
Abstract
Abstract
Purpose:
Pathological retinal angiogenesis is a major cause of vision loss. Endostatin is a natural antiangiogenesis antitumor protein that is widely used in cancer studies. In this study, we investigated the efficacy and potential mechanisms of endostatin for the prevention of retinal neovascularization both in vitro and in vivo.
Methods:
Human umbilical vein endothelial cells (HUVECs) were used for the in vitro studies. HUVECs were incubated with endostatin or the vascular endothelial growth factor (VEGF) and endostatin for different time points. Cell proliferation, migration, cell cycling, and tube formation studies were carried out using a Cell Counting Kit-8 assay, a Transwell assay, flow cytometry, and a Matrigel assay, respectively. Enzyme-Linked Immunosorbent Assay (ELISA) was used to study VEGF and pigment epithelial-derived factor (PEDF) protein secretion from the HUVECs at different time points. A murine oxygen-induced retinopathy (OIR) model was used for the in vivo studies. Seven-day-old C57BL/6J pups (p7) were exposed to 75% oxygen for 5 days. On p12, the animals were returned to a normal atmosphere and were immediately injected intravitreously with 1.5 μL of a 5 mg/mL endostatin solution. At p18, the mice were perfused with fluorescein-dextran-FITC, and their retinas were flat mounted to measure the nonperfused area. Retinal VEGF and PEDF levels were also measured by ELISA Kits in the OIR mice at p18.
Results:
In vitro, endostatin inhibited HUVEC proliferation in a dose-dependent manner and also inhibited HUVEC proliferation in a VEGF-containing medium. Additionally, endostatin can inhibit migration, tube formation, and VEGF secretion in HUVECs, while also inducing apoptosis in HUVECs at several time points. These effects were statistically significant when compared to the control group (P<0.05). In vivo, a single intravitreous injection of endostatin reduced the retinal nonperfused area from 30% in the control group to 23% in the treatment group (P<0.0001). Intravitrous injection of endostatin reduced VEGF levels in retinas, while it increased PEDF levels.
Conclusions:
Endostatin showed convincing inhibitory effects on angiogenesis both in vitro and in vivo. The inhibitory effects may be, at least partly, resulted from the restoration of the PEDF/VEGF ratio. These data suggest that endostatin could offer an innovative pharmaceutical strategy for the prevention of retinal neovascularization.
Introduction
Endostatin was first discovered in the laboratory of Judah Folkman in 1997 and is a naturally occurring 20-kDa C-terminal fragment derived from type XVIII collagen, which is a member of the multiplexin family. 8 Since the discovery of endostatin, it has been reported to be an endogenous inhibitor of angiogenesis, similar to angiostatin and thrombospondin.9,10 Due to its potent antiangiogenesis effects, endostatin is considered to have great potential as an anticancer drug in cases of malignancies that involve ephrin, TNF-α, or NFκB.11,12 Despite there are numerous studies of cancer treatments, the comprehensive studies of endostatin in retina neovascularization are rare. In previously published articles, overexpressed endostatin showed inhibition effects on choroidal neovascularization. 13 Most recently, RetinoStat®, an equine infectious anemia virus-based lentiviral gene therapy vector that expresses the angiostatic proteins endostatin and angiostatin, also displayed well-tolerated, localized characteristics, and capable of persistent expression after subretinal delivery.14,15 All of these results give us hopes for the use of endostatin in retinal neovascularization diseases.
In this study, we extensively investigated the antiangiogenic effects and possible mechanisms of action of endostatin both in vitro and in vivo. We investigated the ability of endostatin to inhibit angiogenesis in human umbilical vein endothelial cells (HUVECs) and to prevent retinal neovascularization in murine oxygen-induced retinopathy (OIR) models. We also probed the secretion of VEGF and pigment epithelial-derived factor (PEDF) in the retina after treatment with endostatin with the goal of analyzing possible mechanisms of action. The encouraging results of our study provide an innovative treatment strategy for retinal neovascularization.
Methods
Cells and animals
HUVECs were obtained from the American Tissue Culture Collection (ATCC, Manassas, VA) and preserved in our laboratory. 16 HUVECs were cultured in Dulbecco's Modified Eagle's Media (DMEM) with 10% fetal bovine serum (FBS; Hyclone) in a humidified 37°C incubator containing a 5% CO2 atmosphere. Neonatal mice (C57BL/6J) were obtained from the Peking University animal center and were raised in the animal room of Peking University People's Hospital. This study adhered to the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research and was performed in accordance with the guidelines provided by the Animal Care Use Committee of Peking University. The animals were housed with free access to laboratory food and water and kept on a 12-h light–12-h dark cycle.
Proliferation assays
A Cell Counting Kit-8 (CCK-8; Dojindo) assay was used to evaluate the effects of endostatin (SIGMA; Cat# E8154) on angiogenesis as previously described. 17 Briefly, HUVECs were synchronized in DMEM at a density of 1×104 cells per well in 96-well plates overnight without FBS. Because there is no consistent use dosage of endostatin in published articles for HUVECs, and according to the dosage range of our previously published article, which involved another endogenous antiangiogenesis agent, 17 endostatin (10−2–10−7 mg/mL) was diluted in a series of 1:10 dilutions and incubated with cells for 24, 48, and 72 h. After adding 10 μL CCK-8 to each well, the cells were incubated at 37°C for another 30–60 min. Absorbance was measured with an Enzyme-Linked Immunosorbent Assay (ELISA) plate reader at 450 nm. Each experiment was repeated in 5 wells and was duplicated at least 3 times. To evaluate the antiangiogenic effects of endostatin under the stimulation of VEGF, a 20 ng/mL solution of VEGF was added to the culture medium in another experimental group. Endostatin (10−2–10−7 mg/mL) was incubated for 24, 48, and 72 h, and the CCK-8 assays were performed as stated before.
