
Abstract
Abstract
Purpose:
The trabecular meshwork (TM) outflow pathways of the aqueous humor show an increase in extracellular matrix in patients with primary open-angle glaucoma (POAG). The increase in TM extracellular matrix appears to be caused by transforming growth factor-β signaling and its downstream mediator connective-tissue growth factor (CTGF). Here we studied whether treatment with the prostaglandin F2α analog fluprostenol modulates the CTGF-mediated increase of the TM extracellular matrix.
Methods:
Human TM cells from 3 different donors were treated with CTGF (50 ng/mL) and/or fluprostenol (10−6 M and 10−7 M) and were analyzed by real-time reverse transcription polymerase chain reaction and Western blotting. Cell supernatants of the treated cells were analyzed by zymography.
Results:
Treatment with CTGF induced the expression and synthesis of CTGF, fibronectin, collagen type IV and VI, while treatment with fluprostenol alone had no effects. The effects of CTGF were blocked by 1-h pretreatment with fluprostenol in a dose-dependent manner. Treatment with fluprostenol or combined fluprostenol/CTGF induced the activity of matrix metalloproteinase 2 (MMP2) in TM cells, whereas treatment with CTGF alone had no effects on MMP2 activity.
Conclusions:
Fluprostenol blocks the fibrotic effects of CTGF on human TM cells and increases the activity of MMP2. Both effects have the distinct potential to attenuate a CTGF-mediated increase in TM extracellular matrix in patients with POAG and any effects on TM outflow resistance that may result from that.
Introduction
P
Prostaglandin (PG) F2α analogs are powerful ocular hypotensive agents that act on the cells in the aqueous humor outflow pathways. Early studies on the IOP-lowering effects of PGF2α analogs indicated that uveoscleral outflow is increased,21–23 an effect that is mediated via a widening of the spaces between the ciliary muscle bundles24,25 and an increase in the activity of metalloproteinases 26 that correlates with the degradation of extracellular matrix components between ciliary muscle bundles.27–29 More recent data obtained in human volunteers30,31 and perfused human anterior eye segments32,33 indicate that, at least in humans, PGF2α analogs may also decrease IOP upon acting on the TM outflow pathways. Still, the mechanisms by which PGF2α analogs may modulate trabecular outflow resistance are largely unclear.
Mechanistic studies on the action of agents that modulate TM outflow resistance in POAG are typically hampered by the lack of a suitable animal model for POAG. We recently developed a mouse model of POAG by increasing the expression of connective-tissue growth factor (CTGF) in the anterior part of the eye of mice. 34 CTGF belongs to the family of CCN (Cyr61; CTGF; NOV) matricellular proteins35,36 and is an important downstream mediator of transforming growth factor-β (TGF-β)–signaling in the TM. 37 In vitro, knocking down the expression of CTGF by siRNA attenuated the increase of extracellular matrix proteins in the TM upon treatment with TGF-β, strongly indicating that those effects are mediated by CTGF. 37 Moreover, CTGF overexpressing mice show a distinct increase in extracellular matrix components in the TM outflow pathways. 34
In the present study, we wanted to investigate whether PGF2α analogues have the potential to inhibit the CTGF-mediated increase in TM extracellular matrix. To this end, we treated cultured human TM cells (HTM) with fluprostenol, a potent PGF2α analog that is also used as fluprostenol isopropyl ester (travoprost) in the treatment of patients with POAG. 38
Methods
Cell culture
Cultures of HTM cells were established from the eyes of 3 human donors according to protocols published previously. 39 The age of the donors ranged from 34 to 76 years. HTM cells of the third to fifth passage were seeded in 35-mm culture wells (4.0×105 cells/well) and grown to a confluent monolayer in F10-HAM medium plus 10% (vol/vol) fetal bovine serum without antibiotics in 5% CO2 at 37°C (PAA, Pasching, Austria). The confluent cells were incubated in serum-free medium for 24 h followed by treatment with CTGF (50 ng/mL) or with fluprostenol (10−7M, 10−6 M) for 24 h. Combined treatment was performed by a 1-h preincubation with fluprostenol followed by adding CTGF for a total of 24 h. Fluprostenol was purchased from Cayman Chemicals (Ann Arbor, MI, Item Number 16768). Each of the described experiments was done with each of the 3 primary cell lines. Methods for securing human tissues were humane, included proper consent and approval, and complied with the Declaration of Helsinki.
