
Abstract
Abstract
Purpose
Methods
Results
Conclusions
Introduction
Normally, the cornea is protected from oxidative damage by the tear film, which contains several antioxidant mechanisms. 5 However, in patients with dry eye disease, where the tear film is deficient, the antioxidant mechanisms may not be adequately available to cells. Several studies have suggested a role for oxidative stress in the pathogenesis of dry eye disease. Wakamatsu et al. have shown that oxidative stress markers are elevated in the tears and conjunctival cells of patients with atopic keratoconjunctivitis. 6 Since the cornea is exposed to direct light, oxidative reactions are induced that can lead to ocular surface damage and DNA alterations. ROS generated by these reactions are generally quenched by primary scavengers, such as glutathione peroxidase, superoxide dismutase, and catalase in the tears, but Dogru et al. showed that patients with dry eye have significantly lower concentrations of scavengers than those with a healthy tear film. 7 A study by Cejkova et al. has also shown significantly reduced levels of naturally occurring antioxidant enzymes in patients suffering from dry eye disease. The decrease in enzyme expression appeared to be dependent on the severity of dry eye signs and symptoms. 8 Insufficiency of protection by antioxidants has also been reported by Augustin et al. to cause corneal inflammation, irritation, and oxidative tissue damage. 9 Since patients with dry eye may lack their natural antioxidant capabilities, supplemental antioxidants might be effective in reducing oxidative stress and damage to the cornea.
In addition to inadequate antioxidant protection, the inflammatory process has also been shown to initiate ROS production. Mast cells have been shown to produce ROS from both immunological and nonimmunological stimulation. 10 The production of ROS by mast cells occurs both intracellularly and extracellularly. Therefore, while the inflammation may be initiated due to insufficient antioxidant protection, ROS production can, in turn, potentially be perpetuated by inflammatory cells.
As more health risks due to oxidative stress are recognized, methods for prevention of oxidative damage have gained attention. As a result, antioxidants that quench the harmful effects of ROS are commonly included in topical dermatologic products and dietary supplements. Patients with dry eye frequently apply lubricant eye drops, also known as artificial tears, to alleviate the symptoms. Since the effects of oxidative stress in dry eye disease have become more apparent, we suggest that it may be desirable to incorporate an antioxidant into artificial tears to provide additional protection of the ocular surface from damage. Adding an antioxidant to a tear formulation would be particularly advantageous because the antioxidant would be applied directly to the target cells. Antioxidant supplements that are taken orally must be absorbed by the digestive tract, and there is little evidence that these compounds are delivered to and taken up by cells in adequate amounts to have a clinically relevant effect. While many antioxidants have been studied using chemical assays, the question still remains whether they are biologically available. For an antioxidant to work effectively, it must be taken up by cells and then quench ROS in the cytoplasm.
The purpose of this present study was to use the cellular antioxidant activity (CAA) assay that was developed by Wolfe and Liu 11 to measure the bioavailability of antioxidants found in fruits, to quantify the efficacy of antioxidants applied directly to corneal epithelial cells. The CAA procedure was first performed to determine if the assay was applicable to corneal cells in culture. Once the usefulness of the assay was established, experiments were conducted to determine if the time cells are exposed to an antioxidant has an effect on the activity. This is important because an eye drop is on the surface of the eye for only a brief period of time. CAA assays were performed using 2 sources of ROS, ABAP, which generates ROS intracellularly, 11 and a hypoxanthine/xanthine oxidase system, which generates ROS extracellularly. 12
Methods
Cell culture
An immortalized human corneal limbal epithelial (HCLE) cell line was obtained from Dr. Ilene Gipson (Harvard Medical School, Boston, MA). This cell line has an advantage over other corneal epithelial cell lines, in that, under appropriate conditions, it will stratify in a culture medium rather than at an air interface. The cells establish an epithelial barrier and multilayered morphology that closely models the in vivo corneal epithelium. Unlike previously developed corneal epithelial cell lines, they express mucins and galectin-3.13–15 As such, it is expected that the uptake of compounds by these cells will be similar to the uptake by the intact corneal epithelium in vivo.