Migration assay
HUVEC migration was assayed using a Transwell (Corning; Cat# 3422) with the pore size of 8.0 μm, as previously described. 17 Briefly, 2×104 cells were placed in the top part of a transwell in 200 μL of a serum-free medium. DMEM (containing 10% FBS) with 10−2 , 10−4 , or 10−6 mg/mL endostatin was placed in the bottom chamber, for a final volume of 600 μL. All migration assays were conducted at 37°C for 5 h, and then the cells were fixed with 4% paraformaldehyde (PFA) for 30 min and stained with 4,6-diamino-2-phenyl indole (DAPI; Roche, 10236276001) for 15 min. The cells that had not migrated were removed with a cotton swab, and the membrane was imaged by fluorescence microscopy (Zeiss Axiophot). Cells from 5 random fields of view were counted, and the average was used for statistical analysis.
ELISA for VEGF and PEDF measurement in HUVECs
HUVECs were seeded in 96-well plates (1×104 per well) and incubated at 37°C overnight. A culture medium containing endostatin (10−2–10−7 mg/mL) was added to the wells after removing DMEM. After 24, 48, or 72 h of incubation, the cell culture supernatant was harvested and centrifuged. The VEGF and PEDF protein secreted by HUVECs in the culture medium was measured using an ELISA Kit (Bostar, EK0575 for VEGF; Bostar, EK0896 for PEDF) according to the manufacturer's instructions.
Flow cytometry analysis of HUVEC apoptosis
HUVEC apoptosis was measured with a FITC Annexin V Apoptosis Detection Kit (BD Science; Cat# 556547). Briefly, HUVECs (1×106) were seeded in 6-well plates and incubated for 24, 48, or 72 h with 10−2 , 10−4 , or 10−6 mg/mL endostatin or controls. Then, the cells were stained with Annexin-V-FITC and propidium iodide (PI), according to the manufacturer's instructions. Flow cytometry analysis was immediately performed (ex/em=488/530 nm). The samples were analyzed by a flow cytometer (FACSCalibur; BD Biosciences) with Cell Quest software (BD Biosciences). About 104 cells were collected and divided into 4 groups: dead cells (Annexin V−/PI+, UL), late apoptotic cells [Annexin V+/PI+, upper right (UR)], viable cells (Annexin V−/PI−, LL), and early apoptotic cells [Annexin V+/PI−, lower right (LR)]. The apoptotic rate was calculated as the percentage of early apoptotic cells (LR) plus late apoptotic cells (UR).
Tube formation study
Tube formation studies are used to test the ability of endothelial cells to form capillary-like structures. According to the manufacturer's instructions and our previous report, 18 150 μL of a Matrigel (BD Sciences; Cat#354234) solution was placed into the wells of 48-well plates, and then incubated at 37°C for 30 min in a 5% CO2 incubator. HUVECs (5×104 per well) treated with 10−2 , 10−4 , or 10−6 mg/mL endostatin or controls, were seeded on the Matrigel and cultured in DMEM for 8–10 h. The networks in Matrigel from 5 randomly chosen fields were counted and photographed. The length of the tubes was measured using ImageJ software. Each experiment was repeated 3 times.
Induction of OIR mouse model and assessment of nonperfusion area
C57BL/6J pups were exposed to hyperoxia (75% oxygen) for 5 days beginning on postnatal day 7 (p7). On p12, the animals were brought back to normoxia until day p17. 17 At p12, the OIR mice were injected intravitreally with endostatin or IgG. Both of the drugs were injected in a 1.5 μL and at 5 μg/μL concentration. At p18, the mice were deeply anesthetized intraperitoneally with chloral hydrate (0.2 mL/10 g body weight), and then perfused with 0.5 mL of PBS containing 50 mg of 2×106 molecular weight fluorescein-dextran-FITC (Sigma). The eyes were removed and fixed in 4% PFA for 30 min, and the retinas were flat mounted using 4 peripheral retinal cuts. The retinas were photographed during fluorescence microscopy. The nonperfused areas were analyzed using ImageJ software. The ratio was determined by comparing the nonperfused area to the whole retina. All of the data were analyzed by 2 individuals, one of whom was completely blind to the study groups, and the average was used for statistical analyses.
Retinal VEGF and PEDF concentration measurement
Five days after injection with endostatin, C57BL/6J pups were sacrificed on day p18, and the retinas were separated for VEGF and PEDF detection. The concentration of VEGF and PEDF (pg/mL protein) in the clarified supernatant was measured using an ELISA Kit (Bostar, EK0541 for VEGF; Hua Yi Bio-technology, HE2453 for PEDF). All of the experiments involving the ELISA tests were performed in 5 pups, and each experiment was repeated 3 times.