RNA analysis
HTM cells were harvested and total RNA was extracted with TRIzol (Invitrogen, Karlsruhe, Germany) according to manufacturer's recommendations. First strand cDNA was prepared from total RNA using the iScript cDNA Synthesis Kit (BioRad, Munich, Germany) according to the manufacturer's instructions. Real-time reverse transcription polymerase chain reaction (RT-PCR) was performed on a BioRad iQ5 Real-time PCR Detection System (BioRad) using the following temperature profile: 40 cycles of 10 s melting at 95°C, 40 s of annealing and extension at 60°C. Primer pairs (Table 1) were purchased from Invitrogen and extended over exon–intron boundaries. RNA that was not reversely transcribed served as negative control for real-time RT-PCR. Glyceraldehyde-3-phosphate dehydrogenase and guanine nucleotide binding protein 2-like-1 were both used as housekeeping genes for relative quantification of the real-time RT-PCR experiments. Quantification was performed using iQ5 Standard-Edition (Version 2.0.148.60623) software (BioRad).
Western blot analysis
To obtain protein extracts, cells were directly lysed in RIPA lysis buffer (150 mM NaCl, 1% NP-40, 0.5% desoxycholic acid, 0.1% sodium dodecyl sulfate (SDS), and 50 mM Tris, pH 8) and protein content was measured with the bicinchoninic acid (BCA) protein assay (Pierce, Rockford, IL). Proteins were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis and transferred to polyvinylidine fluoride membranes. Antibodies were used as follows: goat anti-collagen type IV (1:500; Chemicon/Millipore, Billerica, MA), rabbit anti-collagen type VI (1:250, Rockland, Gilbertsville, PA), rabbit anti-fibronectin (1:1,000; Dako, Glostrup, Denmark), goat anti-human CTGF (1:500, Santa Cruz Biotechnology Inc., Santa Cruz, CA), chicken anti-goat, chicken anti-mouse and chicken anti-rabbit immunoglobulin Gs, coupled to horseradish peroxidase (1:2,000–1:10,000; Santa Cruz). Chemiluminescence was detected on a LAS 3000 imaging workstation (Raytest, Straubenhardt, Germany) after addition of sensitive chemiluminescent substrate, Luminata™ Forte Western HRP Substrate (Millipore, MA). For normalization, the signals of the control protein β-tubulin were determined and calculated using AIDA Image analyzer software (Raytest). The values of β-tubulin were used to normalize the signal intensity of the bands detected in Western blot analysis.
Gelatine zymography
Aliquots (10 mL) of cell culture medium collected from treated and untreated HTM cells were analyzed for gelatinolytic activity. Electrophoresis was performed at 4°C in 10% SDS polyacrylamide cross-linked gels, containing 0.1% gelatine (Sigma–Aldrich, Steinheim, Germany). Following electrophoresis, the gel was washed with 2.5% Triton X-100 and incubated in Tris/HCl, 0.5 mM CaCl2, 10−6 M ZnCl2, pH 8.0, at 37°C for 16 h. Subsequently, Coomassie brilliant blue staining was performed. The gel was washed in 5% acetic acid and 10% methanol in water. Presence of matrix metalloproteinases (MMPs) appeared as unstained bands. The gels were scanned on an LAS 3000 imaging workstation and the quantification was done by using AIDA Image analyzer software. Protein concentration of the cell culture medium was determined using the BCA method.
