
Abstract
Abstract
Purpose:
Dry eye is a multifactorial disease of tears and the ocular surface due to tear deficiency or excessive tear evaporation. Tear film instability is due to a disturbance in ocular surface mucin leading to a dysfunction of mucin, resulting in dry eye. In this study, we examined the effect of rebamipide, an anti-ulcer agent, on glycoconjugate production, as an indicator of mucin-like glycoprotein in cultured corneal epithelial cells. Further, we investigated the effect of rebamipide on the gene expression of membrane-associated mucins.
Methods:
Confluent cultured human corneal epithelial cells were incubated with rebamipide for 24 h. The glycoconjugate content in the supernatant and the cell extracts was measured by wheat germ agglutinin-enzyme-linked lectin assay combined gel-filtration method. In the experiment on mucin gene expression, cultured human corneal epithelial cells were collected at 0, 3, 6, and 12 h after administration of rebamipide. Real-time quantitative polymerase chain reaction was used to analyze the quantity of MUC1, MUC 4, and MUC16 gene expression.
Results:
Rebamipide significantly increased the glycoconjugate contents in the supernatant and cell extract. In the mucin gene expression in the cells, rebamipide increased MUC1 and MUC4 gene expression, but did not increase MUC16 gene expression.
Conclusions:
Rebamipide promoted glycoconjugate, which has a property as a mucin-like glycoprotein, in human corneal epithelial cells. The increased production was mediated by MUC1 and MUC4 gene expression.
Introduction
Mucins are highly O-glycosylated glycoproteins with a high molecular weight that are present on the apical surfaces of wet-surfaced epithelia including the ocular surface, respiratory, esophagus, and gastrointestinal. The roles of ocular surface mucin are maintaining tear film, lubricating the ocular surface during blinking, forming the barrier function of the ocular surface, removing pathogens, and so forth.1–3 On the ocular surface, mucins are classified as secreted type mucin that is supplied from conjunctival goblet cells, and membrane-associated type mucin appears on the surface of the cornea and conjunctival epithelium. 4 The stratified corneal and conjunctival epithelial cells express at least 3 membrane-associated mucins such as MUC1, MUC4, and MUC16.5–7 In dry eye patients, the mRNA of mucins was decreased compared with that of normal eyes. And the distribution of mucin was altered in dry eye patients compared with that of normal eyes.8,9
Rebamipide is an anti-ulcer agent; it indicates gastric ulcer and gastritis (Otsuka Pharmaceutical Co., Ltd.). 10 Rebamipide induces gastric endogenous prostaglandin E2, and promotes gastric epithelial mucin.11,12
Rebamipide eye drops for dry eye is under development to obtain indication in United States and Japan. Urashima et al. reported that rebamipide improved N-acetylcysteine-induced corneal and conjunctival injury in vivo, and the therapeutic effects are due to the increasing effects of corneal and conjunctival mucin-like substances. 13 In the site of mucin production by rebamipide, Rios et al. demonstrated that rebamipide accelerated the mucin production from cultured conjunctival goblet cells in vitro.14,15 However, the effect of rebamipide on the mucin production from corneal epithelial cells remains uncertain.
In this study, we aimed to evaluate the effect of rebamipide on glycoconjugate production by wheat germ agglutinin-enzyme-linked lectin assay (WGA-ELLA) combined gel-filtration method, as an indicator of produced mucin-like glycoprotein in corneal epithelial cells. We also investigated the effect of rebamipide on the gene expression of membrane-associated mucins (MUC1, 4, and 16) in corneal epithelial cells.