Statistical analysis
Data analysis was performed using the statistical software Prism 5 (GraphPad Software, Inc.). All data are presented as the mean±SEM. Differences were evaluated with ANOVA, followed by the Student–Newman–Keuls test for multiple comparisons and the Student's t-test for pairwise comparisons. In the case of ELISA results, the data were analyzed using a 2-way ANOVA, followed by Bonferroni post-tests for comparisons among the 2 groups. A P<0.05 was considered to be a statistically significant difference.
Results
Endostatin inhibits HUVEC proliferation
A proliferation study was used to evaluate the antiangiogenic effects of endostatin in vitro. HUVECs were incubated for 24, 48, or 72 h with various concentrations of endostatin (10−2–10−7 mg/mL). As shown in Fig. 1 and Table 1, endostatin inhibits HUVEC proliferation in the general culture medium until 48 h at 10−2–10−6 mg/mL. This result is significantly different from the control group (Fig. 1A, B, P<0.05). However, at the 72-h time point, endostatin did not show an inhibitory effect (Fig. 1C). To further evaluate the inhibitory effect of endostatin on VEGF stimulation, 20 ng/mL of VEGF was added to the cell culture medium. Figure 1D–F and Table 2 show that endostatin exhibited an inhibitory effect on HUVEC proliferation until 72 h at concentrations of 10−2–10−5 mg/mL (P<0.05).
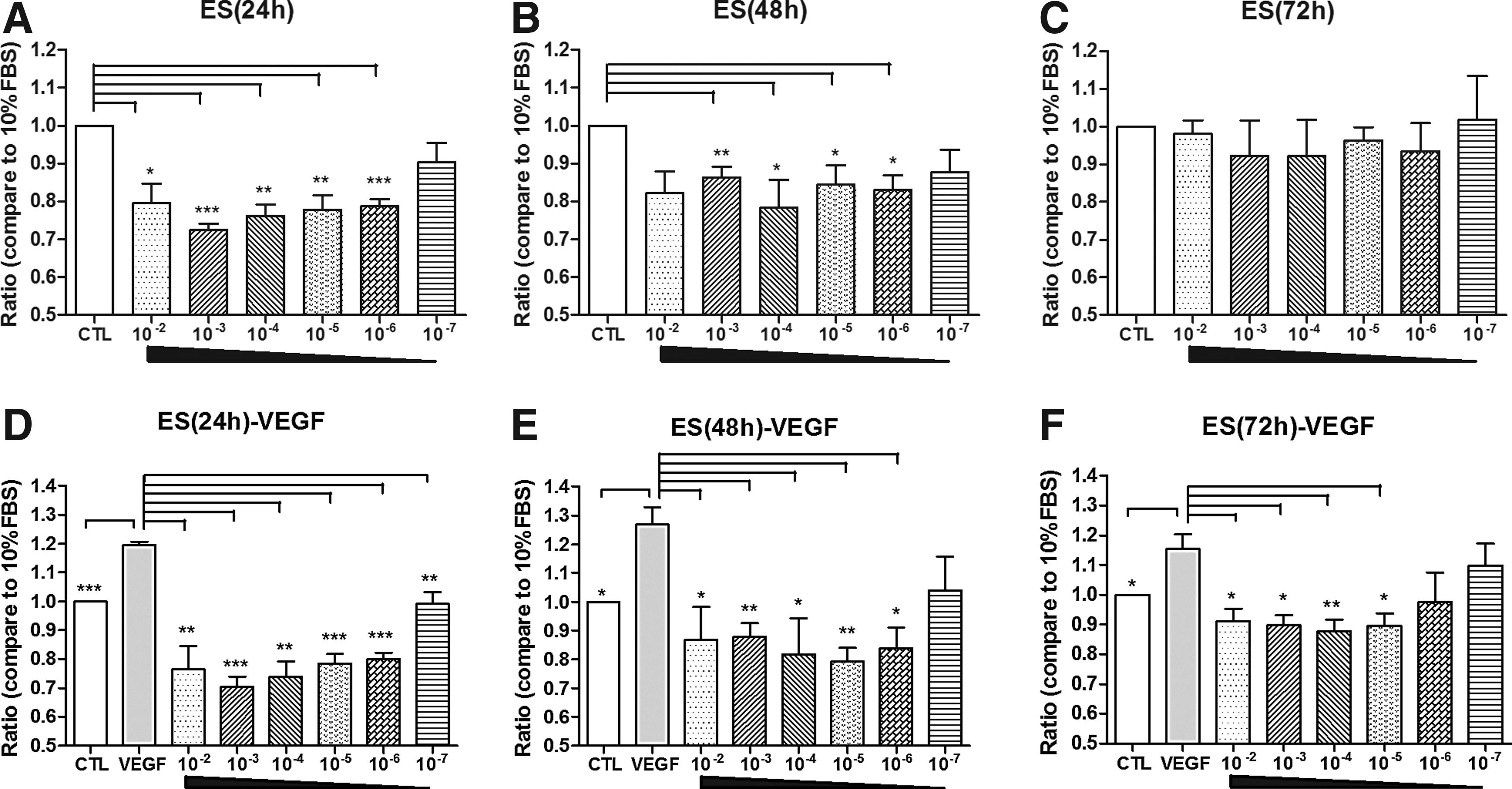
Effects of endostatin on the proliferation of human umbilical vein endothelial cells (HUVECs). HUVEC proliferation was measured with a CCK-8 assay at 24, 48, and 72 h.