Number of experiments and statistical analysis
To assess the effects of the different treatments, each experiment was repeated at least 3 times with the primary HTM cell lines from each of the 3 different donors. An ANOVA test was used for statistical analysis, and results with a P-value<0.05 were regarded as significant and P-value<0.01 were regarded as highly significant.
Results
To analyze whether fluprostenol has an influence on the expression or synthesis of CTGF or typical TM extracellular matrix components such as fibronectin (FN), and collagen types IV (COL IV) and VI (COL VI), 14 HTM cells were treated with 10−6 M or 10−7 M fluprostenol for 24 h. Real-time RT-PCR did not detect a significant increase in the expression of CTGF mRNA in fluprostenol-treated cells when compared with controls (Fig. 1A). The same was true for the amounts of CTGF that were detected by Western blot analysis (Fig. 1B). We next investigated the expression of mRNA for FN, and the collagen subunits COL4A1 and COL6A1 following treatment with 10−6 M or 10−7 M fluprostenol for 24 h. Again, no significant differences were observed between fluprostenol-treated cells and controls (Fig. 1C, E, G). Finally, we analyzed the amounts of FN, COL IV, and COL VI in HTM cells by Western blot analysis to observe no differences between control and HTM cells treated with fluprostenol (Fig. 1D, F, H).
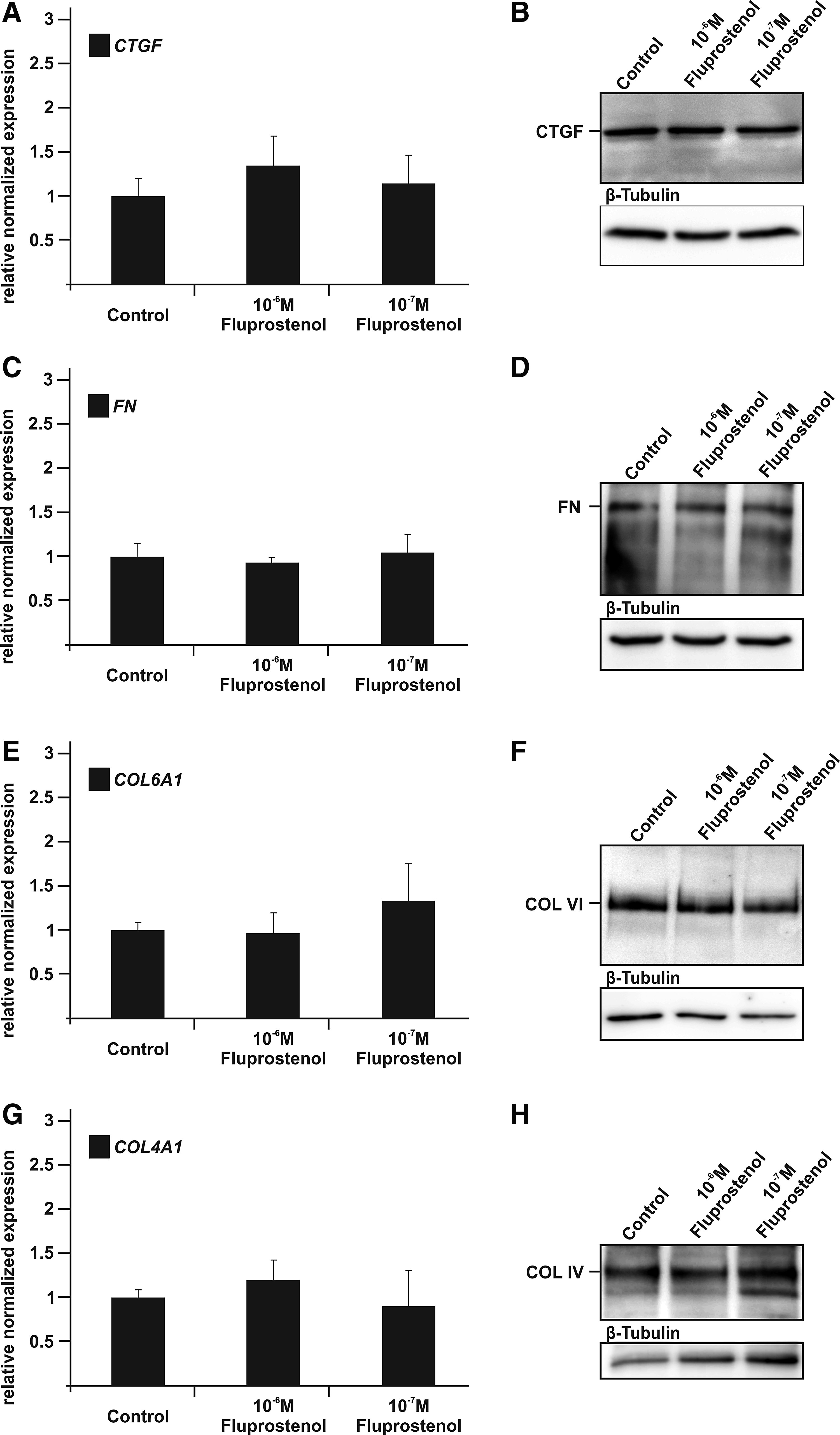
Effects of fluprostenol on human trabecular meshwork (HTM) expression and synthesis of connective-tissue growth factor (CTGF) and extracellular matrix components. (
Since CTGF has an autoinductive effect on its mRNA expression, 37 we were interested to see whether this effect is modulated by fluprostenol. In comparison to untreated controls (1.00±0.26), CTGF mRNA was significantly increased after 24-h CTGF treatment (1.52±0.12, P<0.05). Pretreatment with 10−6 or 10−7 M fluprostenol had no significant modulatory effect on the enhanced CTGF expression after CTGF treatment, which was still significantly higher than in controls (10−7 M: 1.48±0.4; 10−6 M: 1.34±0.15; both P<0.05; Fig. 2A). Comparable data were obtained by Western blot analysis, which showed significantly higher CTGF amounts than in controls (1.00±0.14) after combined fluprostenol/CTGF treatment (10−7 M: 1.61±0.32; 10−6 M: 1.54±0.25; both P<0.05) or treatment with CTGF alone (1.64±0.22; P<0.05) (Fig. 2B). Real-time RT-PCR analysis for FN confirmed previous studies 37 and showed a highly significant inductive effect of CTGF on its expression in HTM cells (2.01±0.16; P<0.01). One-hour pretreatment with 10−7 M fluprostenol did