For CAA assays, HCLE cells were seeded in 96-well plates at 6×104 cells per well and maintained in a 100 μL Keratinocyte-Serum Free medium (Invitrogen, Carlsbad, CA) for 24 h. After 24 h, the confluent cells were switched to a stratification medium (50% DMEM: 50% F12 medium with 10% calf serum and 10 ng/mL EGF), which causes formation of a multilayer culture. 13 Cells were allowed to stratify for 5 days before use. All cultures contained 100 U/mL penicillin and 100 μg/mL streptomycin.
Chemicals
The chemicals, all from Sigma (St. Louis, MO), used for the CAA assay include 2′,7′-dichlorofluorescin diacetate (DCFH-DA), 2,2′-azobis(2-amidinopropane) dihydrochloride (ABAP) xanthine oxidase, EDTA, FeCl3, and hypoxanthine. The antioxidants used in this study include quercetin (quercetin dihydrate), gallic acid, n-propyl gallate, and epigallocatechin gallate (EGCG). A 10-mM stock solution of quercetin was prepared in DMSO. Stock solutions of gallic acid and n-propyl gallate were prepared at 50 mM in ethanol. A 10-mM stock solution of EGCG was prepared in glycerol.
The antioxidants, ABAP, and the hypoxanthine/xanthine oxidase system were tested for effects on cellular viability using a Live/Dead viability/cytotoxicity kit, which uses calcein AM and ethidium bromide to detect viable and dead cells, respectively (Molecular Probes, Life Technologies, Grand Island, NY). No adverse effects on cell viability were detected during exposure times up to 4 h (data not shown).
CAA assay using ABAP as the oxidant
The medium was removed from stratified cultures, and the cells were washed with HBSS. Cells were loaded with 25 μM DCFH-DA in HBSS. Inside the cells, DCFH-DA is cleaved by esterases to form the more polar 2′,7′-dichlorofluorescin (DCFH), which cannot leave the cell. After loading for 1 h, the cells were washed with HBSS to remove extracellular DCFH-DA. Cells were exposed to HBSS with or without an antioxidant for 1 h. After exposure, the solution was removed and cells were washed with HBSS. Then, 100 μL of 600 μM ABAP in HBSS was added to each well. ABAP spontaneously generates ROS intracellularly. Inside the cell, ROS oxidize DCFH to form fluorescent DCF. The optimal concentrations of DCFH-DA and ABAP were determined in preliminary studies and are similar to those previously reported. 11
An initial fluorescence reading was made on a Cary Eclipse fluorimeter (excitation 485 nm, emission 538 nm) immediately following exposure to ABAP, after which, cells were placed in an incubator at 37°C. Fluorescence was measured at 20-min intervals for 60 min. All activity assays were repeated at least 3 times with similar results and the EC50 for each antioxidant was calculated for each experiment. The EC50s reported are the mean±SD for these 3 technical repeats.
Exposure time studies
To measure the effect of time of exposure to an antioxidant on the reduction of oxidative activity, cells were incubated with antioxidant solutions for varying lengths of time. Because ocular residence time of an eye drop has been shown to be about 15 min, the shortest exposure time was set at 10 min. 16 In these experiments, cells were loaded with 25 μM DCFH-DA as previously described. Following DCFH-DA incubation, cells were exposed to a single concentration of the antioxidant in HBSS for 10–60 min. The concentration of antioxidants used in each experiment was the EC50 calculated from the initial 60-min exposure experiments. After incubation with the antioxidant solutions, cells were exposed to 600 μM ABAP and fluorescence was monitored as described above.
To compare the change in EC50 of antioxidants at different exposure times, a CAA assay was run, in which, cells were exposed to varying concentrations of an antioxidant in HBSS for 10 or 60 min.
CAA assay using a xanthine oxidase system as the oxidant
To determine the efficacy of antioxidants in quenching ROS generated extracellularly, CAA assays were performed using a xanthine oxidase system. Xanthine oxidase, being a protein, does not cross the plasma membrane. In this system, ROS are generated by 10 mU/mL xanthine oxidase, 50 μM FeCl3, 75 μM EDTA (to keep iron ions in solution), and 500 μM hypoxanthine. The free radicals are generated by the xanthine oxidase-catalyzed conversion of hypoxanthine to xanthine, and then to uric acid. 12 The ROS that are generated outside of the cell by the xanthine oxidase system are able to diffuse into the cells and cause oxidative damage, or may cause damage to the cell surface. As in previous experiments, HCLE cells were loaded with 25 μM DCFH-DA, and then treated with antioxidants for 10 min. Following antioxidant exposure, the cells were exposed to the xanthine oxidase system. Fluorescence was then monitored for 60 min. Assays were performed at least 3 times with similar results and the means EC50 for each antioxidant was calculated from these technical repeats.