FBS, fetal bovine serum.
VEGF, vascular endothelial growth factor.
Endostatin inhibits HUVEC migration
The migration study involved a transwell assay. As shown in Fig. 2, the number of cells that passed through the membrane in the endostatin-treated HUVEC groups (at both 10−2 and 10−4 mg/mL) was significantly lower than in the number in the control groups (Fig. 2A, B, D, F, P<0.05). At concentrations of 10−2 and 10−4 mg/mL, endostatin also inhibited the mobility of the VEGF-stimulated group (Fig. 2C, E, F, P<0.05).
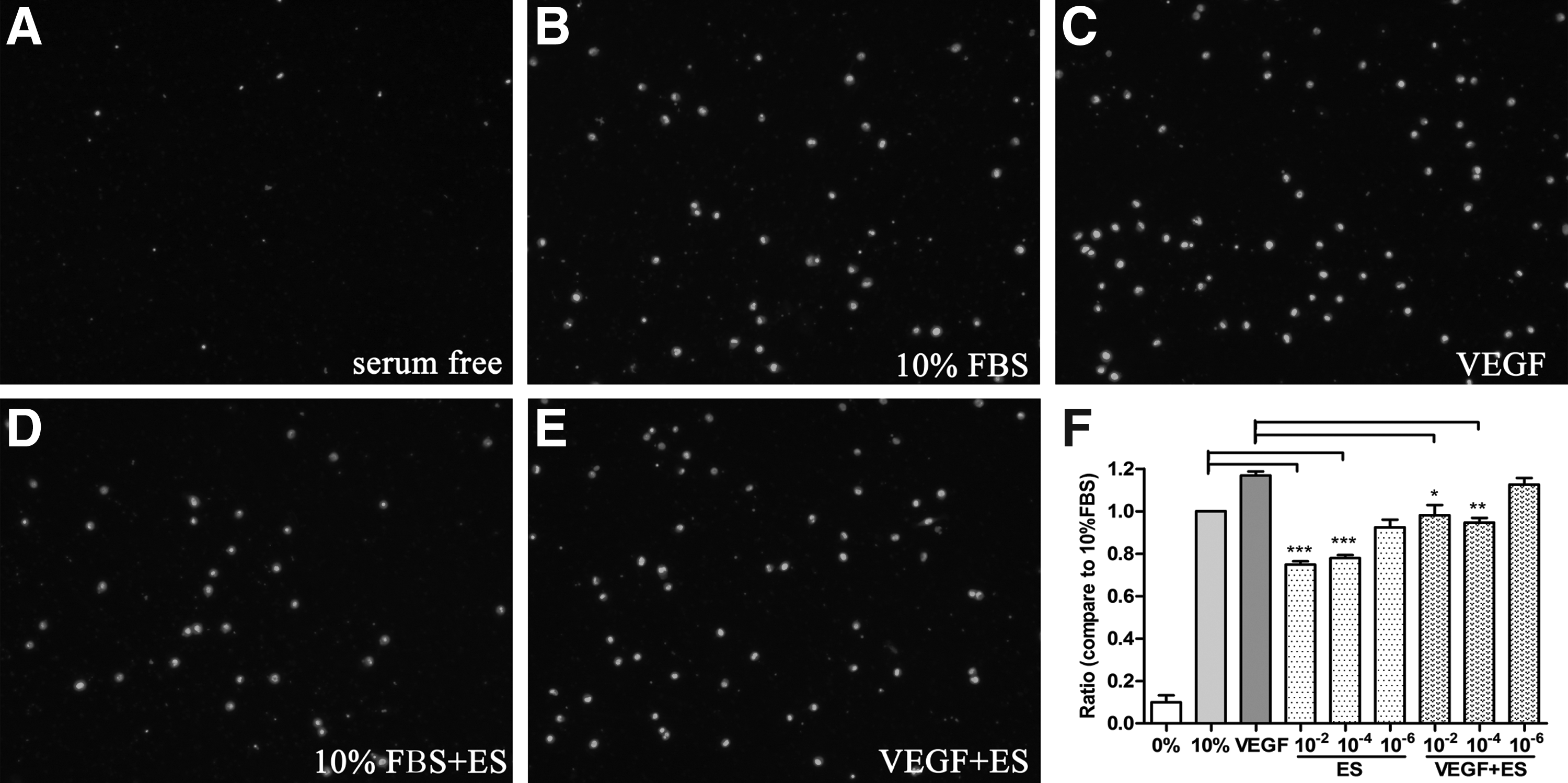
Effects of endostatin on the migration of HUVECs. Migration study was evaluated using a transwell assay. Results
Endostatin induces HUVEC apoptosis
FACS was used to evaluate early and late apoptosis effects. As shown in Fig. 3 and Table 3, after incubation with endostatin for 24, 48, or 72 h, the early and late apoptotic HUVECs showed significant differences from the controls in the 10−2 and 10−4 mg/mL treated groups at the 24- and 48-h time points. The percentage of apoptotic cells (UR+LR%) in these treated groups was significantly higher than in the control group (P<0.05).