Methods
Materials
Rebamipide (Otsuka Pharmaceutical Co., Ltd.), 2-(4-chlorobenzoylamino)-3- [2(1H)-quinolinon-4yl-]-propionic acid, is a novel quinolinone derivative. EpiLife™ medium contained 5 μg/mL insulin, 0.18 μg/mL hydrocortisone, 5 μg/mL transferrin, 1 ng/mL mEGF, 0.4% bovine pituitary extract, 25 ng/mL amphotericin B, 10 U/mL penicillin, 100 μg/mL streptomycin, and 0.06 mM Ca2+ were obtained from Kurabo Industries Ltd. Bovine serum albumin and protease inhibitor cocktail were obtained from Sigma-Aldrich. Bovine submaxillary gland mucin (BSM) was obtained from Wako Pure Chemical Industries. Horse radish peroxidase conjugated WGA (HRP-WGA) was obtained from HONEN.
Cell culture of human corneal epithelial cells
Human corneal epithelial cells were purchased from Kurabo Industries Ltd. These human corneal epithelial cells were primary cultured cells isolated from human corneas, not immortalized cells. Cells were cultured in collagen-coated plate with EpiLife™ medium until sub-confluent at 37°C in a 5% CO2 incubator.
Test compound treatment and sample recovery for glycoconjugate content measurement
After human corneal epithelial cells reached confluence, the medium was replaced with a medium supplemented with or without rebamipide.
In the time-course study, after 12, 24, 48, and 72-h incubation, the culture supernatant was recovered and a protease inhibitor cocktail was added. In the dose-dependent study, after 24-h incubation, the culture supernatant was recovered and a protease inhibitor cocktail was added. After removing the culture supernatant, an extraction buffer [1% SDS, 10% protease inhibitor cocktail in D-phosphate-buffered saline (PBS)] was added to the remaining cells. After the cells were dissolved, the cell extract was collected and stored overnight. The extracts were centrifuged (15,000 rpm 10 min at room temperature) to remove cell debris.
Gel filtration for glycoconjugate content measurement
Gel filtration was performed to obtain a high molecule weight fraction from samples. Econo-column (Inner diameter 0.7 cm and height 50 cm; BIO-RAD) and sepharose CL-4B (Amersham Pharmacia Biotech) were used for gel filtration. The culture supernatant and cell extract (0.6 mL) were applied to the column. Samples of culture supernatant and cell extract were recovered from void volume (very high molecule weight fraction not isolated by gel) fraction (culture supernatant; 5.2–7.6 mL, cell extract; 5.2–8.4 mL).
Measurement of the amounts of glycoconjugate using the WGA-ELLA method
After the samples and standard (BSM) were added, the 96-well plates were stored overnight in a freeze dryer. After washing with ice-cold PBS, 1.0% bovine serum albumin was added and the plates were kept for 1 h at 4°C. After washing, 1.0 μg/mL HRP-WGA was added and the plates were kept for 1 h at 37°C. The WGA bonding amount was measured by the absorbance of 405 nm. The WGA bonding amount was calculated by the standard curve of BSM.
Test compound treatment and DNA extraction
After human corneal epithelial cells reach confluence, the medium was replaced with a medium supplemented with or without 10−5 M rebamipide. After 0, 3, 6, and 12-h incubation, RNA extract agent (ISOGEN®, Nippon gene) was added after removing the culture supernatant. After that, total RNA from human corneal epithelial cells was extracted based on the instructions of the procedural manual included with ISOGEN®. The quality of total RNA was confirmed by checking of the 28S:18S rate.
The complementary DNA was synthesized using total RNA and Oligo (dT) 20 primer. A 5 μg portion of total RNA was used for reverse transcription with SuperScript™ III First-Strand Synthesis System (Invitrogen™ life technologies).