Xanthine oxidase coincubation studies
After determining that intracellular antioxidants could quench ROS that had been generated extracellularly, coincubation studies were performed to determine whether antioxidants could quench ROS both inside and outside of the cell. As in previous experiments, HCLE cells were loaded with 25 μM DCFH-DA followed by loading with an antioxidant for 10 min, and then washed with HBSS. After antioxidant treatment, the cells were exposed to the xanthine oxidase system with antioxidants also present in the medium. Fluorescence was monitored for 60 min. Assays were performed at least 3 times with similar results.
Quantification of CAA
The data from the fluorescence readings are reported as CAA units:
where∫SA is the integrated area under the sample fluorescence curve and∫CA is the integrated area under the control curve. From the CAA units, an effective dose (EC50) for the antioxidant is calculated by plotting the log of
Results
Dose–response CAA assay using ABAP as the oxidant
The dose–response studies show that all the antioxidants were able to quench ROS generated by ABAP (Fig. 1). Quercetin was the most potent with an EC50 of 2.98±0.48 μM (mean±SD for 3 experiments). EGCG and n-propyl gallate quenched ROS with EC50s of 3.41±0.27 μM and 6.3±1.5 μM, respectively. Gallic acid was an effective antioxidant, but required a greater concentration to effectively quench ROS, with an EC50 of 50.7±8.7 μM.

Efficacy of antioxidants against ROS generated by ABAP. HCLE cells were loaded with 25 μM DCFH-DA for 1 h followed by a 1-h treatment with
Effect of loading time on antioxidant activity
Cells were incubated with a single concentration of an antioxidant for 10–60 min. The concentration of each antioxidant was determined from the EC50 value calculated in the dose–response CAA experiments. The results of the exposure time studies show that all antioxidants effectively quenched ROS when applied to cells for less than 60 min. CAA values for exposure of cells to n-propyl gallate for less than 60 min were not significantly different from the CAA value for a 60-min