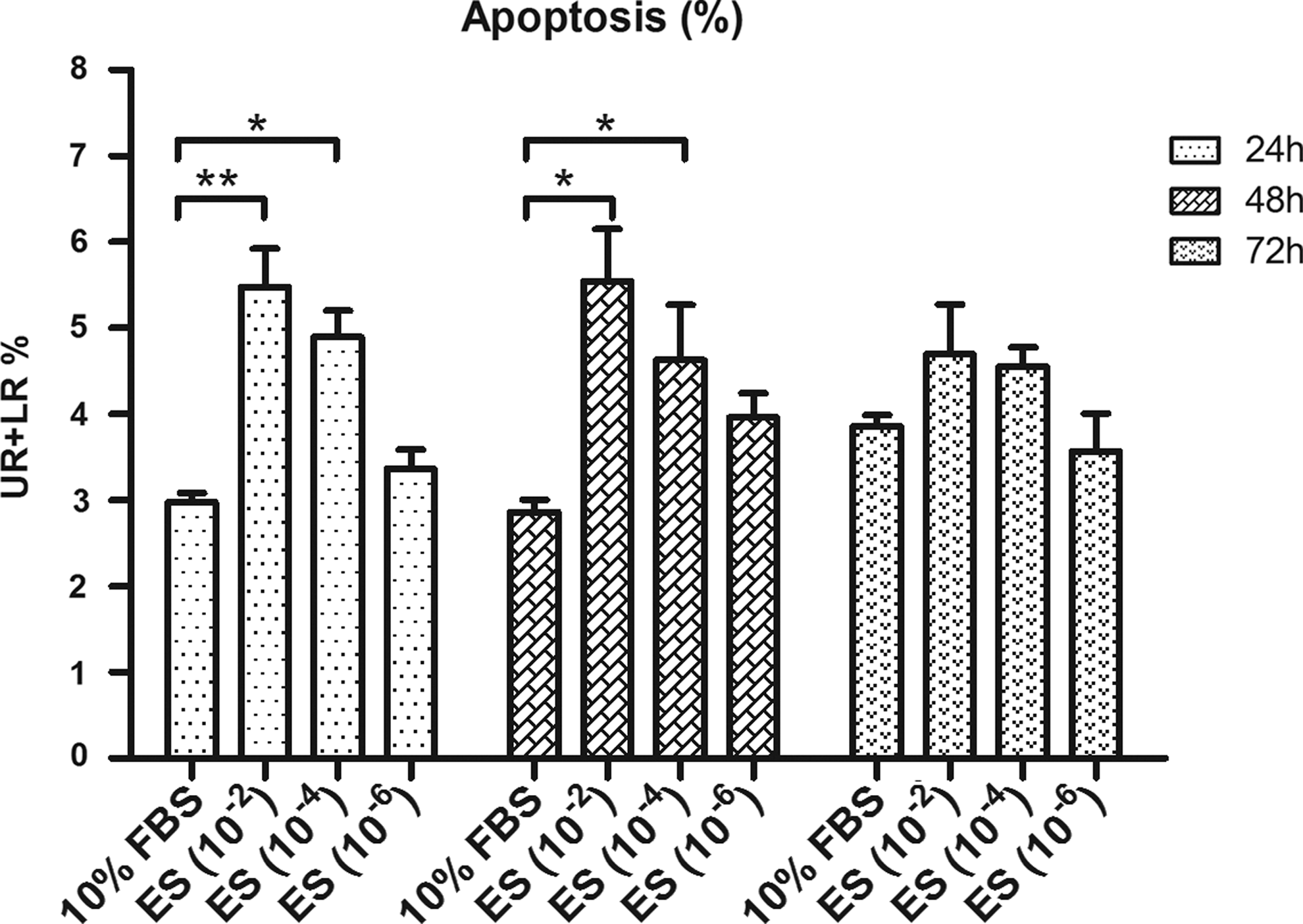
Effects of endostatin on HUVEC apoptosis. Apoptosis was quantified by flow cytometry measured by Annexin V and propidium iodide staining. The concentrations of endostatin investigated were 10−2 mg/mL, 10−4 mg/mL, and 10−6 mg/mL, and the testing time points were 24, 48, and 72 h. Data are presented as the mean±SEM. Each experiment was repeated in at least 3 independent trials. UR: upper right, present late apoptotic cells; LR: lower right, present early apoptotic cells; UR+LR: apoptotic cells. *P<0.05; **P<0.01.
UR, upper right quadrant; LR, lower right quadrant.
Endostatin inhibits HUVEC tube formation
Matrigel assays are one of the most widely used in vitro methods to evaluate the reorganization found in angiogenesis. In our study, endostatin-treated HUVECs showed an impaired capacity to form a regular network when treated with endostatin at concentrations of 10−2 and 10−4 mg/mL (Fig. 4B–D). Furthermore, the tubes formed in the presence of endostatin showed a significant difference in length when compared to the control and VEGF-treated groups (Fig. 4A).

Effects of endostatin on HUVEC tube formation. A Matrigel assay was used to evaluate the angiogenic potential of endostatin-treated HUVECs. The total length of the tube in each group is shown. The DMEM+10% FBS control was set to 100%. All treated groups were compared to the control group.