Gene quantity analysis
A real-time quantitative polymerase chain reaction (PCR) (iCycler™; Japan Bio-rad Laboratories, Inc.) was used to analyze the quantity of gene expression. The genes of MUC1, MUC4, and MUC16 were analyzed. The housekeeping gene β-actin was used as an internal control. Each primer is shown in Table 1.5,16,17
Diluted samples of reverse-transcribed DNA were amplified using PCR with iQ™ SYBR® Green Supermix and the PCR was performed under the following conditions: β-actin was used to control RNA quality. The amplification reactions of all targets were performed in iCycler iQ. The conditions consisted of an initial denaturation for 3 min at 95°C followed by 45 cycles (40 cycles for β-actin) of denaturation for 30 s at 95°C, amplification for 30 s at 57°C, and extension for 30 s at 72°C. A final extension was performed for 10 min at 72°C. The threshold cycle value with determined parameters was calculated using real-time PCR analysis software (iCycler; Bio-Rad Laboratories, Inc.). Measured levels of MUC1, MUC4, and MUC16 mRNA were divided by the level of human β-actin mRNA in each group.
The amplified PCR products were analyzed by electrophoresis on agarose gel.
Statistical analysis
In the time-course study for glycoconjugate content measurement, the differences between control and rebamipide were analyzed by 2-tailed t-test.
In the dose-dependent study, linear regression analysis confirmed that the glycoconjugate contents in the supernatant showed monotonous increase with the logarithmic concentration of rebamipide; it was followed by upper-tailed Williams test.
Linear regression analysis also confirmed that the glycoconjugate contents in the cell extracts did not show monotonous increase with the logarithmic concentration of rebamipide; it was followed by 2-tailed Dunnett's test.