not cause a significant decrease of the FN mRNA induction (1.47±0.18; n.s.), whereas 10−6 M fluprostenol pretreatment led to a significant reduction of FN mRNA in comparison to treatment with CTGF alone (1.31±0.23; P<0.05; Fig. 2C). Quite comparable results were obtained by Western blot analysis (Fig. 2D). Treatment with CTGF caused a substantial significant increase in the amounts of FN (2.41±0.19; P<0.01), which was not changed after 1-h pretreatment with 10−7 fluprostenol (1.81±0.51; n.s.). In contrast, after pretreatment with 10−6 M fluprostenol, the amounts of FN were comparable to those in untreated controls and barely detectable. The impairment of the CTGF mediated FN synthesis by 10−6 M fluprostenol was significant (1.23±0.24; p<0.05) (Fig. 2D). Treatment with CTGF did also induce the expression of COL6A1 mRNA in the HTM cells (2.37±0.22; P<0.01), an effect that was not influenced by pretreatment with 10−7 M fluprostenol followed by CTGF treatment as the amounts of COL6A1 mRNA were still significantly higher than in controls (1.92±0.35; P>0.05). In contrast, pretreatment with 10−6 M fluprostenol significantly attenuated the effect of CTGF on the expression of COL6A1 mRNA (1.53±0.25; P<0.05; Fig. 2E). In correlation with the data obtained by real time RT-PCR, Western blot analysis showed that the amounts of COL VI in proteins from HTM cells were higher than in controls following CTGF (2.12±0.32; P<0.05) or 10−7 fluprostenol/CTGF treatment (1.92±0.42; n.s.), an effect that was significantly attenuated after 10−6 fluprostenol/CTGF treatment (1.45±0.12; P<0.05) (Fig. 2F). The significant induction of mRNA for Col4A1 mRNA, which was seen after CTGF treatment (2.10±0.33; P<0.01), was significantly attenuated by both concentrations of fluprostenol (10−7 M: 1.49±0.18; P<0.05; 10−6 M: 1.40±0.24; P<0.05; Fig. 2G). By Western blot analysis, the amounts of COL IV in proteins from HTM cells were detected to be higher than in controls following CTGF (2.53±0.41; P<0.05) or 10−7 fluprostenol/CTGF treatment (2.23±0.31; n.s.) (Fig. 2H). In marked contrast, pretreatment with 10−6 fluprostenol almost completely prevented the CTGF-mediated increase in COL IV (1.36±0.31; P<0.01) (Fig. 2H).

Fluprostenol modulates the effects of connective-tissue growth factor (CTGF) on extracellular matrix expression and synthesis in human trabecular meshwork (HTM) cells. (