Effects of endostatin on the secretion of VEGF and PEDF
Endostatin-treated HUVECs showed a decrease in VEGF secretion in a time-dependent and dose-dependent manner. As shown in Fig. 5, after treatment with endostatin for 24 and 48 h, VEGF was downregulated when compared to the control group at concentrations greater than 10−5 mg/mL (P<0.05). However, after treatment for 72 h, the endostatin-treated group did not have significantly different levels of VEGF secretion when compared to the control group. In the PEDF secretion experiment, there was no significant change in PEDF after incubation of HUVECs with endostatin (data not shown here).
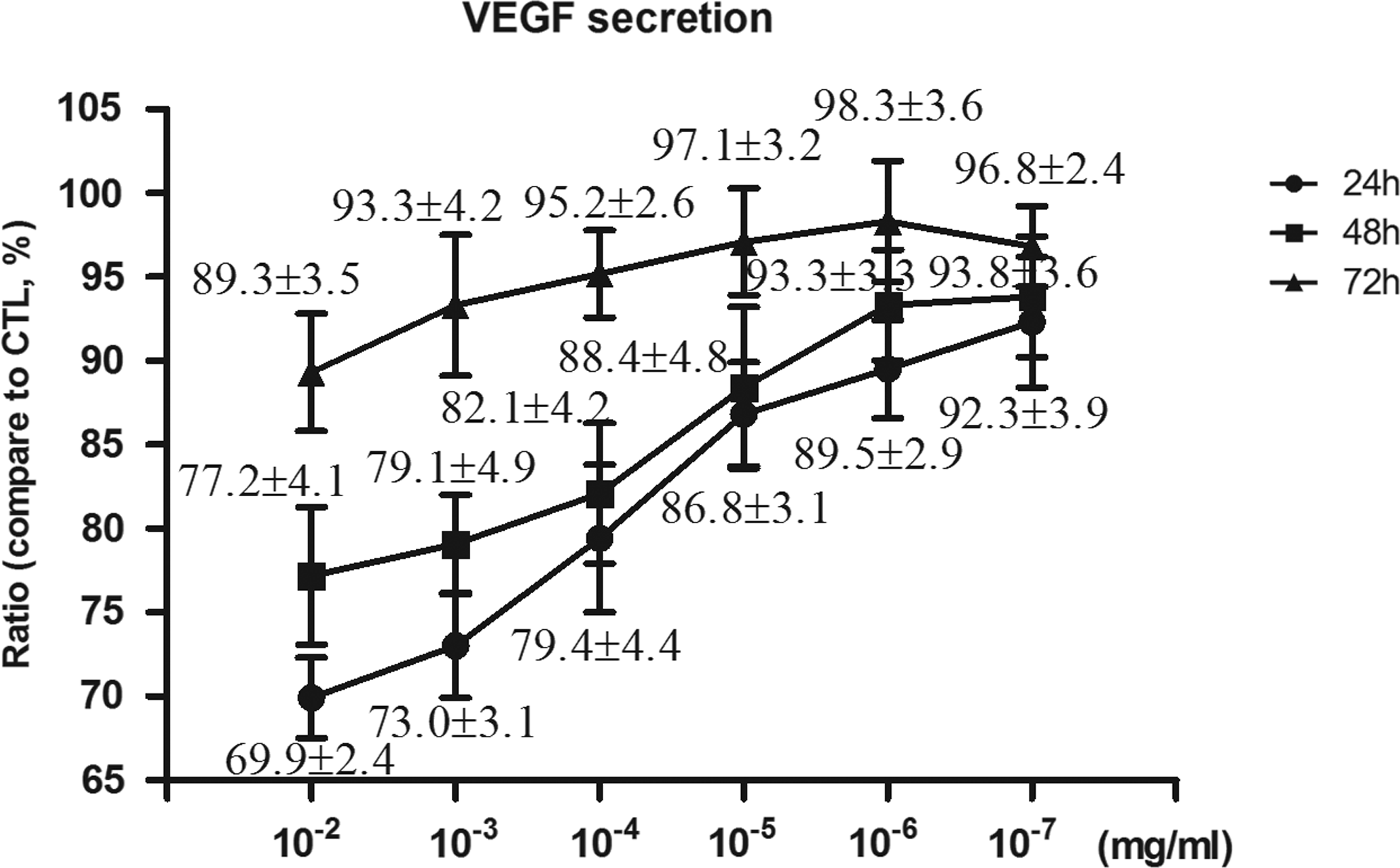
Effects of endostatin on VEGF secretion from HUVECs. VEGF released into the culture supernatant was measured using an Enzyme-Linked Immunosorbent Assay (ELISA) Kit. Endostatin-treated groups showed significant suppression in the level of VEGF secretion, both at 24 and 48 h. The DMEM+10% FBS control was set to 100%. Data are presented as the mean±SEM. Each experiment was repeated in at least 3 separate trials.
Endostatin protects against OIR retinal vessel loss
Endostatin has been shown to inhibit tumor growth. To determine whether endostatin had an antiangiogenesis effect in the murine OIR model, endostatin and IgG were injected intravitreally into the right eyes of retinopathy mice at p12 and to age-matched normal pups. As shown in Fig. 6, endostatin intravitreaous injection significantly reduced the neovascularized area to 23.0%±1.3%, which is significantly less than the area in the untreated control group (30.2%±0.8%, P<0.0001).