A P-value of less than 0.05 was considered statistically significant. Statistical analysis was performed using SAS software (Release 8.1, SAS Institute Japan).
Results
Time course of rebamipide-induced glycoconjugate production in human corneal epithelial cells
The time course of glycoconjugate production in the supernatant after treatment of 10−5 M rebamipide in human corneal epithelial cells was examined. The time course of glycoconjugate production is shown in Fig. 1.
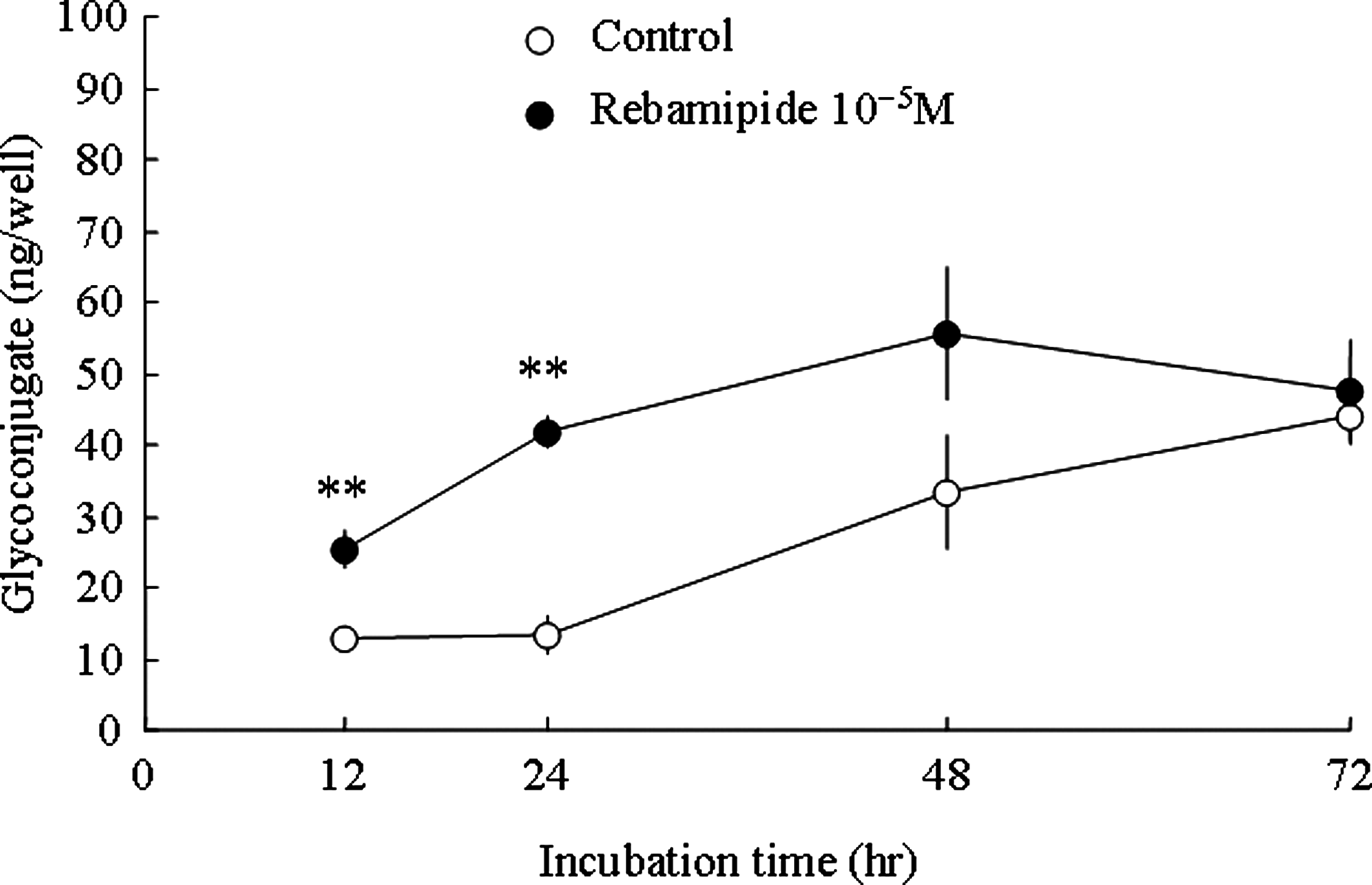
Time course of rebamipide-induced glycoconjugate production in human corneal epithelial cells. Twelve, 12, 24, 48, and 72 h after administration of 10−5 M rebamipide. The supernatant was recovered to measure the glycoconjugate content. All values were expressed as mean±S.E (n=3–4). Statistical analysis was performed by t-test (2-tailed). **P<0.01 versus control.
Glycoconjugate significantly increased from control groups at 12 or 24 h after treatment of rebamipide. At 48 or 72 h after treatment, it showed no significant difference between rebamipide and control group. The maximal difference between both groups was shown at 24 h after treatment.
Effects of rebamipide on the glycoconjugate in human corneal epithelial cells
The effects of rebamipide on glycoconjugate production in human corneal epithelial cells were examined. The glycoconjugate contents in the culture supernatant and cell extract are shown in Fig. 2.
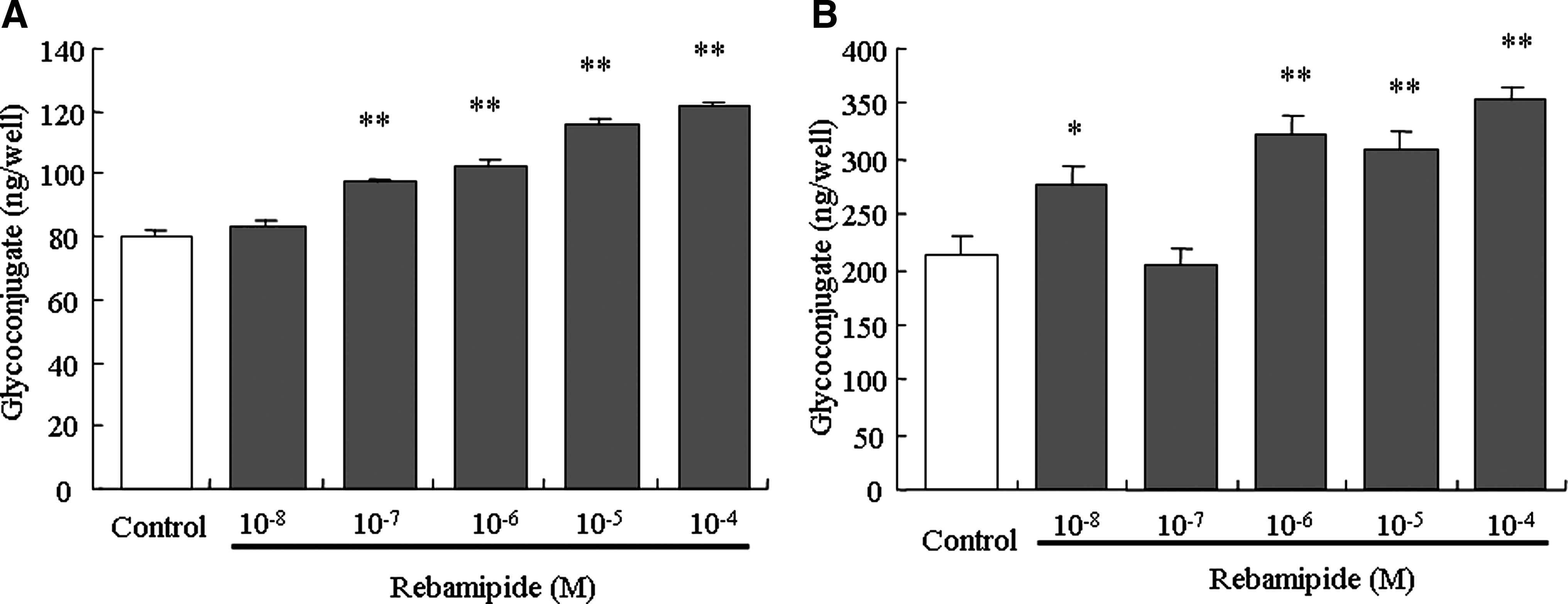
Effects of rebamipide on the glycoconjugate contents in human corneal epithelial cells. (
In the supernatant, rebamipide dose-dependently increased the glycoconjugate contents. Rebamipide at concentrations of 10−7 M or higher showed a significant increase in glycoconjugate.