Since treatment with prostaglandin analogs leads to an enhanced activity of MMPs in the cells lining the outflow region,40–42 we tested whether fluprostenol leads to an increase in the activity of MMP2 in HTM cells. Indeed, we observed a significant increase of MMP2 activity after treatment with 10−6 M and 10−7 M fluprostenol (10−7 M: 2.9±0.28; P<0.05; 10−6 M: 3.18±0.31; P<0.05; Fig. 3A). To analyze whether CTGF has a modulatory effect on the activity of MMP2 after fluprostenol treatment, we again performed combined treatments. CTGF treatment of HTM cells caused no changes in the activity of MMP2 in their cultured medium when compared to controls (0.89±0.22; P>0.05). Combination treatment with CTGF and 10−6 M fluprostenol showed no modulating influence of CTGF on the MMP2-stimulating effect of fluprostenol (2.89±0.41; P>0.05, Fig. 3B).
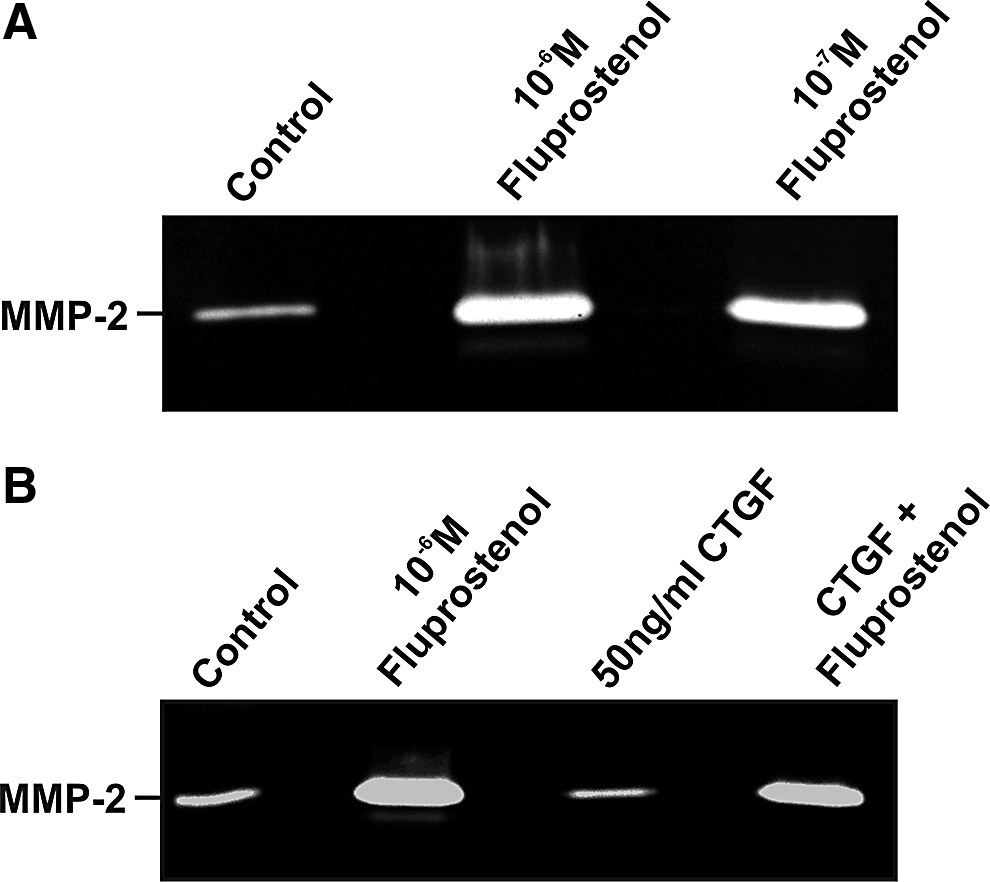
Influence of fluprostenol and connective-tissue growth factor (CTGF) on the activity of matrix metalloproteinase (MMP)-2. Zymography for MMP2 in culture medium from human trabecular meshwork cells after treatment with fluprostenol for 24 h (
Discussion
We conclude that the PGF2α analog fluprostenol reduces the fibrotic effects of CTGF on HTM cells. This conclusion is based on (1) the observation that pretreatment of HTM cells with fluprostenol reduces or prevents the inductive effects of CTGF on the expression of typical TM extracellular matrix molecules such as FN, COL IV, and COL VI, and (2) the finding that treatment with fluprostenol/CTGF increases the activity of MMP2 in HTM cells. Both effects may well attenuate a TGF-β/CTGF-mediated increase in TM extracellular matrix in patients with POAG.
The typical histopathological finding in the TM of patients with more advanced forms of POAG is an increase in fibrillar extracellular matrix in the juxtacanalicular region of the TM. 43 The increase involves an accumulation of banded fibrillar elements that are embedded in different glycoproteins and which have been termed plaque material.44,45 The molecular nature of plaque material is unclear, but there is evidence that COL VI is associated with it. 46 Other components of the extracellular matrix that have been found in higher concentrations in the TM of patients with POAG are COL IV, laminin, and FN.47,48 A likely explanation for the increase in TM extracellular matrix in POAG is the fact that aqueous humor of the majority of patients with POAG contains higher amounts of activated TGF-β2.49–54 TGF-β2 increases the expression of a multitude of extracellular matrix molecules including that of COL IV, COLVI, and FN in cultured HTM cells, and it is reasonable to assume that a similar scenario occurs in POAG eyes in situ. 55 In cultured HTM cells, the effects of TGF-β2 on the synthesis of extracellular matrix are mediated by CTGF, 37 which is highly expressed in the normal TM. 56 Ocular overexpression of CTGF in the mouse eye causes an increase in TM extracellular matrix along with an increase in IOP and subsequent optic nerve damage. 34 CTGF has also been detected in the aqueous humor of humans, 57 where it is increased in patients with pseudoexfoliation glaucoma. 58