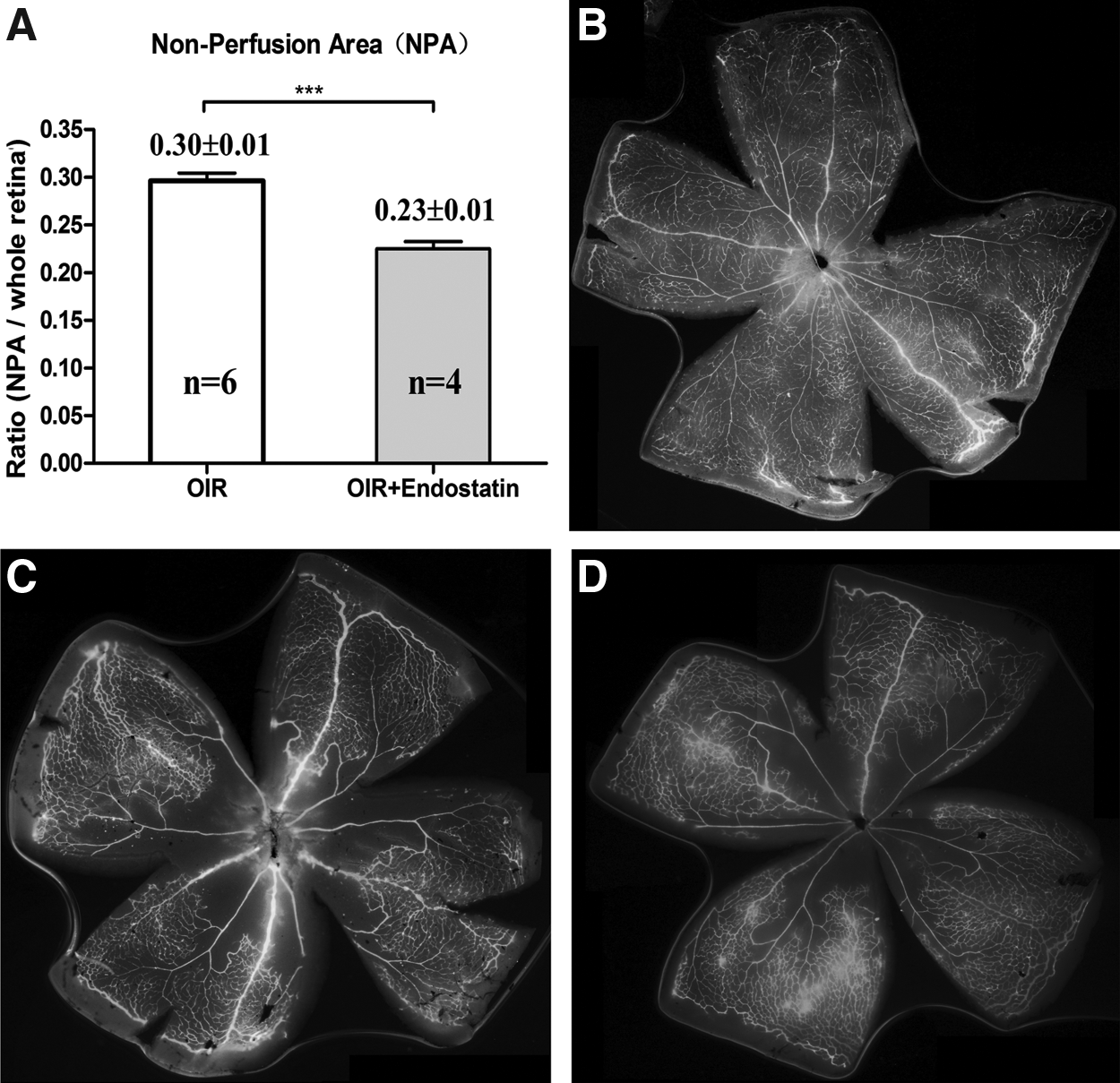
Effects of endostatin on oxygen-induced retinopathy (OIR) model. P7 mice pups were put into a hyperoxia condition for 5 days. At p12, the pups were intravitreously injected with PBS and endostatin (5 mg/mL) for 1.5 μL. At p18, the pups were euthanized to detect the retinal nonvascularization area.
Endostatin restores the PEDF/VEGF ratio
In our OIR model, treatment with endostatin reduced VEGF levels from 198.4±12.1 pg/mL retina protein in the control group to 81.6±7.7 pg/mL in the endostatin-treated group (Fig. 7A, P<0.0001). The PEDF level increased from 50.2±4.3 pg/mL in the OIR control group to 82.0±4.3 pg/mL in the endostatin-treated group (Fig. 7B, P<0.01). To better understand the balance between VEGF and PEDF, we measure the ratio of PEDF/VEGF. As shown in Fig. 7C, PEDF/VEGF is 0.26±0.03 in the OIR pups, and changes to 1.06±0.18 in the endostatin-treated group (P<0.0001), which is similar to the normal control group 0.93±0.09.

Retina concentration of VEGF and pigment epithelial-derived factor (PEDF) after treatment with endostatin. In the OIR model, the pups (5 pups in each group) were intravitreously injected with 5 mg/mL endostatin for 1.5 μL at p12 (immediately removed to normal room air). At p18, the pups were euthanized to collect their retinas to detect VEGF and PEDF by the ELISA Kit.