In the cell extract, rebamipide increased the glycoconjugate contents. Rebamipide at concentrations of 10−8 M and 10−6 M or higher showed a significant increase in the glycoconjugate.
Effect of rebamipide on expression of mucin genes in human corneal epithelial cells
Gene expression of MUC1, MUC4, and MUC16 after treatment of 10−5 M rebamipide in human corneal epithelial cells was examined. MUC1 and MUC4 mucin gene expression are shown in Table 2. In MUC1 gene expression, rebamipide increased to 1.10, 1.91, 1.86, and 1.20 (the ratio of the mucin to β-actin) at 0, 3, 6, and 12-h incubation, respectively. MUC1 gene expression increased at 3 or 6 h after treatment; after that decreased to the constitutive level.
The medium was replaced with a medium supplemented with or without 10−5 M rebamipide.
After 0, 3, 6, and 12-h incubation, samples were recovered for the gene expression.
The results were shown as the ratio of the quantity of gene expression in the β-actin group and each mucin gene (MUC1, MUC4).
All values were expressed as mean±S.E (n=4).
In MUC4 gene expression, rebamipide increased to 1.05, 1.09, 1.45, and 1.17 at 0, 3, 6, and 12-h incubation, respectively. MUC4 gene expression increased at 6 h after treatment; after that decreased to the constitutive level.
Rebamipide did not increase MUC16 gene expression (data not shown).
In the electrophoresis, it was confirmed that the amplified PCR products of MUC1, 4 and, 16 showed a single band, which located the theoretically-based molecular weights.
Discussion
We evaluated the effect of rebamipide on glycoconjugate production as an indicator of mucin-like glycoprotein in the cultured corneal epithelial cells. We observed an increased glycoconjugate by the administration of rebamipide.