Under normal conditions, the action of TGF-β/CTGF on TM extracellular matrix synthesis is embedded in a homeostatic signaling network 18 and antagonized by bone morphogenetic factors39,59 via Smad7, which inhibits the activity of TGF-β signaling. 60 PGF2α is synthesized by TM cells, 61 which have the appropriate receptors, 62 and it is tempting to speculate that locally secreted PGF2α serves an autocrine role in the TM microenvironment to contribute to the complex homeostatic system that balances any fibrotic effects of TGF-β/CTGF signaling. The balancing effects of PGF2α appear also to include other factors that lead to an increase in TM extracellular matrix, as we could show in a previous study that PGF2α analogs did also attenuate an increase in TM extracellular matrix following oxidative stress. 63 After failure of the homeostatic systems involved in the regulation of TM extracellular matrix synthesis followed by an excess deposition of TM extracellular matrix in POAG, medical therapy with PGF2α analogs may well have the potential to reduce this effect. It is of interest to note that our results point toward two different pathways by which PGF2α analogs modulate the amounts of extracellular matrix molecules in the TM, namely, by acting on their transcription and by increasing the activity of the degrading enzyme MMP-2, the dominant MMP in the TM.64,65
Our finding that 24-h treatment with 10−7 or 10−6 M fluprostenol did not affect the expression of CTGF mRNA in HTM cells is at variance with findings reported by Liang and coworkers, who found an increase in CTGF mRNA in TM cells from a 30-year-old donor after a 6-h treatment with 10−7 M PGF2α. 66 Currently we have no explanation for the different findings, but the different durations of treatment might be a contributing factor.
The effects of PGF2α analogs on the HTM extracellular matrix may well contribute to their IOP-lowering effects. In support of this assumption are observations that an increase in the activity of MMPs decrease TM outflow resistance in a perfused anterior eye segment organ model. 