exposure (Fig. 2). To further support the data from the exposure time study, a dose–response CAA assay was performed to compare the EC50 of the antioxidant incubated for 10 or 60 min. The EC50s calculated after 60- and 10-min incubations were not significantly different, showing that loading time was not a factor in antioxidant efficacy (Table 1).
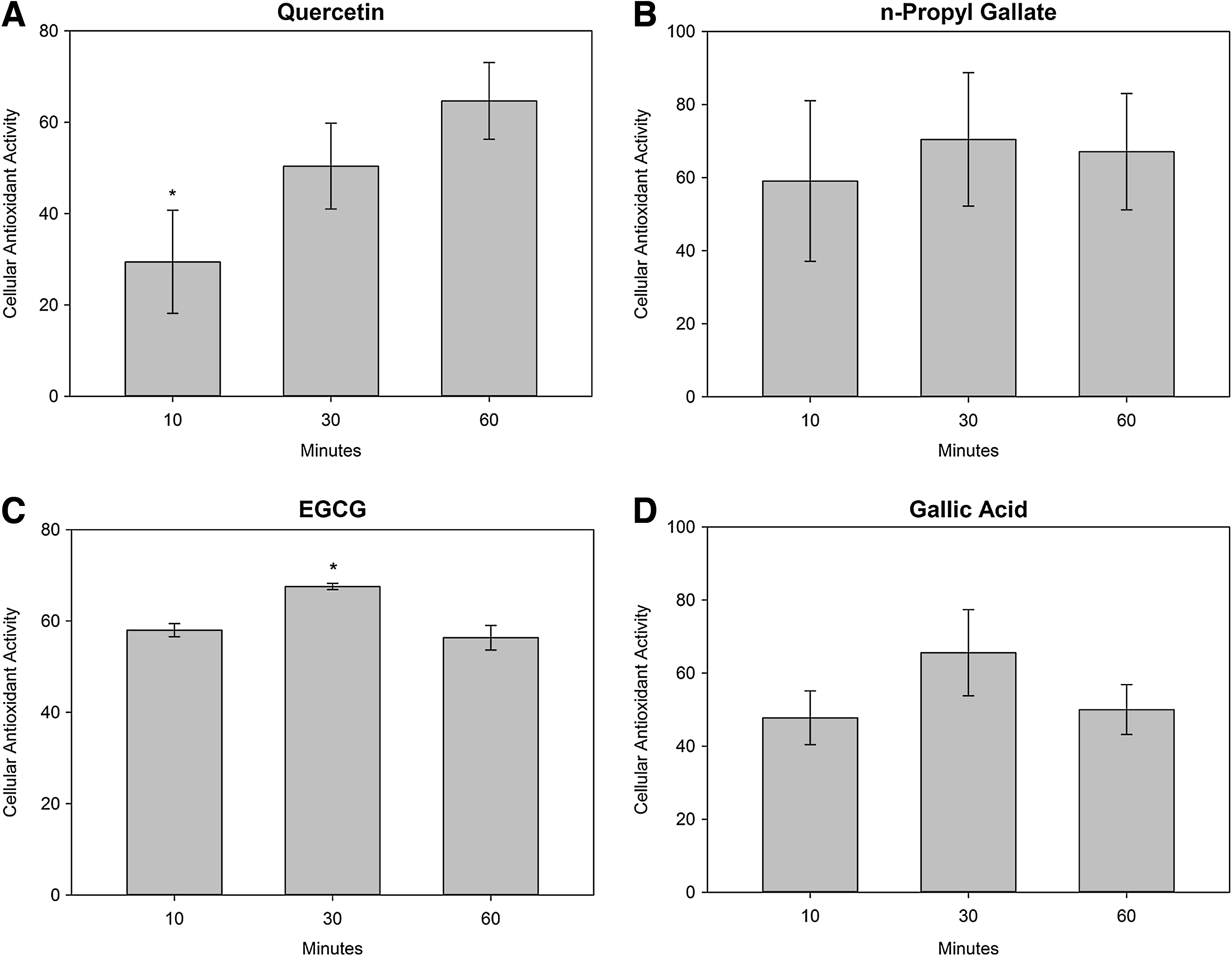
The effect of exposure time on the antioxidant activity of
Significantly different than 60-min exposure (mean±SD, t-test, P<0.05).
EGCG, epigallocatechin gallate.
Gallic acid was also able to quench ROS generated by ABAP at exposure times less than 60 min, with no significant difference in CAA values during exposure times of 10 or 60 min. Additionally, the EC50s after 10- and 60-min exposures were not significantly different.
When cells were exposed to EGCG for less than 60 min, ROS were effectively quenched and the 10-min exposure CAA value was not significantly different than the CAA value after loading for 60 min. The 30-min EGCG exposure did show a small, but significant difference from the 60-min exposure, however, this difference is probably not physiologically significant. In agreement with the exposure time data, the EC50s after a 10-or 60-min exposure to EGCG were identical.
In contrast to the other antioxidants, quercetin did not quench ROS as effectively after loading for 10 min, resulting in a CAA value significantly less than after a 60-min exposure. In agreement with the exposure time data, the EC50 of quercetin in cells loaded for 10 min was significantly higher than the EC50 after a 60-min incubation with antioxidants.
Dose–response CAA using xanthine oxidase as the oxidant
Results of CAA experiments using xanthine oxidase show that all antioxidants were able to quench ROS that were generated extracellularly (Fig. 3). Quercetin was the most potent with an EC50 of 41.3±5.6 μM (mean±SD for 3 experiments). n-Propyl gallate was able to quench ROS at an EC50 of 70.5±2.45 μM, while EGCG quenched ROS at an EC50 of 56.5±2.89 μM. Gallic acid was much less effective with an EC50 of 337.5±65.4 μM.