Discussion
Antiangiogenic treatment is one of the most effective therapeutic strategies for the management of retinal neovascularization diseases, including ROP, AMD, and DR, which can cause irreversible blindness.19,20 Recently, several agents have been demonstrated to be helpful in inhibiting retinal neovascularization, such as Avastin (bevacizumab) and Lucentis (ranibizumab), which mainly target VEGF.6,21 Although these agents have demonstrated significant antiangiogenesis effects, the efficiency is ∼60%–70%, and the efficacy is much lower when patients are in the later stages of disease, especially after scar formation.22–24
In recent years, the comprehensive study in angiogenesis has led scientists to develop “broad-spectrum” antiangiogenesis drugs with “multiple targets”, 25 such as KH902, which targets VEGF and PIGF at the same time.26,27 Endostatin was first discovered in 1997 8 and was reported as a powerful new physiological antiangiogenic agent with a broad-spectrum. 25 It has been widely used in tumor therapy based on its ability to cut off blood supply.28,29 According to subsequent in-depth studies, researchers found that endostatin not only targets VEGF, as Avastin targets VEGF, but that endostatin affects a wide variety of targets.11,30 Based on a gene array study, it was revealed that ∼12% of all genes in cells are affected after treatment with endostatin. In the context of cancer, endostatin specifically acts on genes that regulate angiogenesis (proangiogenesis and antiangiogenesis). 11 Endostatin's important signaling pathway may involve binding to α5β1 integrin on endothelium 31 ; blocking the activity of metalloproteinases 2, 9, and 13 32 ; upregulation of Thrombspodin-1, Ephrin A3, and Ephrin B3; and downregulation of Neuropilin-1 and HIF-1α. 19 In addition to the ability to inhibit angiogenesis, another prominent advantage of endostatin is its superior safety profile. According to the clinical reports, cancer patients experienced no toxicity effects and no resistance after administration for over 3 years without interruption.33,34 All of the evidences above show that endostatin is an exciting potential therapeutic for retinal neovascularization. Thus, thoroughly evaluating its effect on retinal neovascularization is meaningful. In previously published articles, endostatin has been demonstrated to inhibit retinal neovascularization by downregulation of VEGF and integrinβ3, 35 inhibit vasopermeability by stabilizing endothelial cell junctions.36,37 However, the other mechanisms need to be addressed also.
In the present study, we evaluated the antiangiogenesis effects of endostatin both in vitro and in vivo, and provided solid evidences for its potential use in retinal neovascularization. The in vitro study shows that endostatin can inhibit HUVEC proliferation (Fig. 1), even under conditions of VEGF stimulation (Fig. 1). The inhibitory effects were dose dependent and time dependent. The mechanism of proliferation inhibition may involve the suppression of VEGF expression (Fig. 5). Additionally, endostatin can inhibit HUVEC migration (Fig. 2) and induce HUVEC apoptosis (Fig. 3), which corresponds to previous reports. 38 We also evaluated the antiangiogenic properties of endostatin in the context of a tube formation study in vitro and with an OIR model in vivo. Both of these studies demonstrated that endostatin is a very potent inhibitor of angiogenesis. A possible mechanism for this inhibition is the restoration of the PEDF/VEGF ratio (Fig. 7).
As we know that VEGF plays a critical role in angiogenesis, overexpression of VEGF exacerbates neovascularization, while the withdrawal or blocking of VEGF causes suppression of vascular growth. 39 PEDF was discovered in 1991 by Tombran-Tink et al., and has been reported to be one of the most potent endogenous antiangiogenic agents. 40 Previous research has revealed that PEDF is downregulated in ROP. 41 Intravitreous injections of exogenous PEDF reduces areas of nonvascularization and induces apoptosis of vascular endothelial cells. 41 In our study, endostatin was able to restore the ratio of PEDF/VEGF in an OIR model; this may illustrate an important mechanism of endostatin inhibition of angiogenesis in vivo beyond the already known mechanisms. This result also corresponds with a previous study that reported the ratio between PEDF and VEGF is critical for angiogenesis. 42 As far as we know, this is the first study that demonstrated that endostatin plays a role in balancing the PEDF/VEGF ratio. Although we found the PEDF/VEGF ratio changed in our OIR model after treatment with endostatin, the application of endostatin to HUVECs did not induce the upregulation of PEDF in vitro. This may be because the HUVECs were cultured under normoxia conditions and not under simulated hypoxia as in other studies. 42
In summary, we comprehensively studied the antiangiogenesis effects of endostatin both in vitro and in vivo and found that endostatin could effectively inhibit retinal neovascularization in an OIR model and restore the balance between PEDF and VEGF, which may be a new and important mechanism in inhibition of angiogenesis. Our data suggest that endostatin may be an innovative approach, or an adjunctive therapeutic strategy, for future therapeutic strategies against retinal neovascularization.
Footnotes
Acknowlegment
This work was supported by the National Basic Research Program of China (973 Program, 2011CB510200) for LXX and Peking University People's Hospital Research and Development Fund for BYJ (RDB2012-24).
Author Disclosure Statement
No competing financial interests exist.