67 We realize that the concentrations of CTGF used in the present study were considerably higher than those measured in the human aqueous humor in previous studies.56,68 It is important to keep in mind, though, that CTGF is among the most highly expressed genes in the human TM in situ 56 and that the biologically relevant concentrations of CTGF in the TM microenvironment are very likely considerably higher than those measured in the aqueous humor. We are also aware of the fact that the concentrations of fluprostenol that have been measured in humans after daily administration69,70 are lower than those that were required to attenuate the biological effects of CTGF in our study. Still, it is difficult to compare the biological effects on cultured TM of an acute treatment with those on TM cells in situ that are treated chronically for weeks and months. Chronic treatment may well require lower doses to achieve similar effects to those seen with a high dose in an acute experiment. Finally, our experiments were performed in TM cells from obviously healthy donors, and TM cells from POAG patients might react differently. POAG TM cells might have changed their phenotype to that of a more ECM-synthesizing cell type, which is actually more reactive to the effects of Fluprostenol on CTGF than normal TM cells. Alternatively, the known loss of TM cells in POAG71,72 may blunt the effects of fluprostenol, especially in advanced cases of the disease. Clearly, animal experiments are needed to follow up on these issues, and the recently developed POAG mouse model with CTGF overexpression 34 appears to provide an ideal animal model to address those questions.
Quite intriguingly, CTGF is not only acting on extracellular matrix synthesis, but is also inducing the actin cytoskeleton in cultured mesenchymal cell such as kidney podocytes 73 or HTM cells 34 and, upon overexpression, in the mouse outflow pathways in situ. 34 The increase in components of its actin cytoskeleton very likely increases the contractility of the TM, a scenario that is known to increase trabecular outflow resistance.74–76 Thieme and colleagues reported that PGF2α analogs such as fluprostenol antagonize an endothelin-1-induced increase in TM contractility. Moreover, PGF2α and fluprostenol increase HTM monolayer impedance corresponding to a decreased cellular contractility. 77 It is tempting to speculate that PGF2α analogs may also be capable of antagonizing an increase in TM contractility induced by CTGF, an effect that may contribute to their IOP-lowering effects. Studies are under way in our laboratory to test this hypothesis.
Footnotes
Acknowledgment
The authors thank Corinna Unger for the excellent technical assistance.
Author Disclosure Statement
This study was supported by an unrestricted grant from Alcon Pharma GmbH, Freiburg im Breisgau, Germany.