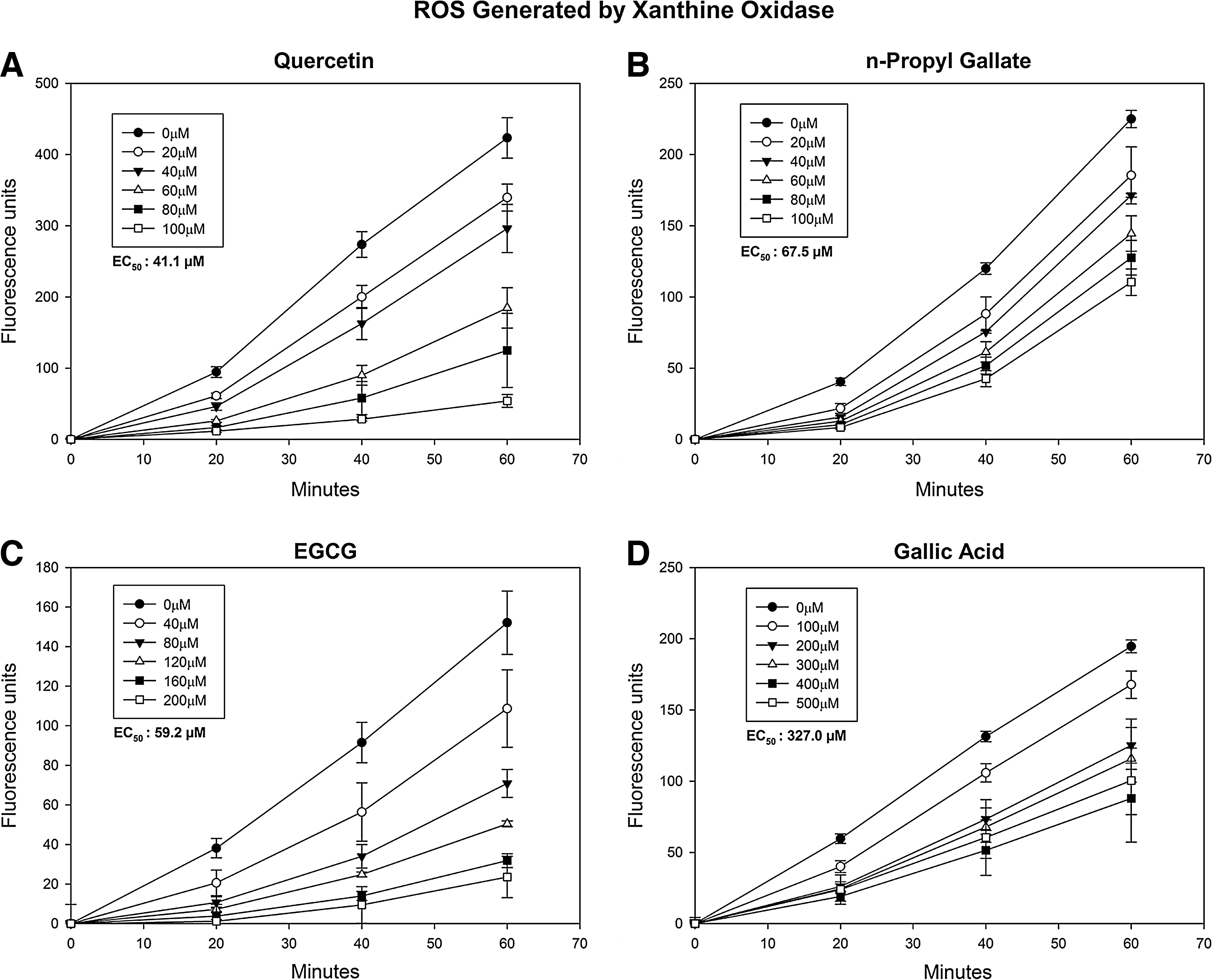
Efficacy of antioxidants against ROS generated by xanthine oxidase. HCLE cells were loaded with 25 μM DCFH-DA for 1 h followed by a 10-min treatment with
Xanthine oxidase coincubation studies
The coincubation experiments demonstrate that all antioxidants were able to quench ROS at significantly lower concentrations when the antioxidant was present in the culture medium as well as within the cells (Table 2). Under these conditions, n-propyl gallate was the most effective antioxidant. Quercetin and EGCG quenched ROS with similar EC50s. Gallic acid was much more effective when coincubated with xanthine oxidase, since EC50 decreased to 8 μM from a value of 338 μM when the antioxidant was only present intracellularly.
Significantly different than xanthine oxidase incubated separately (mean±SD, t-test, P<0.05).
