PCR assay for RTP801 and β-actin. Cycle threshold (Ct) versus RNA dilution factor is graphed for each gene and the corresponding right or left eye. The horizontal line indicates the Ct value of the −RT control and indicates a positive limit of detection for the assay.
Discussion
Our data demonstrate that RTP801 expression can be induced under hypoxic conditions in rabbit cells, similar to what has been reported in human and rodent cell lines.18,19,25 To the best our knowledge, this is the first report of RTP801 induction and expression in a rabbit cell line. More importantly, our studies extend the utility of the Dutch-Belted rabbit as a model species for ocular siRNA therapeutic testing by validating gene silencing of the RTP801 gene with PF-655 in a rabbit cell line that has the same genetic sequence for the RTP801 region as the Dutch-Belted. Given the level of gene knockdown by PF-655 in rabbit cells with its existing single base sequence mis-match, our results further add to the complexity and promiscuity of the mechanism for siRNAs and how they recognize their target genes, and the importance of considering not only the type of nucleotide mis-match but also the position of the mis-match within the 19- to 21-mer sequence before deciding to validate a given siRNA in a different species. Previous studies have shown that a single mis-match between the siRNA guide strand and the target gene could be tolerated without abolishing gene silencing. 26 One report showed that both the position of the mis-match as well as the identity of the nucleotides forming the mis-match influence gene silencing. 27 In addition, purine-purine mis-matches disproportionately influence siRNA target discrimination compared with other nucleotide combinations. 28 We also examined RTP801 expression in primary retina tissue of the Dutch-Belted rabbit and generated a confidence interval for the detection of this gene by q-PCR with a bottom positive threshold level.
When comparing protein expression to that of mRNA in the HEK cell line experiment (Fig. 4A, B), we observed no discernible decrease in RTP801 protein when the cells were treated with CoCl2 and PF-655 at 10 nM, but did see a significant decrease in mRNA in the same experiment. On the other hand, in SIRC cells, CoCl2 and PF-655 at 10 nM exhibited RTP801 silencing as evidenced by decreases in both protein and mRNA (Fig. 4C, D). This discrepancy could be explained perhaps by a difference in mRNA translational efficiency for the RTP801 gene in HEK compared with SIRC cells such that one needs to bring down the mRNA to a critical mass in HEK cells to observe a gene silencing effect at the protein level. However, at the 10 nM concentration of PF-655, we did observe an increase in HIF-1α protein expression relative to untreated and NS siRNA treated cells, indicating that there was an effect on the downstream hypoxia pathway associated with a modulation of RTP801 expression.
Since all RNAs, including siRNA, have the potential to trigger IFN responses in cells, we also examined the induction of these genes to determine whether nonspecific responses to siRNA treatment were occurring in our experiments. 29 We did not observe the induction of IFN response genes at the more commonly seen magnitudes of 5–20-fold increases associated with siRNAs as indicated in published reports.24,30,31 The lack of a strong IFN response in HEK cells when treated with PF-655 is not surprising, because this therapeutic molecule was developed to minimize such nonspecific effects through novel chemical modifications of the siRNA. The induction or lack of a strong immunostimulatory response from an siRNA has been attributable to not only chemical modifications of the RNA duplex, but also the specific sequence of the siRNA itself.32,33
As more research continues into therapeutic siRNAs and greater numbers of targets are validated in vitro and in vivo using rodent models, there will become a growing need to use larger relevant animal species for regulatory toxicology studies before moving forward to the clinic, as well as supporting Phase III trials and registration. This is especially true for ocular disease where the small size of mice and rats severely limits the amount and exposure of drug that can be applied with intravitreal methods along with the increased possibility of trauma and irritation to the eye. 34 The rabbit species is amendable to long-term dosing and reproductive toxicity studies.20,35 With more genomic sequence coverage being added to online research databases (Broad Institute, Ensembl, UCSC), the rabbit can emerge as a relevant larger animal model for preclinical gene silencing studies. The future direction of this research study will be to move forward and intravitreally dose rabbits with PF-655 and determine the level of RTP801 gene knockdown in vivo.
Footnotes
Acknowledgments
The authors thank Walter Collette for isolation of retina tissue from the Dutch-Belted rabbit.
Author Disclosure Statement
The study was supported by Pfizer Global Research and Development, and all the authors are employees of the company.