In this study, glycoconjugate productions were measured by WGA-ELLA combined gel-filtration method. WGA specifically binds to N-acetylglucosamine, 18 which is a specific component of the mucin. Further, a number of histochemical studies have demonstrated WGA binding to the ocular surface glycocalyx, in which membrane-associated type mucin is located.19,20 In the characterization of the ocular surface material in dogs, Hick et al. reported that the extracted materials in void volume by gel filtration showed a positive peak for anti-mucin antibody. 21 Therefore, the high molecular weight glycoconjugate measured by this method has a property as a mucin-like glycoprotein.
Rebamipide increased glycoconjugate in the cell extract and the culture supernatant. The membrane-associated type mucin has the plasmotomy part in the outside of the cell. It is released in soluble form by plasmotomy, and it has been shown to be released from corneal epithelial cells.22,23 Increasing mucin-like glycoprotein in the cell extract showed an increase in the membrane-associated type mucin. On the other hand, increasing mucin-like glycoprotein in the culture supernatant showed an increase in mucin released from the membrane surface.
It has been reported that some agents promote mucin-like glycoprotein production in conjunctival goblet or corneal epithelial cells. Gefarnate, a therapeutic agent for gastric ulcer and gastritis as well as rebamipide, increases PAS positive cells in rabbit conjunctive. 24 DA-6034, a potent secretagogue, increased conjunctival goblet cell number in dry eye mouse model. 25 P2Y2 agonist INS 365 secreted mucin-like glycoprotein from conjunctival goblet cells. 26 15-(S)-HETE, stimulate secretion of mucin-like glycoprotein in rabbit corneal epithelium. 27 However, no agent acts both on corneal epithelium and conjunctival goblet cells. Rebamipide promoted the conjunctival secreted type mucin in the previous study, 15 and promoted the corneal membrane-associated type mucin in this study. Therefore, rebamipide is a new type of agent that acts both on corneal epithelium and conjunctival goblet cells.
We investigated the gene expression of 3 membrane-associated mucins: MUC1, MUC4, and MUC16 in this study. Rebamipide upregulated the expression of MUC1 and MUC4 genes.
As for MUC1, a role as non-adhesion molecules has been reported. 28 Additionally, MUC1 plays a role in defending against bacterial infection. Fleiszig et al. reported that adhesion for the Pseudomonas aeruginosa was elevated by decreasing the MUC1 level. 2 MUC4 also plays a role in the maintenance of tear fluid stability. 29 Further similar to MUC1, MUC4 acts as an anti-adhesive molecule, preventing integrin-mediated cell adhesion in breast carcinoma cells. 30 Thus, the elevated formations of MUC1 and MUC4 might contribute to the prevention of infection by bacteria and the lubricating action of the ocular surface, and rebamipide might promote such actions.
Rebamipide did not activate the expression of MUC16 in our present study. In the next study, we should investigate under other conditions such as changing the experimental method and/or measurement timing for MUCs.
In the mechanism of rebamipide-induced mucin-like glycoprotein production, we speculated a prostaglandin E2 production, EP4 receptor, and EGF receptor activation. Suetsugu et al. reported that rebamipide promoted gastric prostaglandin E2 production and mucus secretion, and these effects are seen to be mediated by EP4. 31 Rios et al. reported that the secreted mucin production by rebamipide in cultured conjunctival goblet cells was related to activation of EGF receptor and MAP kinase. 15 However, the mechanism of increasing the membrane-associated mucin in corneal epithelial cells by rebamipide remains uncertain and requires future study.
In conclusion, rebamipide induced glycoconjugate, indicating a mucin-like glycoprotein property, in corneal epithelial cells. Additionally, rebamipide induced MUC1 and MUC4 gene expression. These data demonstrate that rebamipide accelerated not only conjunctival secreted type mucin, but also corneal mucin-like glycoprotein, and this suggests that rebamipide has an effect on the barrier function and the lubricating action of the ocular surface during blinking, in which the membrane-associated mucin plays an important role.
Footnotes
Author Disclosure Statement
All authors are employees of Otsuka Pharmaceutical Co, Ltd.