Discussion
All of the antioxidants tested in this study were bioavailable to corneal epithelial cells and effectively quenched ROS generated intracellularly by ABAP and extracellularly by xanthine oxidase. Of the 4 antioxidants, quercetin was the most potent, followed, in order of effectiveness, by EGCG, n-propyl gallate, and gallic acid. Because the tear film turnover is rapid, the residence time of a topical ophthalmic formulation is relatively short. According to Meadows et al., the residence time of low viscosity fluids is ∼15 min. 16 Therefore, as is the case with all topical ophthalmic drugs, to be effective, it is necessary for an antioxidant to be taken up quickly by cells. Therefore, in addition to potency, the antioxidant activity was also evaluated based on the time cells were exposed to a particular compound. While quercetin proved to be the most potent antioxidant, results from the exposure time studies revealed that it was more slowly taken up by cells than the other compounds.
Buddi et al. 17 have provided the histochemical evidence for oxidative damage in several corneal diseases, including keratoconus, bullous keratopathy, and Fuchs' endothelial dystrophy. Oxidative damage has, however, received limited attention in relation to dry eye disease and was not emphasized by the International Dry Eye Workshop. 18 It is now becoming increasingly evident that ROS are involved in the dry eye syndrome. A major sign of dry eye is inflammation of the ocular surface and the mechanisms involved in this inflammation have recently been detailed by Barabino et al.18,19 Wakamatsu et al. hypothesized that oxidative stress could potentially play a causative role in the inflammatory process and pathogenesis of dry eye. 20 Their recent study demonstrates that the conjunctiva of patients with Sjögren's syndrome contains significantly more inflammatory cells than those with normal tear production. The increased inflammation was closely associated with increases in hexanoyl-lysine and 4-hydroxy-2-nonenal staining, 2 oxidative stress indicators resulting from lipid peroxidation. In a study by Cejkova et al., the antioxidant enzyme activity in the cornea was shown to be significantly decreased after exposure to UVB radiation that led to increased levels of ROS. 21 This is an important observation since the cornea is directly exposed to UVB radiation on a daily basis. Adverse effects of UVB and inflammation, which may result in oxidative damage in dry eye disease, suggest that an antioxidant applied directly to the cornea may be a rational means of preventing damage to the ocular surface epithelium.
Of the 4 antioxidants evaluated in this study, quercetin and EGCG have received the most attention as dietary supplements and nutriceuticals. Quercetin has been shown to effectively inhibit lipid peroxidation caused by UVB exposure. 22 It also inhibits production of inflammatory molecules, and directly inhibits apoptotic pathways. 23 We have also observed, however, that quercetin loses its antioxidant activity very quickly in aqueous solutions (unpublished data), which may limit its utility in an eye drop.
Recently, EGCG, a compound found in green tea, has been shown to be an effective antioxidant in attenuating symptoms of dry eye. In a study by Lee et al., dry eye mice were treated with topical EGCG. 24 Their results showed decreased corneal fluorescein staining, as well as decreased levels of inflammatory molecules such as interleukin-1β and chemokine ligand 2, all of which are clinical signs of dry eye disease. A study by Chan et al. also demonstrated the protective effects of EGCG. Chan showed that EGCG reduces H2O2 production induced by UVA exposure, as well as activation of MAP kinase, and cyclooxygenase-2 expression. 25 Cavet et al. tested the anti-inflammatory and antioxidative effects of EGCG on corneal epithelial cells in culture and reported similar results. 26 When treated with EGCG, the cells showed decreased concentrations of interleukin-1β and inhibition of MAPKs. Additionally, EGCG effectively quenched ROS generated by glucose oxidase in a dose-dependent manner.
In conclusion, all 4 antioxidants tested in this study were effective in quenching ROS in corneal epithelial cells, demonstrating their bioavailability when applied to the cells. While quercetin was an effective antioxidant, its low rate of cellular uptake and instability may limit its utility in a lubricant eye drop. EGCG and n-propyl gallate were taken up quickly and were effective at low concentrations. Given other reports of EGCG efficacy, this compound may prove to be an effective antioxidant in a lubricant eye drop.
Footnotes
Acknowledgments
Supported by a research fellowship provided by Calvin College (JLU) and the Den Ouden Undergraduate Research Fellowship (ARS). The authors thank Jenna Kennedy for technical assistance.
Author Disclosure Statement
This study was funded by a research grant from Alcon Research, Ltd., Fort Worth, TX. No competing financial interests exist.